- Эпигенетика. Эпигенетическая патология в процессах канцерогенеза. Часть 3

Содержание
- 2. Запросы клиники Ранняя диагностика Классификация, дифференциальная диагностика Прогноз развития заболевания, в том числе выживаемости, метастазирования, рецидивов
- 3. Что является молекулярно-генетическим маркером опухоли? Cтруктурные и функциональные изменения в ДНК -последовательности генов в опухоли, приводящие
- 5. Современная двухударная модель канцерогенеза
- 6. Изменение метилирования генома в процессе канцерогенеза
- 7. Пути инактивации классических генов-супрессоров опухолевого роста По экспериментальным оценкам около 200 генов мутируют в злокачественных опухолях
- 8. ПРОФИЛЬ МЕТИЛИРОВАНИЯ ГЕНОВ-СУПРЕССОРОВ В РАЗЛИЧНЫХ ТИПАХ ОПУХОЛЕЙ
- 9. Профиль метилирования генов, вовлеченных в канцерогенез, при раке молочной железы, немелкоклеточном раке легких, остром лейкозе и
- 10. ГЕТЕРОГЕННОСТЬ РПЖ Гиперплазия ПЖ. Стрелкой указаны области микродиссекции: гиперплазированный ацинус (ж) и смежная строма (стр).
- 11. Молекулярные маркеры рака предстательной железы Эпигенетические маркеры Экспрессионные маркеры Мутации AR, KLF6, p53, PTEN Делеции 8p22,
- 12. Частоты метилирования генов HIC1, р16, N33 и GSTP1 в микродиссекционных образцах предстательной железы
- 13. Сравнение частот аномального метилирования генов Р16, HIC1, N33 и GSTP1 в биопсийных и микродиссекционных образцах РПЖ
- 14. Сравнение молекулярно-генетических нарушений в опухолевом эпителии и смежной строме Метилирование Потеря гетерозиготности и нестабильность
- 15. Система маркеров аномального метилирования для ранней дифференциальной диагностики опухолей предстательной железы (частоты метилирования генов 50-90%): ADCY4,
- 16. Маркеры метилирования, определяемые на ранних стадиях канцерогенеза.
- 17. Частоты метилирования генов р16, MLH1, HIC1, N33, MGMT и RB1 в образцах ткани шейки матки.
- 18. МАРКЕРЫ МЕТИЛИРОВАНИЯ В ПРОГНОЗИРОВАНИИ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ ЖЕНСКОЙ РЕПРОДУКТИВНОЙ СИСТЕМЫ Частота аномального метилирования генов-супрессоров опухолевого роста (RASSF1,
- 19. Инактивация посредством гиперметилирования ключевых генов-супрессоров сопровождается гипометилированием и активацией целого ряда онкогенов (raf, c-fos, c-myc, c-Ha-ras,
- 20. Аномальное деметилирование гена IGF2 в образцах опухолей. Дорожка 18 – положительный контроль (ДНК здорового донора); 11
- 21. Свободная ДНК в периферической крови онкологического больного составляет около 200 нг/мл, в норме ее на порядок
- 22. 1 – отрицательный контроль; 2 и 7 – положительный контроль (негидролизованная ДНК); 3, 5, 6 и
- 23. Системы молекулярных маркеров метилирования
- 24. CpG-островки присутствуют в промоторных районах 60% генов, составляют от 0,5 до 1,5 т.п.н., содержание С +
- 25. Карта метилирования CpG-островка гена BIN1
- 26. Промоторы достаточно большого числа генов имеют CpG-островки протяженностью свыше 1000 п.н., которые характеризуются гетерогенным статусом метилирования.
- 27. Спектр геномной локализации продуктов непредвзятого скрининга Выявлено 26 новых локусов, аномально метилированных при РМЖ: 15% -
- 28. ФОРМИРОВАНИЕ СИСТЕМ МАРКЕРОВ МЕТИЛИРОВАНИЯ ДЛЯ ДИАГНОСТИКИ РМЖ Для диагностики рака молочной железы предложены две системы, оптимизированные
- 29. Метилирование ДНК является ценным биомаркером для диагностики рака. 1. Целый ряд генов, вовлеченных в канцерогенез, инактивируется
- 30. Метилирование промоторных районов генов RASSF и Р16 достоверно чаще происходит в клетках уротелиальных карцином с инвазией
- 31. Схема получения операционного материала РЖ
- 32. Изменение частот аномального метилирования ряда генов в ряду опухоль (рак легкого) – нормальная ткань 2см. –
- 33. Метилирование ДНК, как диагностический маркер онкологического заболевания, имеет ряд преимуществ перед другими маркерами Мы определяем положительный
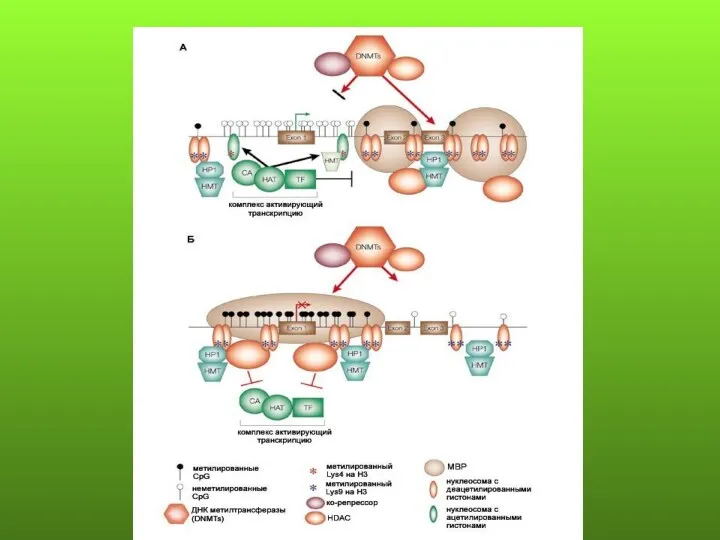
- 34. Деацетилирование гистонов, триметилирование лизина К27 и диметилирование лизина К9 гистона Н3 в районах активного хроматина, а
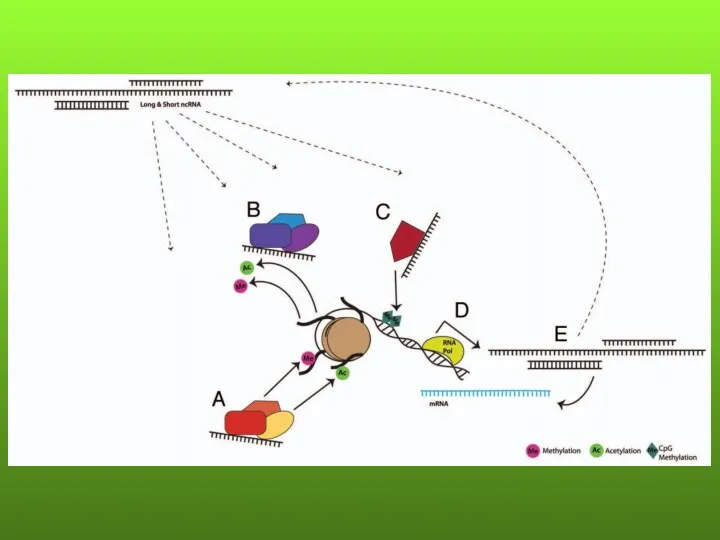
- 35. Патология генов, вовлеченных в эпигенетическую регуляцию, и канцерогенез
- 36. miРНК И КАНЦЕРОГЕНЕЗ miРНК часто локализуются в районах фрагильных участков, в минимальных районах перекрывания делеций при
- 37. Ломкие сайты и картированные miРНК 113 ломких сайтов и 186 miR. 61 miR локализованы в ломких
- 38. Локализация miРНК в районах делеций, амплификаций и транслокаций при раке
- 39. Хронический В-клеточный лимфолейкоз – минимальный район перекрывания делеций (~65% случаев) расположен в интроне гена LEU2, где
- 41. Клеточные онкогены K-RAS и MYC имеют множественные сайты комплементации с let-7, расположенные в 3’- НТР. Гиперэкспресия
- 43. miРНК могут действовать как онкогены и гены-супрессоры. Причем профиль экспрессии miРНК в опухолях более точно отражает
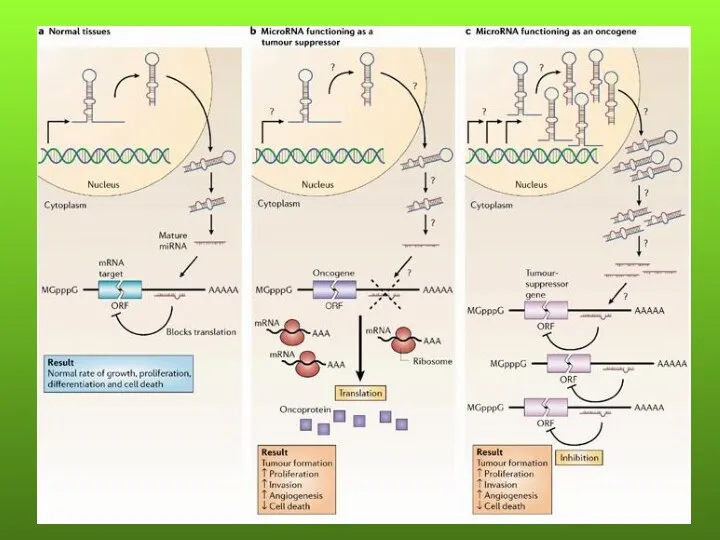
- 44. МОЛЕКУЛЯРНЫЕ МЕХАНИЗМЫ ДЕЙСТВИЯ нкРНК ПРИ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЯХ
- 45. Роль некодирующих РНК в регуляции эпителиально-мезенхимального перехода
- 46. Локализация miРНК, вовлеченных в канцерогенез различных органов и рак
- 48. микроРНК и рак oncomiR - онкогенные микроРНК, экспрессия в опухоли увеличена tumor-suppressor tsmiR – супрессорные микроРНК,
- 50. Двойственная роль многих микроРНК – онкогенная и супрессорная
- 52. Внеклеточные микроРНК В крови микроРНК циркулируют в составе экзосом или в комплексе с: Ago2, Argonaute2 NPM1,
- 53. Циркулирующие нкРНК как биомаркеры различных видов рака
- 54. Одна микроРНК может регулировать сотни генов-мишеней. Аберрантная экспрессия микроРНК инициирует многие заболевания, в том числе рак.
- 55. Возможности терапии с помощью нкРНК
- 56. Стратегии эпигенетической терапии
- 57. ХИМЕРНЫЕ ГЕНЫ
- 58. Типы химерных генов
- 59. Схема транслокации хромосом 8 и 14, в результате которой возникает гиперэкспрессия протоонкогена MYC
- 60. Структурные перестройки при лимфоме Беркита Протоонкоген с-Myc в результате перестроек оказывается под влиянием сильных промоторов генов
- 61. Транслокация хромосом 9 и 22 с образованием филадельфийской хромосомы и химерного онкогена BCR/ABL при хроническом миелоидном
- 62. Химерный ген TMPRSS2/ERG Особенностью канцерогенеза РПЖ является гиперэкспрессия факторов транскрипции семейства ETS вследствие хромосомных перестроек del(21)(q22.2;q22.3),
- 64. Схема образования химерных онкогенов RET/PTC в результате инверсий и транслокаций хромосомы 10. Схематическая структура протоонкогена RET,
- 65. Модель обусловленного транскрипцией химеризма (межгенного сплайсинга) Транс-сплайсинг подразумевает две различные молекулы РНК генов, находящихся на разных
- 66. Межгенный сплайсинг, обусловленный транскрипцией
- 67. От 2 до 5% всех генов может быть вовлечено в процесс межгенного сплайсинга. Описано явление транс-сплайсинга
- 68. Химерные гены при раке предстательной железы: В результате структурных перестроек: TMPRSS2-ERG, TMPRSS2-ETV1, TMPRSS2-ETV4, TMPRSS2-ETV5, HNRNPA2B1-ETV1, HERV-K-ETV1,
- 69. Химерные гены при ювенильной карциноме почки В результате структурных перестроек: MALAT1-TFEB, ASPSCR1-TFE3, PRCC-TFE3, CLTC-TFE3, NONO-TFE3, SFPQ-TFE3
- 70. Подвижность генов – важный фактор их регуляции. Кластеры активных генов - 150-200 т.п.н., расстояние между кластерами
- 71. АЛЬТЕРНАТИВНЫЙ СПЛАЙСИНГ И ПАТОЛОГИЯ У ЧЕЛОВЕКА
- 72. 13985 23341
- 73. Около 70% генов человека могут подвергаться альтернативному сплайсингу, что вносит значительный вклад в разнообразие протеома и
- 74. А. – консенсусные последовательности донорного сайта сплайсинга, акцепторного сайта сплайсинга и branch-сайта в интронах эукариот. В.
- 75. Четыре коротких последовательности нуклеотидов позволяют определить интрон: экзон-интронные границы в 5’ и 3’ концах интрона, сайт
- 76. Y –пиримидины (С/Т); ESE – экзонные энхансеры сплайсинга; ESS – экзонные сайленсеры сплайсинга; ISE - интронные
- 77. Экзонные энхансеры сплайсинга представляют собой сайты связывания обогащенных серином и аргинином белков (SR-белки). Эти белки являются
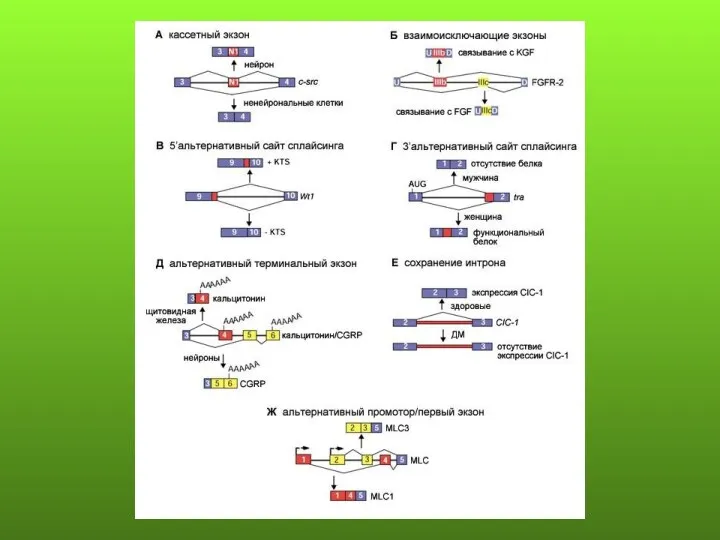
- 79. Основные формы альтернативного сплайсинга: 1) пропускание экзона составляет около 38%; 2) альтернативные 3’ и 5’ сайты
- 82. CD44 - мультифункциональный рецептор, вовлеченный в межклеточные взаимодействия, перемещение клеток, метаболизм гиалуроновой кислоты, передачу сигналов для
- 83. миРНК могут регулировать альтернативный сплайсинг. Их возможное участие согласуется с данными о высокой консервативности последовательностей, примыкающих
- 84. 60-70% генома транскрибируется с одной или обеих цепей ДНК. 5880 транскрипционных кластеров генома человека формируют смысловые/антисмысловые
- 86. ПАТОЛОГИЯ ТИПА «ЭФФЕКТ ПОЛОЖЕНИЯ» У ЧЕЛОВЕКА
- 87. Эффект положения - нарушение или изменение уровня экспрессии гена в результате изменения его местоположения в структуре
- 88. ПРЕДПОЛАГАЕМЫЕ МЕХАНИЗМЫ ЭФФЕКТА ПОЛОЖЕНИЯ Перемещение гена из района с определенной структурой хроматина в район с другой
- 94. Доказанный эффект положения Ген PAX6 - транскрипционный фактор. Хромосома 11p13. Аниридия - врожденная патология глаз, характеризующаяся
- 95. Ген PITX2 - транскрипционный фактор. Хромосома 4(q26). Синдром Ригера - олигодонтия, аномалии глаз, аномалии развития. В
- 96. ОБЩИЕ ЧЕРТЫ РАЙОНОВ, ПОДВЕРЖЕННЫХ ИМПРИНТИНГУ И ЭФФЕКТУ ПОЛОЖЕНИЯ 1. Протяженность эффекта (4 м.п.о. и более); 2.
- 99. «белок-кодирующий район с определенными регуляторными последовательностями» «транскрипционная единица» или «фрагмент ДНК, ответственный за синтез функционально значимого
- 100. 1860 – 1900х: «Ген – это дискретная единица наследственности» (Wilhelm Johannsen); 1910х: «Ген – это отдельный
- 101. Роль некодирующих РНК в регуляции эпителиально-мезенхимального перехода
- 103. Система молекулярно-генетических маркеров для карцином желудка Эпигенетические Генетические (структурные) Делеции и МН 16q22.1 (CDH1) 17p13 (ТР53)
- 104. Различия в поведении молекулярных маркеров в клинических группах можно использовать в качестве прогностических показателей течения рака
- 105. Методы поиска дифференциально метилированных CpG-островков Рестрикционно-ориентированное геномное сканирование Репрезентативно-дифференциальный анализ Микрочипы Метил-чувствительный фингерпринтинг - наименьшая себестоимость
- 108. Протоколы диагностики систем молекулярных маркеров в нейроонкологии Материал опухоли (парафиновый блок) Периферическая (венозная) кровь Морфологическое исследование
- 109. Принцип метода метилчувствительного фингерпринтинга MspI HpaII
- 110. Гены, идентифицированные методом МЧФП
- 111. Принцип АИМС Геномную ДНК обрабатывают метилчувствительной рестриктазой SmаI, которая формирует фрагменты с «тупыми» концами. Метилированные сайты
- 112. Высокоразрешающие карты метилирования CpG-островков генов BIN1, SEMA6B & LAMC3
- 113. Карта метилирования CpG-островка гена SEMA6B контрольные образцы (n=9) образцы РМЖ (n=8)
- 117. Теория «полей канцеризации»
- 118. Лечение с использованием ингибиторов гистондеацетилаз индуцирует ацетилирование гистонов и негистоновых белков, которые модифицируют уровень экспрессии генов,
- 119. Вопросы и методы науки Геномика Эпигеномика Транскриптомика Протеомика Сколько и каких изменений достаточно для классификации и
- 120. Как найти «ген рака»?
- 121. Сколько и какие гены несут мутации в опухоли? До внедрения полногеномного секвенирования было известно около 400
- 122. Соматические мутации в опухоли: драйверные и пассажирские Драйверные мутации возникают в ключевых генах контроля пролиферации, дифференцировки,
- 123. Соматические мутации в опухоли: драйверные и пассажирские Прочие соматические мутации, составляющие в геномах опухолей большинство, получили
- 124. Каталоги соматических мутаций Каталог 1 - ОМЛ: Ley 2008
- 125. Каталог 2 - ОМЛ: Mardis 2009 Уровень ложноположительных результатов снижен с 96% до 47% 12 мутаций
- 126. микроРНК – прогноз и лечение Выживаемость – корреляция с экспрессией miR-21, miR-155, miR-210, miR-197 Экспрессия miR-21
- 127. Методы анализа метилирования 1. Метилчувствительная ПЦР (Not1, Eag1, SacII, HpaII, HhaI) аналитическая чувствительность - 1: 2000
- 128. Принцип метода метилчувствительного фингерпринтинга MspI HpaII
- 129. Молекулярная патология гена BIN1 при РМЖ: метилирование Метилспецифическое секвенирование Результаты метилспецифического секвенирования фрагмента промоторного CpG-островка гена
- 130. УЧАСТИЕ В КАНЦЕРОГЕНЕЗЕ НЕКОТОРЫХ ИЗ ВЫЯВЛЕННЫХ ГЕНОВ
- 131. Схема анализа метилирования с использованием биологических микрочипов
- 132. Частоты метилирования генов MGMT, RASSF1A, GSTP1, RARβ, CDKN2A, ING в различных опухолях
- 133. Определение метилирования генов, вовлеченных в канцерогенез, методом метилчувствительной ПЦР
- 134. Схема чипа для определения метилирования генов, вовлеченных в канцерогенез
- 135. Примеры анализа образцов клинического материала с применением микрочипа низкой плотности для анализа метилирования. Вверху слева: гибридизация
- 137. Среднее значение экспрессии let-7a, miR-155, miR-205 в образцах рака легкого
- 138. Примеры генов-супрессоров опухолевого роста, CpG-островки которых могут быть метилированы при различных злокачественных новообразованиях.
- 139. Примеры генов-супрессоров опухолевого роста, CpG-островки которых могут быть метилированы при различных злокачественных новообразованиях.
- 140. Метилирование позволяет предсказать поведение опухоли (эффективность терапии, метастазирование): - метилирование WIT1 коррелирует с хеморезистентностью при ОМЛ;
- 141. микроРНК – регуляторы различных процессов
- 142. Лекарственные средства для аномальных эпигенетических событий Ингибиторы ДНК-метилтрансфераз 5-Azacytidine Vidaza (лейкемия, МДС) 5-Aza-2’-deoxycytidine Decitabine (лейкемия, МДС)
- 143. Выявленные гены и локусы
- 145. Скачать презентацию

Запросы клиники
Ранняя диагностика
Классификация, дифференциальная диагностика
Прогноз развития заболевания, в том числе выживаемости,
Запросы клиники
Ранняя диагностика
Классификация, дифференциальная диагностика
Прогноз развития заболевания, в том числе выживаемости,

Что является молекулярно-генетическим маркером опухоли?
Cтруктурные и функциональные изменения в
Что является молекулярно-генетическим маркером опухоли?
Cтруктурные и функциональные изменения в

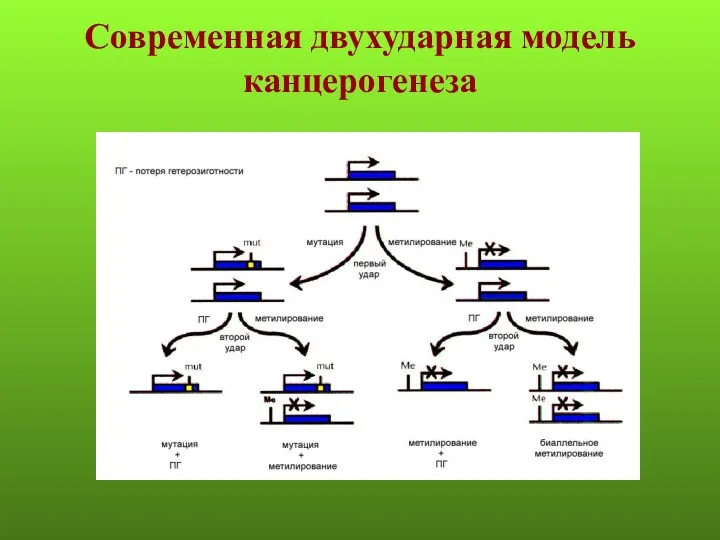
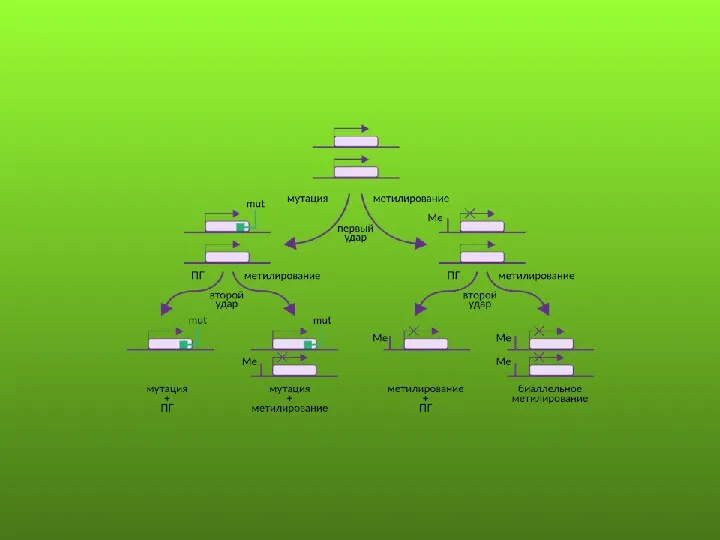
Современная двухударная модель канцерогенеза
Современная двухударная модель канцерогенеза
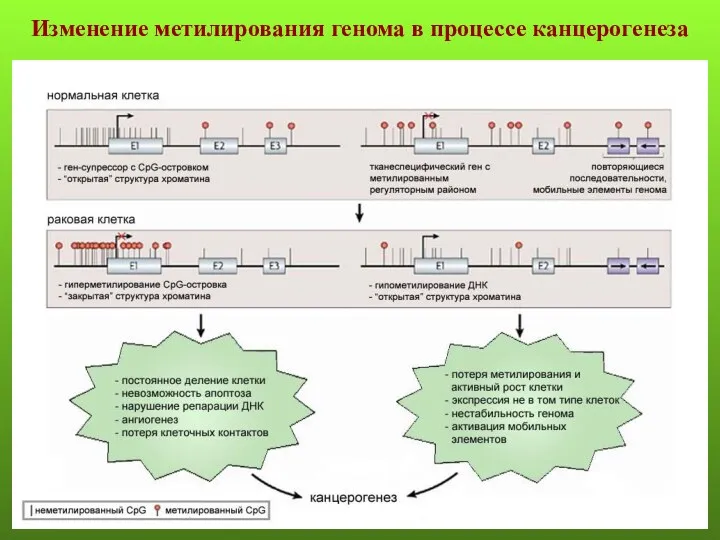
Изменение метилирования генома в процессе канцерогенеза
Изменение метилирования генома в процессе канцерогенеза
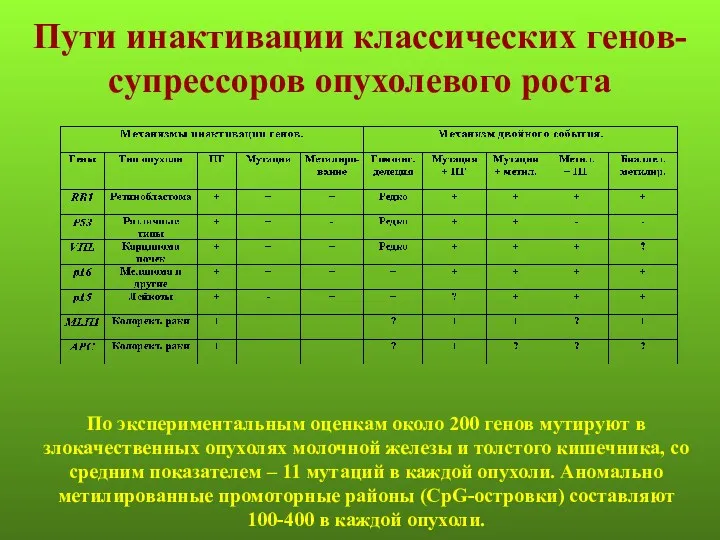
Пути инактивации классических генов-супрессоров опухолевого роста
По экспериментальным оценкам около 200 генов
Пути инактивации классических генов-супрессоров опухолевого роста
По экспериментальным оценкам около 200 генов
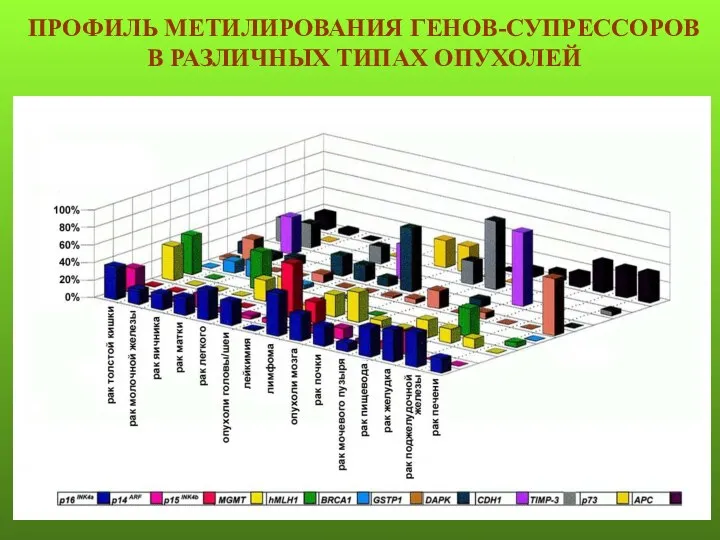
ПРОФИЛЬ МЕТИЛИРОВАНИЯ ГЕНОВ-СУПРЕССОРОВ
В РАЗЛИЧНЫХ ТИПАХ ОПУХОЛЕЙ
ПРОФИЛЬ МЕТИЛИРОВАНИЯ ГЕНОВ-СУПРЕССОРОВ
В РАЗЛИЧНЫХ ТИПАХ ОПУХОЛЕЙ
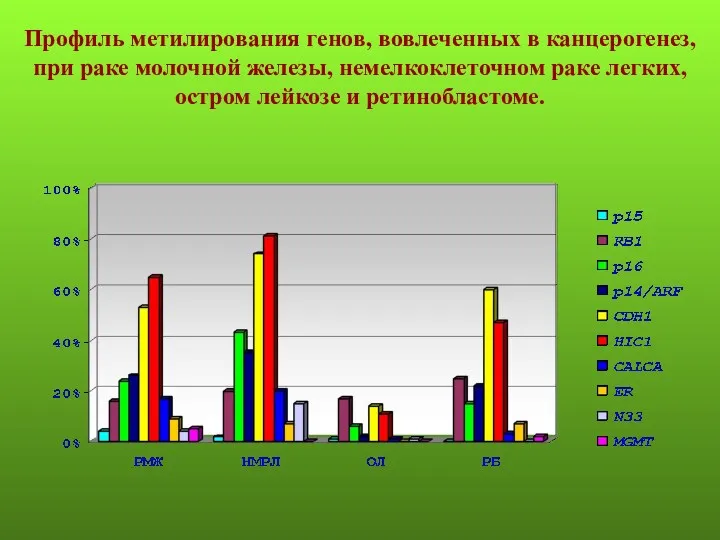
Профиль метилирования генов, вовлеченных в канцерогенез, при раке молочной железы, немелкоклеточном
Профиль метилирования генов, вовлеченных в канцерогенез, при раке молочной железы, немелкоклеточном
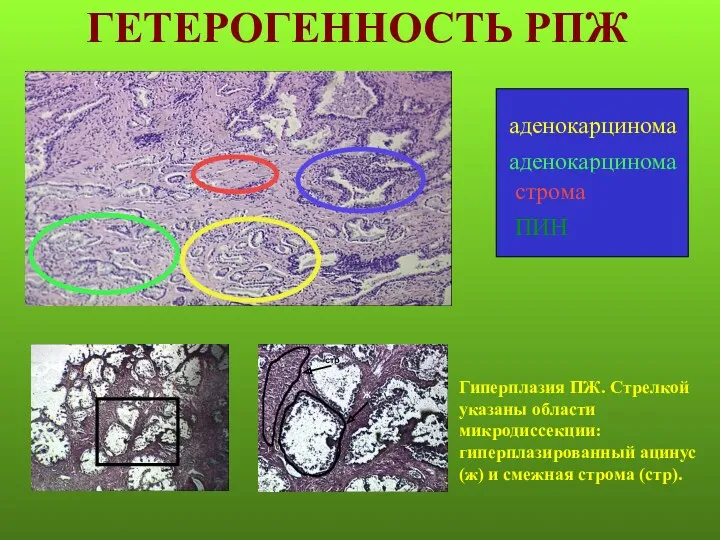
ГЕТЕРОГЕННОСТЬ РПЖ
Гиперплазия ПЖ. Стрелкой указаны области микродиссекции: гиперплазированный ацинус (ж) и
ГЕТЕРОГЕННОСТЬ РПЖ
Гиперплазия ПЖ. Стрелкой указаны области микродиссекции: гиперплазированный ацинус (ж) и

Молекулярные маркеры рака предстательной железы
Эпигенетические маркеры
Экспрессионные маркеры
Мутации
AR, KLF6, p53,
PTEN
Делеции
8p22, 16q24
RB1,
Молекулярные маркеры рака предстательной железы
Эпигенетические маркеры
Экспрессионные маркеры
Мутации
AR, KLF6, p53,
PTEN
Делеции
8p22, 16q24
RB1,
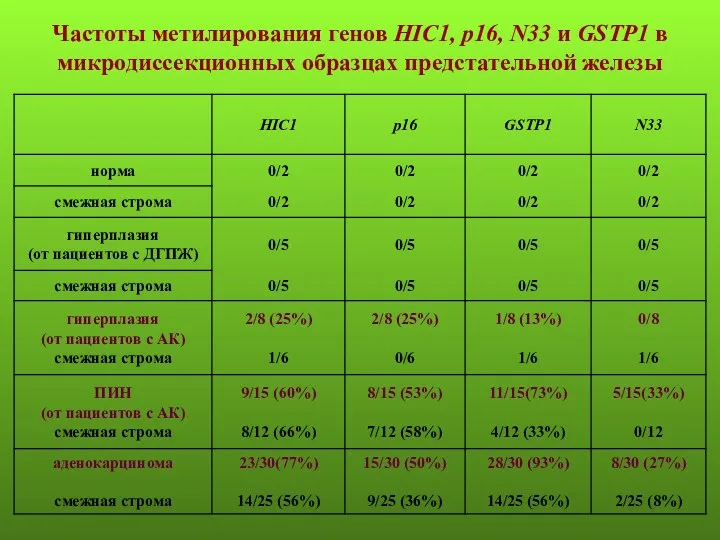
Частоты метилирования генов HIC1, р16, N33 и GSTP1 в микродиссекционных образцах
Частоты метилирования генов HIC1, р16, N33 и GSTP1 в микродиссекционных образцах
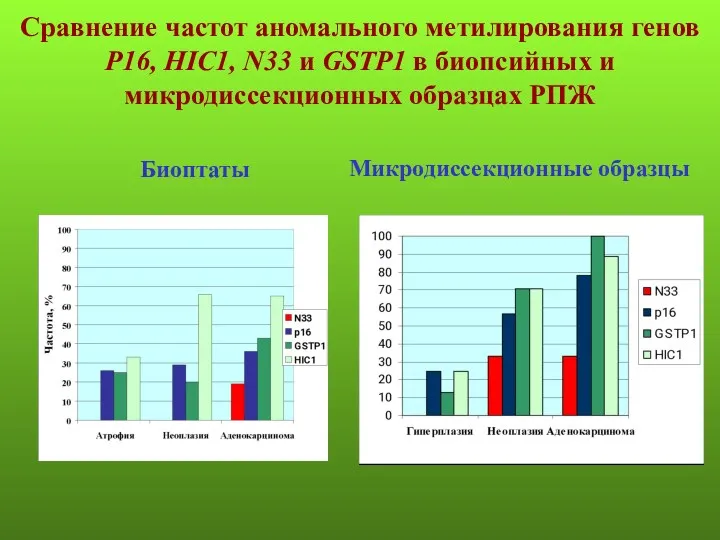
Сравнение частот аномального метилирования генов Р16, HIC1, N33 и GSTP1 в
Сравнение частот аномального метилирования генов Р16, HIC1, N33 и GSTP1 в
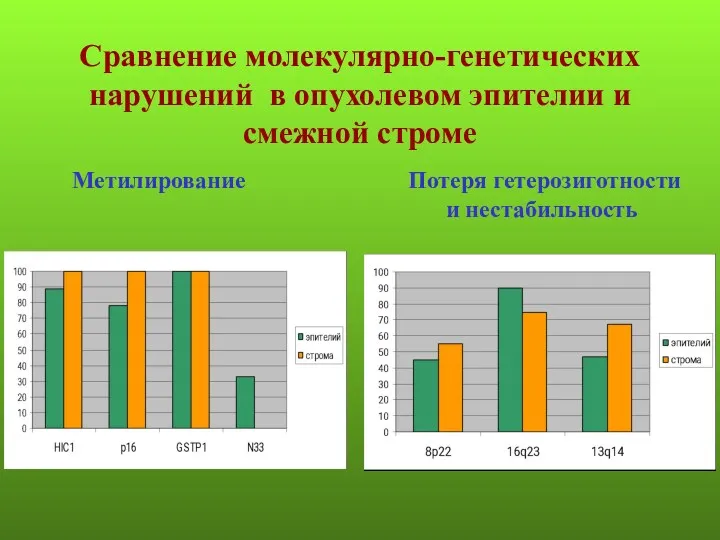
Сравнение молекулярно-генетических нарушений в опухолевом эпителии и смежной строме
Метилирование
Потеря гетерозиготности
и
Сравнение молекулярно-генетических нарушений в опухолевом эпителии и смежной строме
Метилирование
Потеря гетерозиготности
и

Система маркеров аномального метилирования для ранней дифференциальной диагностики опухолей предстательной железы
Система маркеров аномального метилирования для ранней дифференциальной диагностики опухолей предстательной железы

Маркеры метилирования, определяемые на ранних стадиях канцерогенеза.
Маркеры метилирования, определяемые на ранних стадиях канцерогенеза.
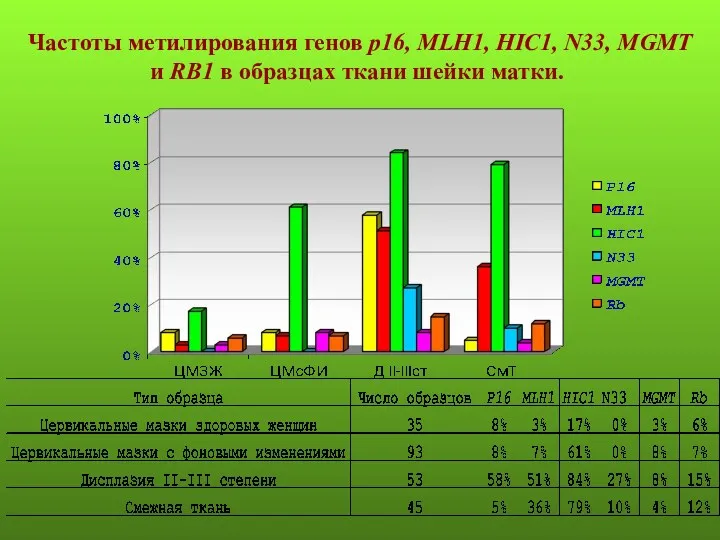
Частоты метилирования генов р16, MLH1, HIC1, N33, MGMT и RB1 в
Частоты метилирования генов р16, MLH1, HIC1, N33, MGMT и RB1 в
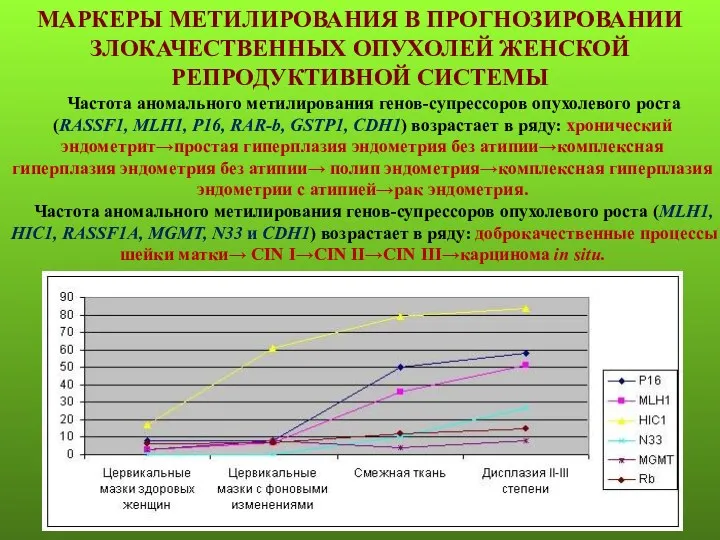
МАРКЕРЫ МЕТИЛИРОВАНИЯ В ПРОГНОЗИРОВАНИИ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ ЖЕНСКОЙ РЕПРОДУКТИВНОЙ СИСТЕМЫ
Частота аномального
МАРКЕРЫ МЕТИЛИРОВАНИЯ В ПРОГНОЗИРОВАНИИ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ ЖЕНСКОЙ РЕПРОДУКТИВНОЙ СИСТЕМЫ
Частота аномального

Инактивация посредством гиперметилирования ключевых генов-супрессоров сопровождается гипометилированием и активацией целого ряда
Инактивация посредством гиперметилирования ключевых генов-супрессоров сопровождается гипометилированием и активацией целого ряда

Аномальное деметилирование гена IGF2 в образцах опухолей.
Дорожка 18 – положительный контроль
Аномальное деметилирование гена IGF2 в образцах опухолей.
Дорожка 18 – положительный контроль
Свободная ДНК в периферической крови онкологического больного составляет около 200
Свободная ДНК в периферической крови онкологического больного составляет около 200

1 – отрицательный контроль; 2 и 7 – положительный контроль (негидролизованная
1 – отрицательный контроль; 2 и 7 – положительный контроль (негидролизованная
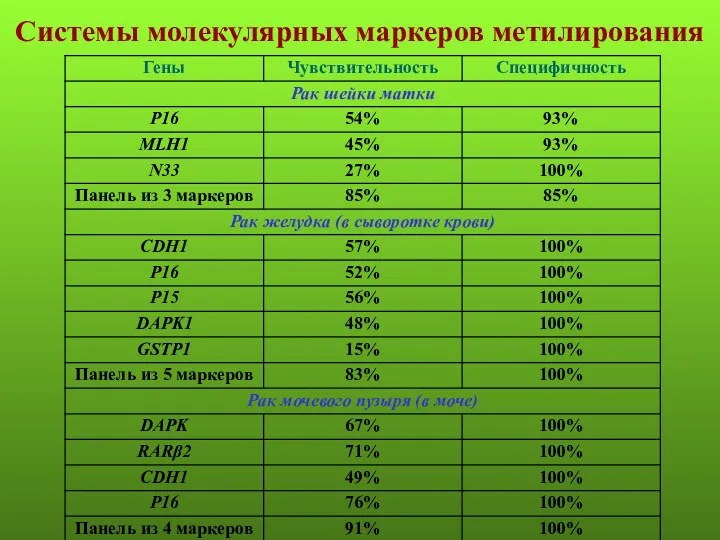
Системы молекулярных маркеров метилирования
Системы молекулярных маркеров метилирования
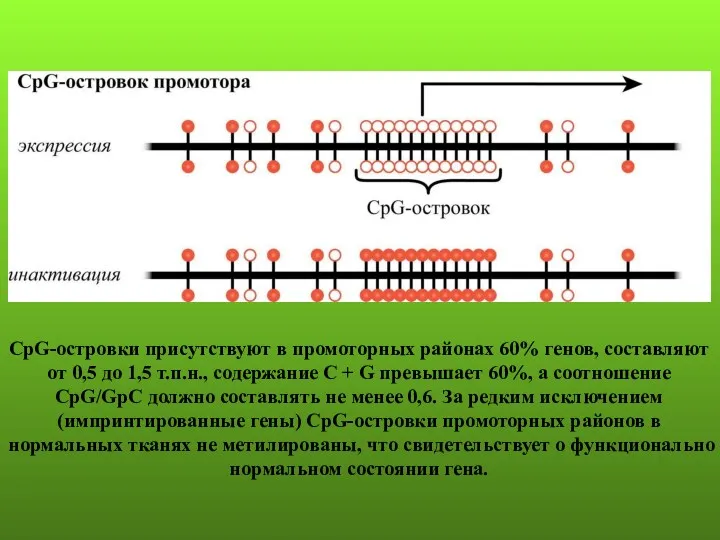
CpG-островки присутствуют в промоторных районах 60% генов, составляют от 0,5 до
CpG-островки присутствуют в промоторных районах 60% генов, составляют от 0,5 до

Карта метилирования
CpG-островка гена BIN1
Карта метилирования
CpG-островка гена BIN1

Промоторы достаточно большого числа генов имеют CpG-островки протяженностью свыше 1000
Промоторы достаточно большого числа генов имеют CpG-островки протяженностью свыше 1000

Спектр геномной локализации продуктов непредвзятого скрининга
Выявлено 26 новых локусов, аномально метилированных
Спектр геномной локализации продуктов непредвзятого скрининга
Выявлено 26 новых локусов, аномально метилированных

ФОРМИРОВАНИЕ СИСТЕМ МАРКЕРОВ МЕТИЛИРОВАНИЯ ДЛЯ ДИАГНОСТИКИ РМЖ
Для диагностики рака молочной железы
ФОРМИРОВАНИЕ СИСТЕМ МАРКЕРОВ МЕТИЛИРОВАНИЯ ДЛЯ ДИАГНОСТИКИ РМЖ
Для диагностики рака молочной железы

Метилирование ДНК является ценным биомаркером
для диагностики рака.
1. Целый ряд
Метилирование ДНК является ценным биомаркером
для диагностики рака.
1. Целый ряд

Метилирование промоторных районов генов RASSF и Р16 достоверно чаще происходит
Метилирование промоторных районов генов RASSF и Р16 достоверно чаще происходит
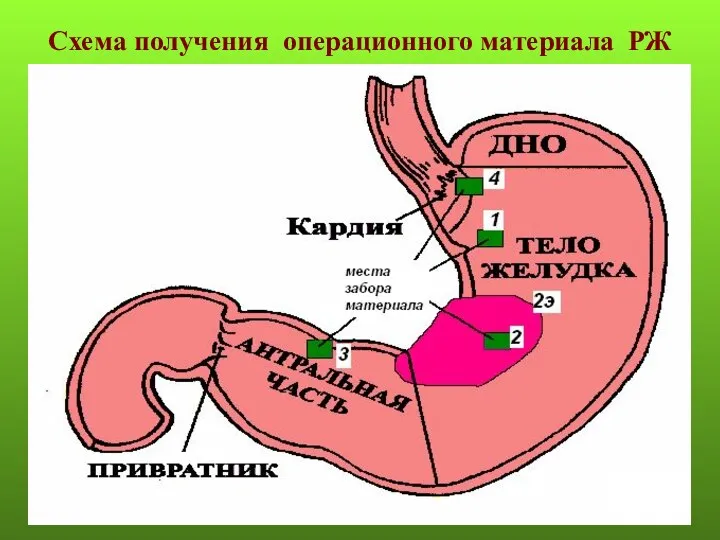
Схема получения операционного материала РЖ
Схема получения операционного материала РЖ
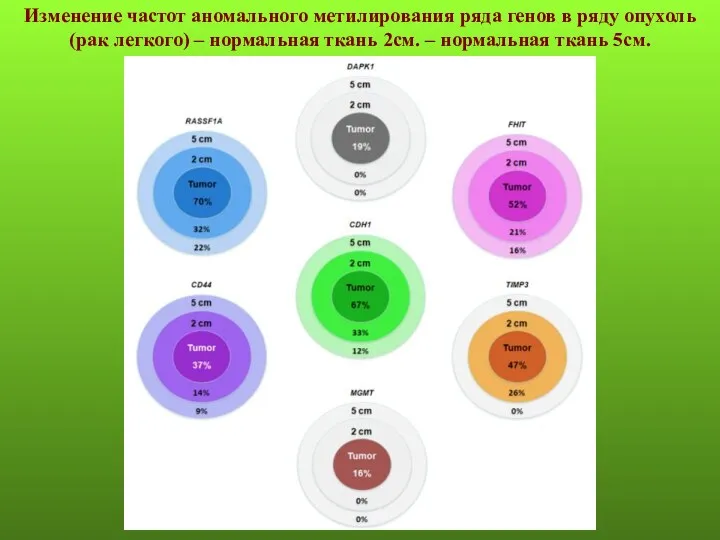
Изменение частот аномального метилирования ряда генов в ряду опухоль (рак легкого)
Изменение частот аномального метилирования ряда генов в ряду опухоль (рак легкого)

Метилирование ДНК, как диагностический маркер онкологического заболевания, имеет ряд преимуществ перед
Метилирование ДНК, как диагностический маркер онкологического заболевания, имеет ряд преимуществ перед
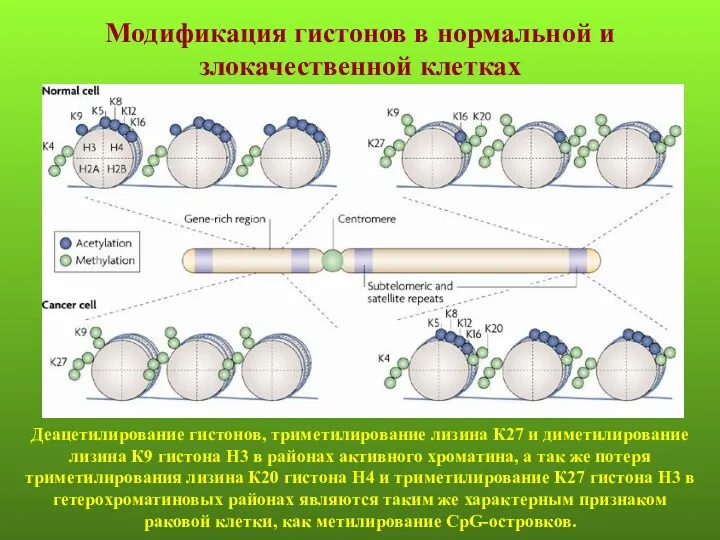
Деацетилирование гистонов, триметилирование лизина К27 и диметилирование лизина К9 гистона Н3
Деацетилирование гистонов, триметилирование лизина К27 и диметилирование лизина К9 гистона Н3

Патология генов, вовлеченных в эпигенетическую регуляцию, и канцерогенез
Патология генов, вовлеченных в эпигенетическую регуляцию, и канцерогенез

miРНК И КАНЦЕРОГЕНЕЗ
miРНК часто локализуются в районах фрагильных участков, в минимальных
miРНК И КАНЦЕРОГЕНЕЗ
miРНК часто локализуются в районах фрагильных участков, в минимальных

Ломкие сайты и картированные miРНК
113 ломких сайтов и 186 miR. 61
Ломкие сайты и картированные miРНК
113 ломких сайтов и 186 miR. 61
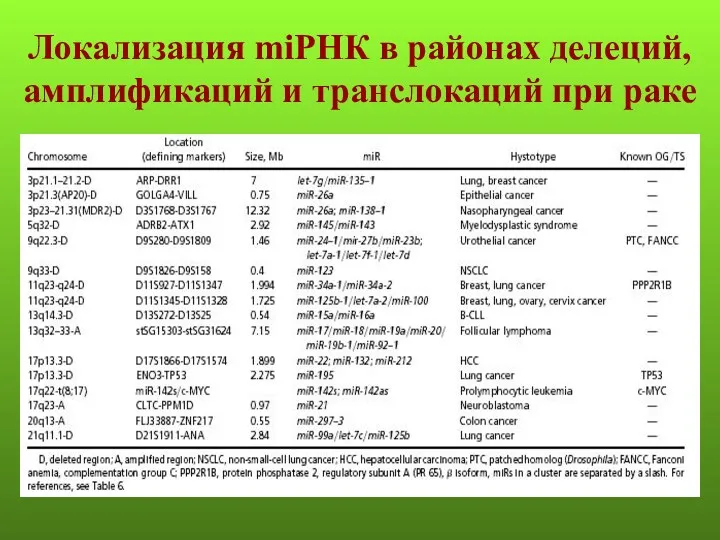
Локализация miРНК в районах делеций, амплификаций и транслокаций при раке
Локализация miРНК в районах делеций, амплификаций и транслокаций при раке

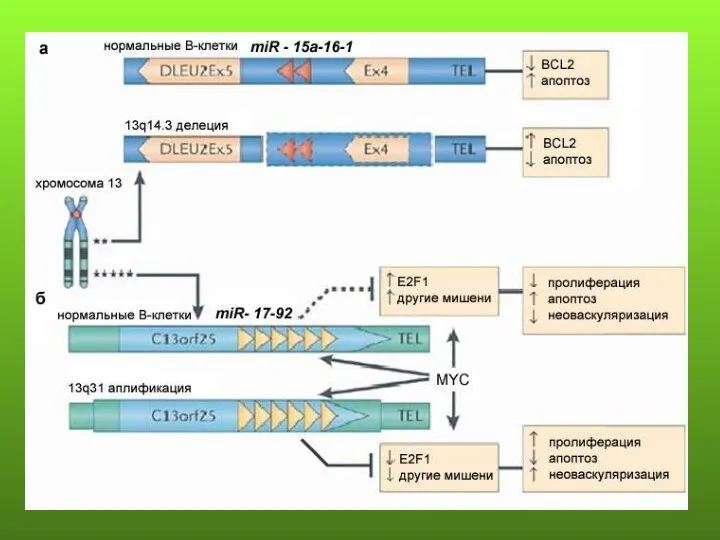
Хронический В-клеточный лимфолейкоз – минимальный район перекрывания делеций (~65% случаев)
Хронический В-клеточный лимфолейкоз – минимальный район перекрывания делеций (~65% случаев)

Клеточные онкогены K-RAS и MYC имеют множественные сайты комплементации с
Клеточные онкогены K-RAS и MYC имеют множественные сайты комплементации с


miРНК могут действовать как онкогены и гены-супрессоры. Причем профиль экспрессии miРНК
miРНК могут действовать как онкогены и гены-супрессоры. Причем профиль экспрессии miРНК

МОЛЕКУЛЯРНЫЕ МЕХАНИЗМЫ ДЕЙСТВИЯ нкРНК ПРИ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЯХ
МОЛЕКУЛЯРНЫЕ МЕХАНИЗМЫ ДЕЙСТВИЯ нкРНК ПРИ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЯХ
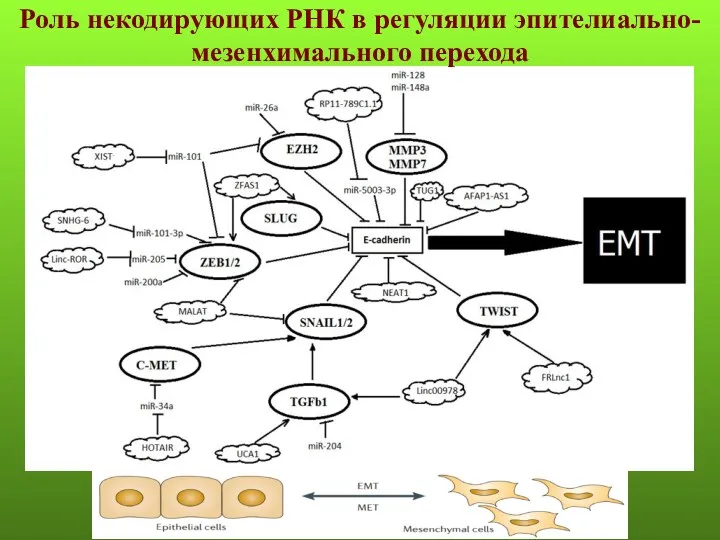
Роль некодирующих РНК в регуляции эпителиально-мезенхимального перехода
Роль некодирующих РНК в регуляции эпителиально-мезенхимального перехода

Локализация miРНК, вовлеченных в канцерогенез различных органов и рак
Локализация miРНК, вовлеченных в канцерогенез различных органов и рак
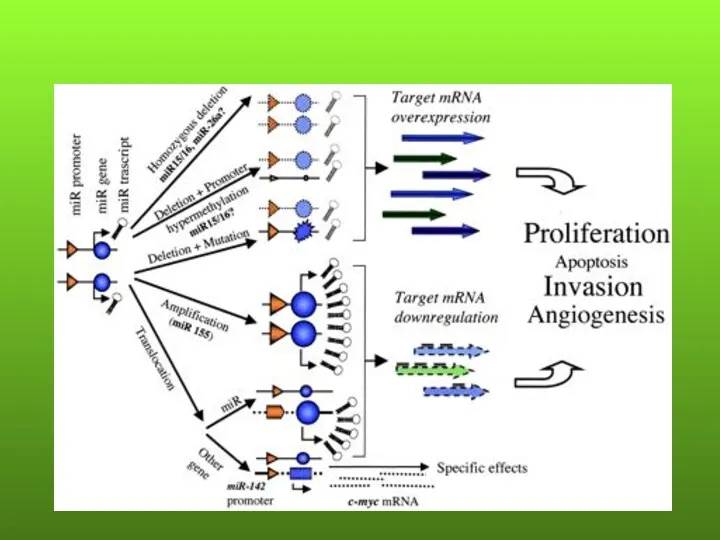
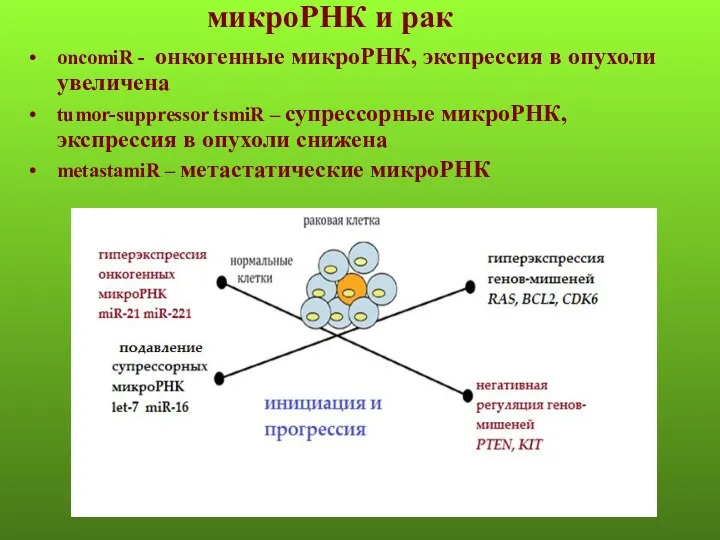
микроРНК и рак
oncomiR - онкогенные микроРНК, экспрессия в опухоли увеличена
tumor-suppressor tsmiR
микроРНК и рак
oncomiR - онкогенные микроРНК, экспрессия в опухоли увеличена
tumor-suppressor tsmiR
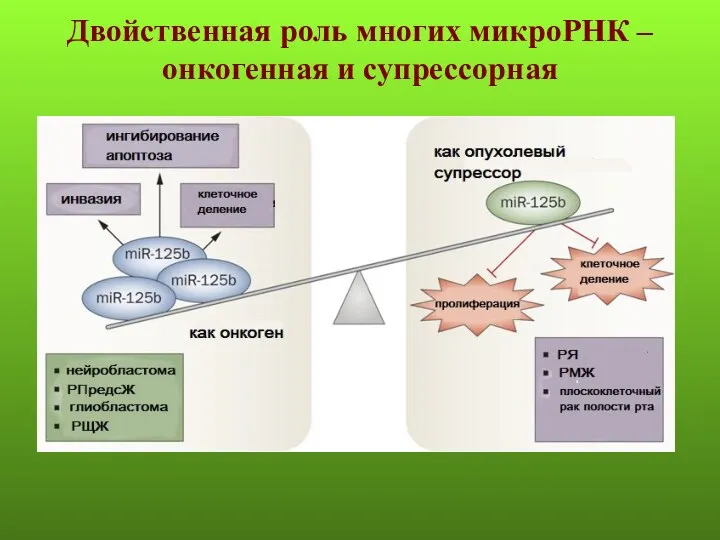
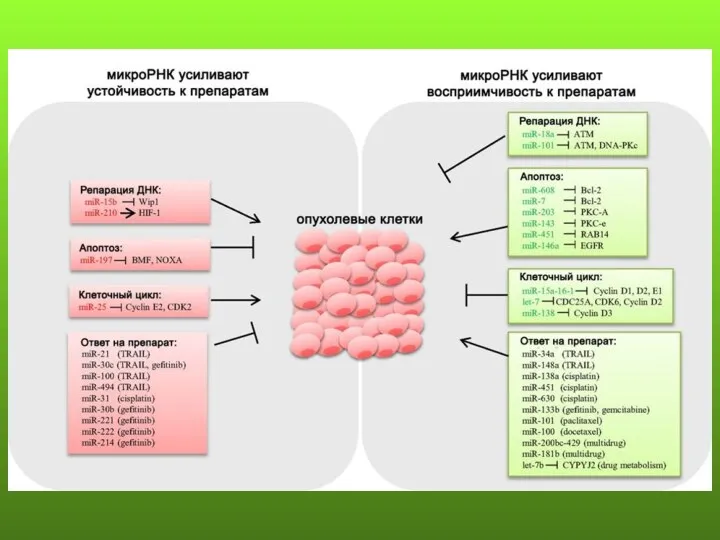
Двойственная роль многих микроРНК –
онкогенная и супрессорная
Двойственная роль многих микроРНК –
онкогенная и супрессорная
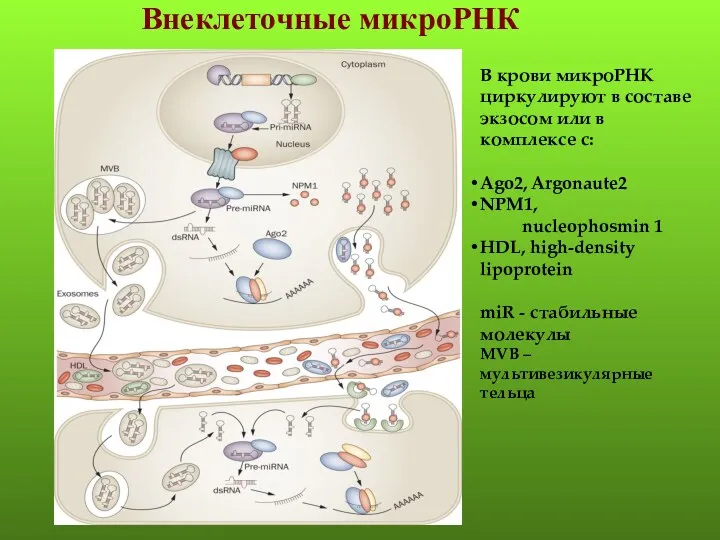
Внеклеточные микроРНК
В крови микроРНК циркулируют в составе экзосом или в комплексе
Внеклеточные микроРНК
В крови микроРНК циркулируют в составе экзосом или в комплексе
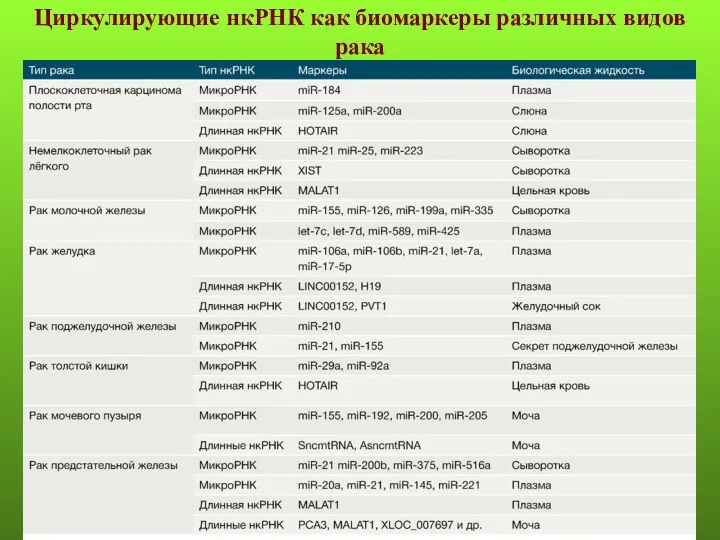
Циркулирующие нкРНК как биомаркеры различных видов рака
Циркулирующие нкРНК как биомаркеры различных видов рака

Одна микроРНК может регулировать сотни генов-мишеней. Аберрантная экспрессия микроРНК инициирует
Одна микроРНК может регулировать сотни генов-мишеней. Аберрантная экспрессия микроРНК инициирует
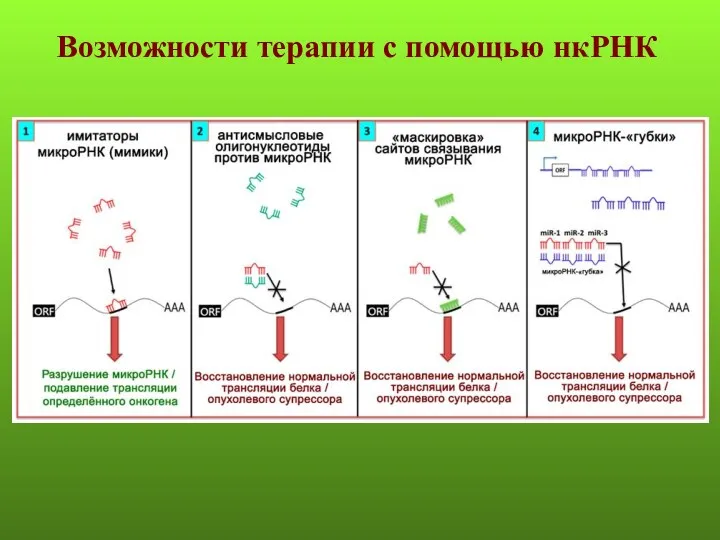
Возможности терапии с помощью нкРНК
Возможности терапии с помощью нкРНК
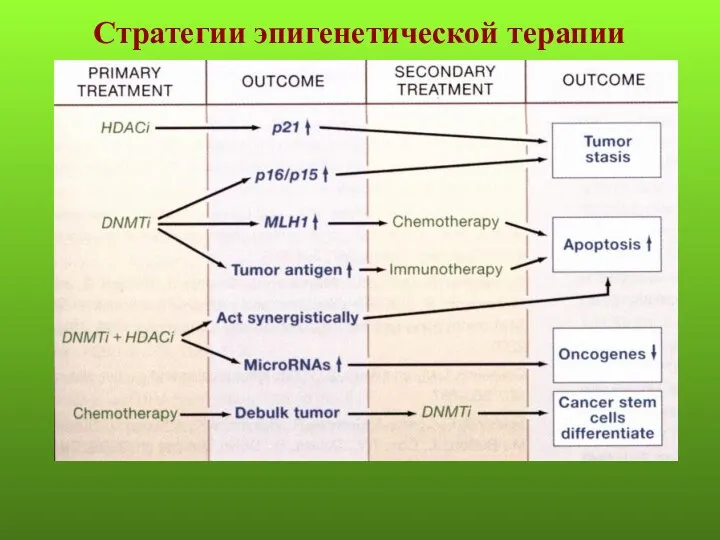
Стратегии эпигенетической терапии
Стратегии эпигенетической терапии

ХИМЕРНЫЕ ГЕНЫ
ХИМЕРНЫЕ ГЕНЫ
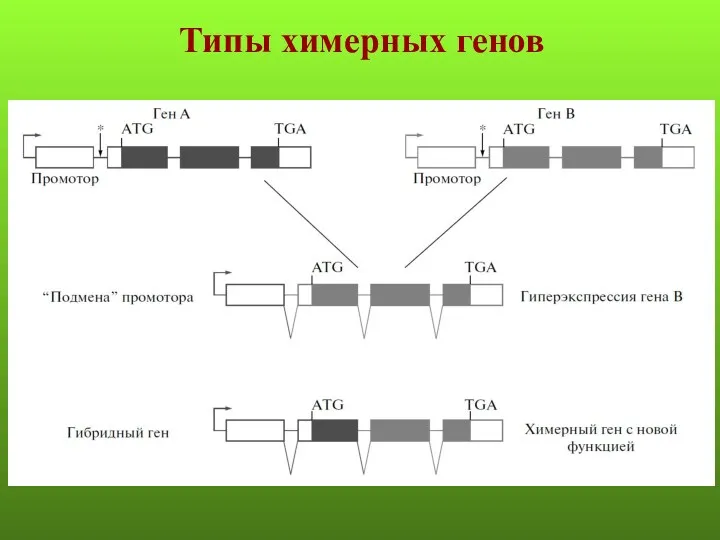
Типы химерных генов
Типы химерных генов
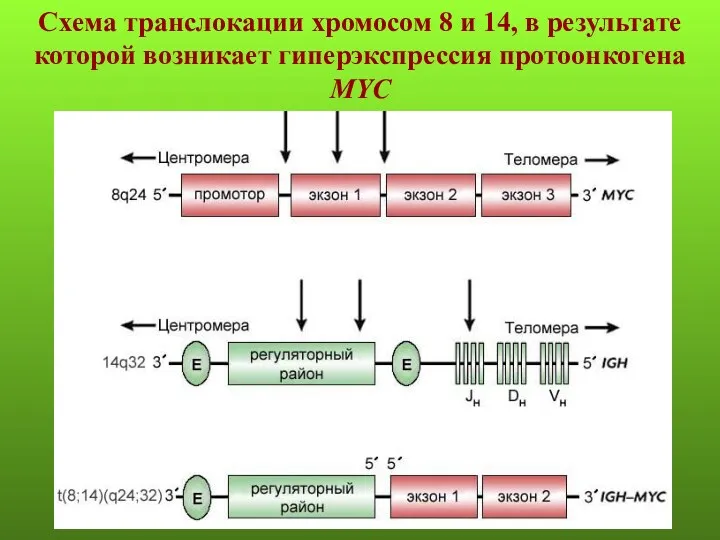
Схема транслокации хромосом 8 и 14, в результате которой возникает гиперэкспрессия
Схема транслокации хромосом 8 и 14, в результате которой возникает гиперэкспрессия
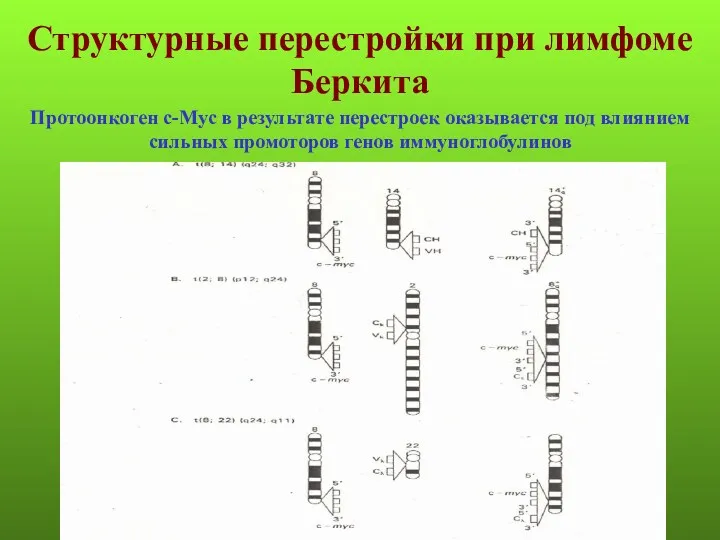
Структурные перестройки при лимфоме Беркита
Протоонкоген с-Myc в результате перестроек оказывается
Структурные перестройки при лимфоме Беркита
Протоонкоген с-Myc в результате перестроек оказывается
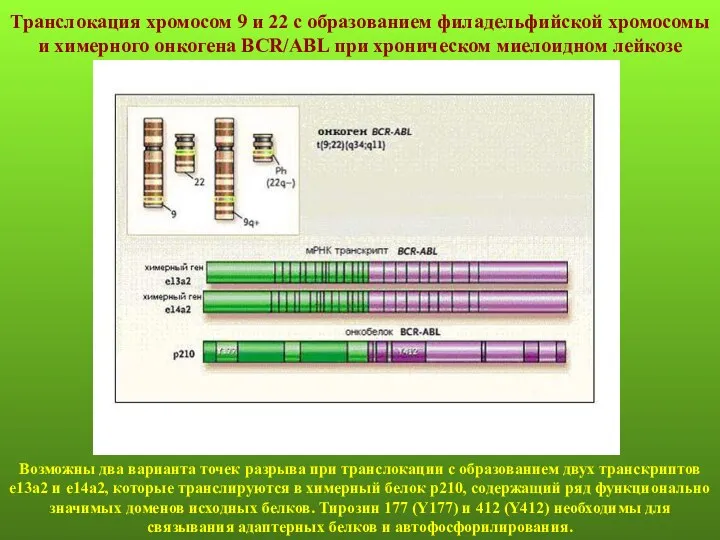
Транслокация хромосом 9 и 22 с образованием филадельфийской хромосомы и химерного
Транслокация хромосом 9 и 22 с образованием филадельфийской хромосомы и химерного
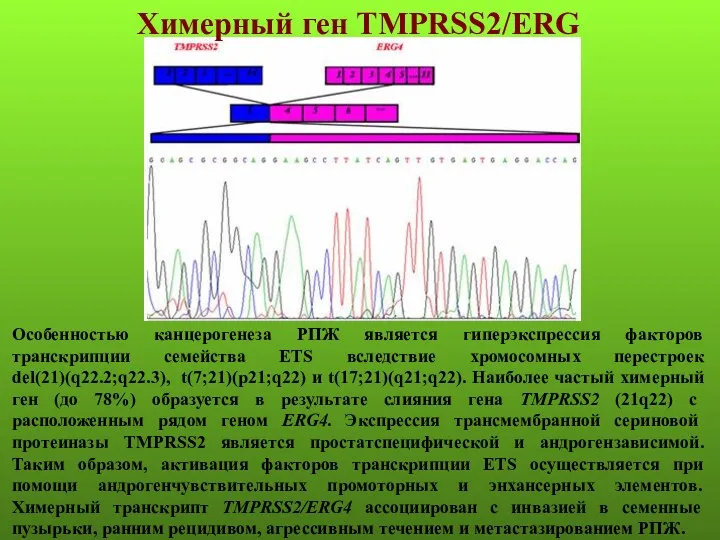
Химерный ген TMPRSS2/ERG
Особенностью канцерогенеза РПЖ является гиперэкспрессия факторов транскрипции семейства ETS
Химерный ген TMPRSS2/ERG
Особенностью канцерогенеза РПЖ является гиперэкспрессия факторов транскрипции семейства ETS
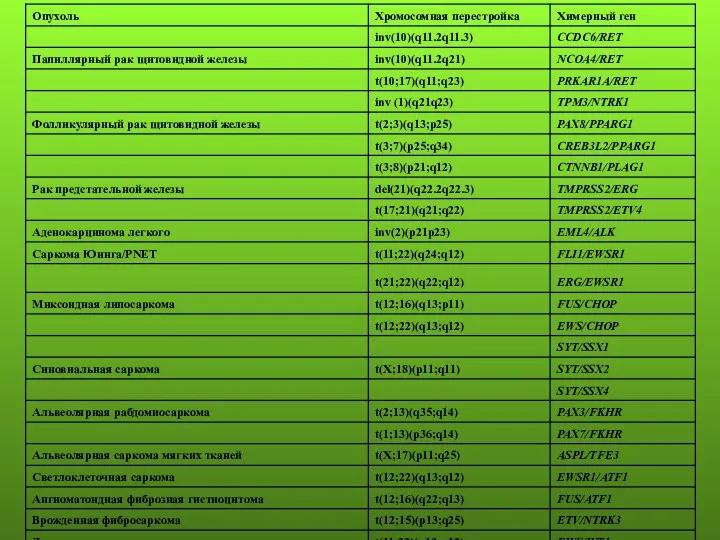
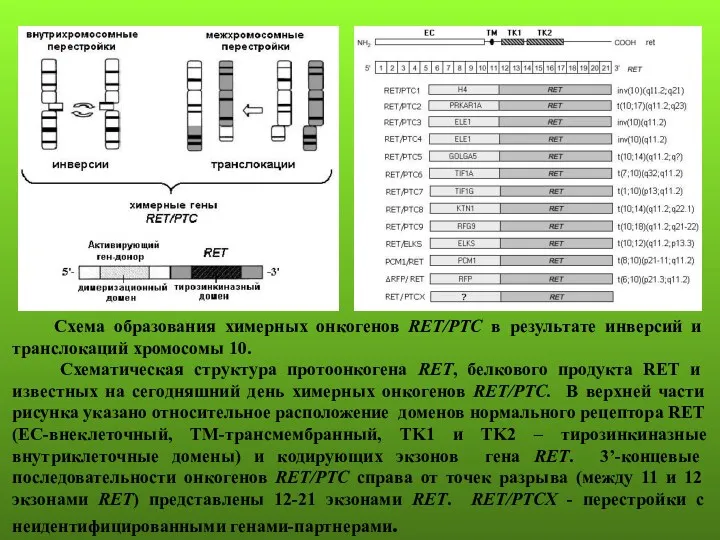
Схема образования химерных онкогенов RET/PTC в результате инверсий и транслокаций
Схема образования химерных онкогенов RET/PTC в результате инверсий и транслокаций
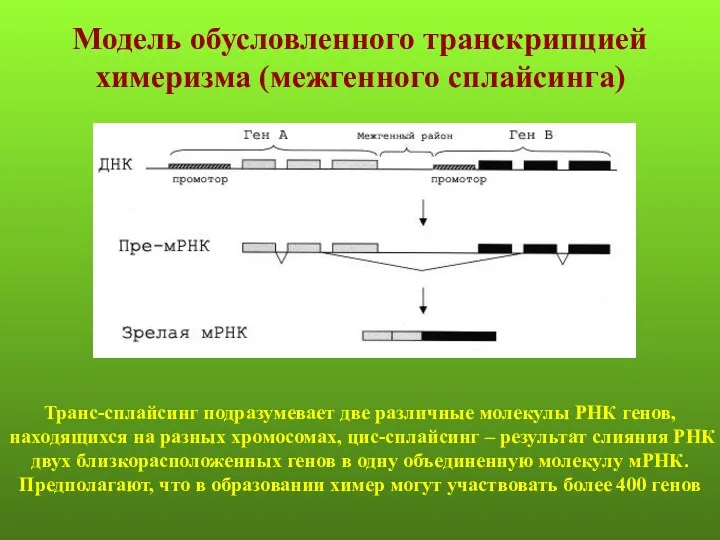
Модель обусловленного транскрипцией химеризма (межгенного сплайсинга)
Транс-сплайсинг подразумевает две различные молекулы
Модель обусловленного транскрипцией химеризма (межгенного сплайсинга)
Транс-сплайсинг подразумевает две различные молекулы
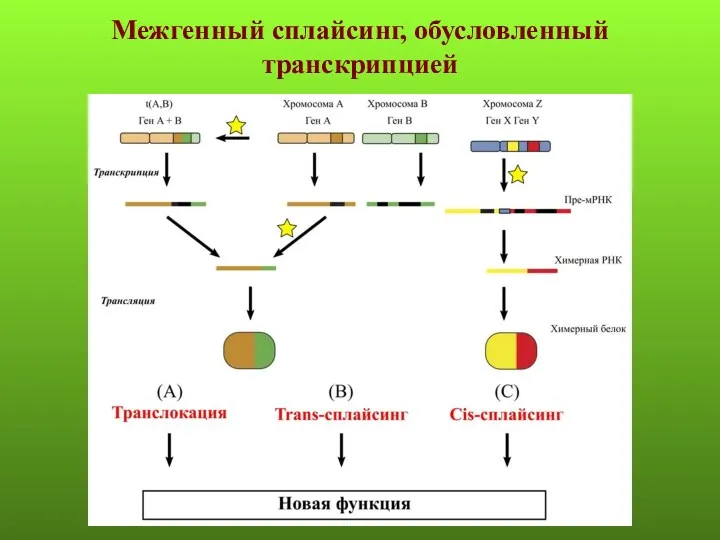
Межгенный сплайсинг, обусловленный транскрипцией
Межгенный сплайсинг, обусловленный транскрипцией

От 2 до 5% всех генов может быть вовлечено в процесс
От 2 до 5% всех генов может быть вовлечено в процесс

Химерные гены при раке предстательной железы:
В результате структурных перестроек:
TMPRSS2-ERG, TMPRSS2-ETV1, TMPRSS2-ETV4,
Химерные гены при раке предстательной железы:
В результате структурных перестроек:
TMPRSS2-ERG, TMPRSS2-ETV1, TMPRSS2-ETV4,

Химерные гены при ювенильной карциноме почки
В результате структурных перестроек:
MALAT1-TFEB, ASPSCR1-TFE3, PRCC-TFE3,
Химерные гены при ювенильной карциноме почки
В результате структурных перестроек:
MALAT1-TFEB, ASPSCR1-TFE3, PRCC-TFE3,
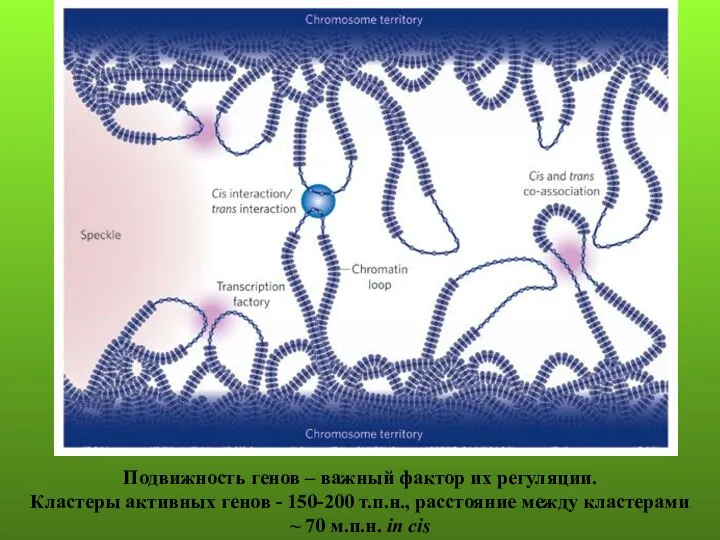
Подвижность генов – важный фактор их регуляции.
Кластеры активных генов - 150-200
Подвижность генов – важный фактор их регуляции.
Кластеры активных генов - 150-200

АЛЬТЕРНАТИВНЫЙ СПЛАЙСИНГ И ПАТОЛОГИЯ У ЧЕЛОВЕКА
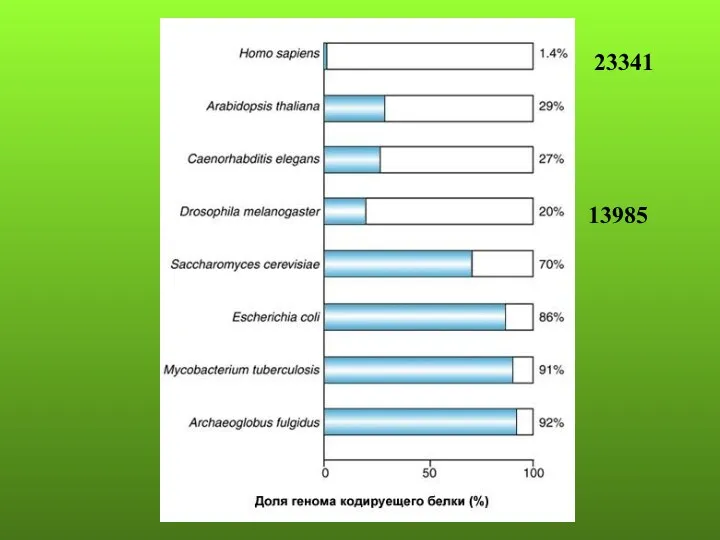
13985
23341
13985
23341

Около 70% генов человека могут подвергаться альтернативному сплайсингу, что вносит
Около 70% генов человека могут подвергаться альтернативному сплайсингу, что вносит
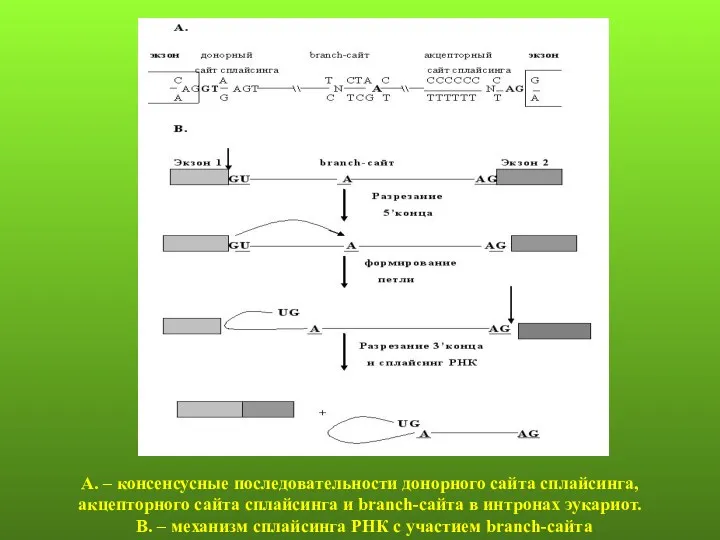
А. – консенсусные последовательности донорного сайта сплайсинга,
акцепторного сайта сплайсинга и
А. – консенсусные последовательности донорного сайта сплайсинга,
акцепторного сайта сплайсинга и

Четыре коротких последовательности нуклеотидов позволяют определить интрон: экзон-интронные границы в
Четыре коротких последовательности нуклеотидов позволяют определить интрон: экзон-интронные границы в
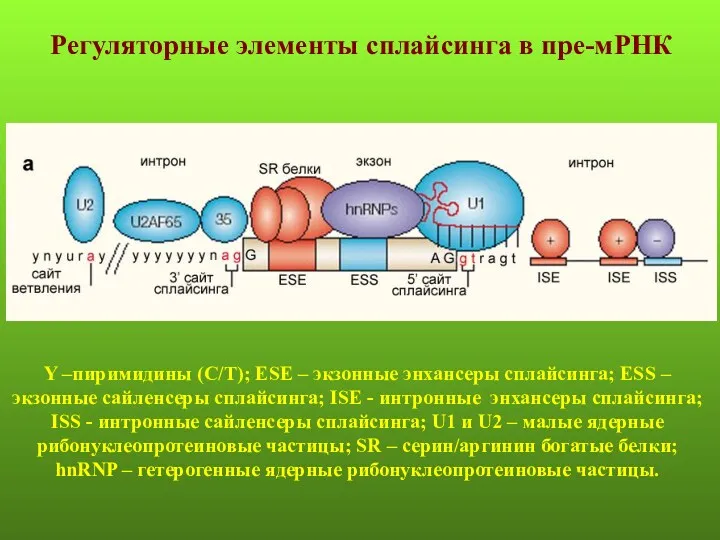
Y –пиримидины (С/Т); ESE – экзонные энхансеры сплайсинга; ESS – экзонные
Y –пиримидины (С/Т); ESE – экзонные энхансеры сплайсинга; ESS – экзонные

Экзонные энхансеры сплайсинга представляют собой сайты связывания обогащенных серином и
Экзонные энхансеры сплайсинга представляют собой сайты связывания обогащенных серином и
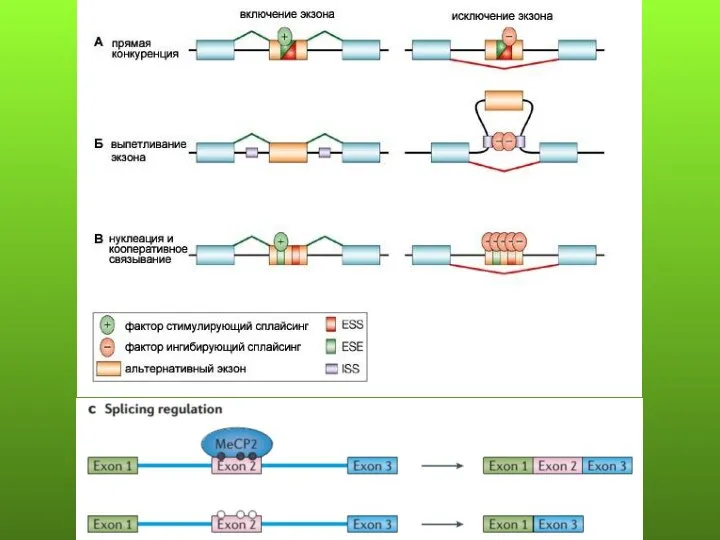
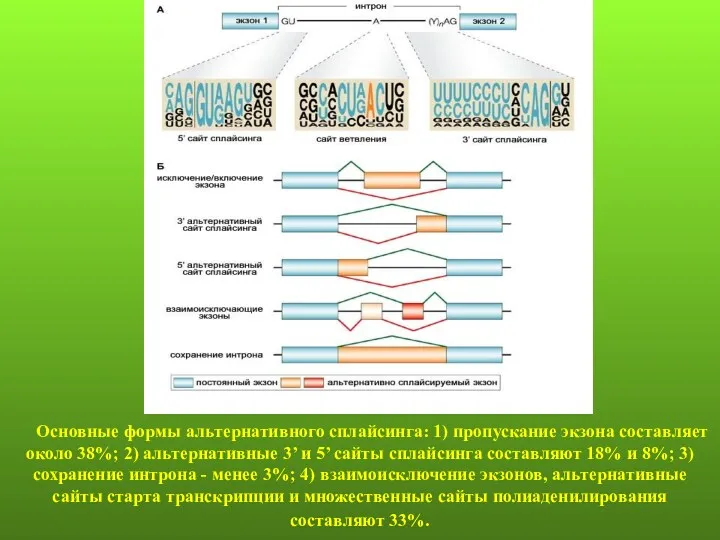
Основные формы альтернативного сплайсинга: 1) пропускание экзона составляет около 38%;
Основные формы альтернативного сплайсинга: 1) пропускание экзона составляет около 38%;
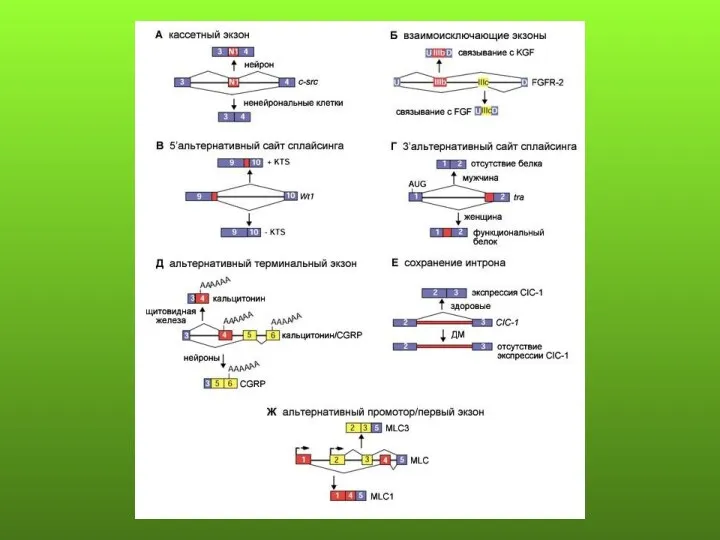

CD44 - мультифункциональный рецептор, вовлеченный в межклеточные взаимодействия, перемещение клеток, метаболизм
CD44 - мультифункциональный рецептор, вовлеченный в межклеточные взаимодействия, перемещение клеток, метаболизм

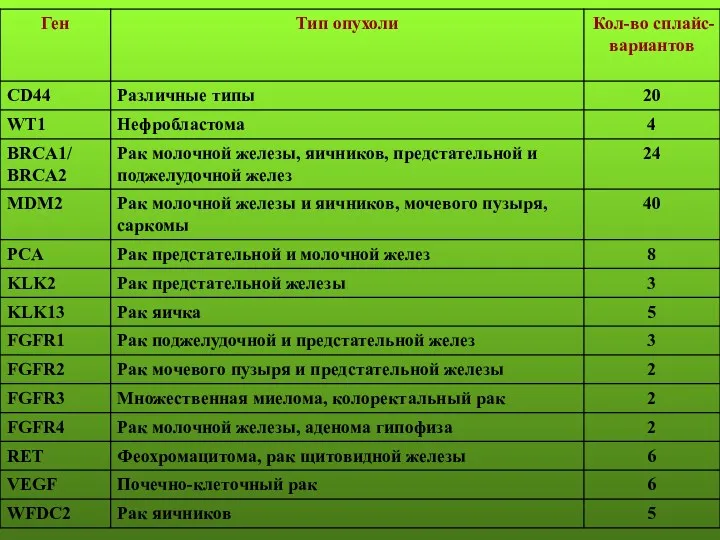
миРНК могут регулировать альтернативный сплайсинг. Их возможное участие согласуется с
миРНК могут регулировать альтернативный сплайсинг. Их возможное участие согласуется с
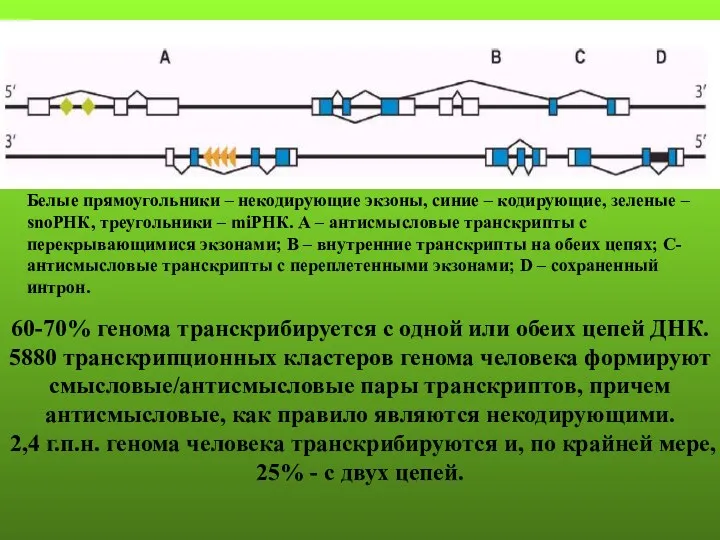
60-70% генома транскрибируется с одной или обеих цепей ДНК.
5880 транскрипционных кластеров
60-70% генома транскрибируется с одной или обеих цепей ДНК.
5880 транскрипционных кластеров


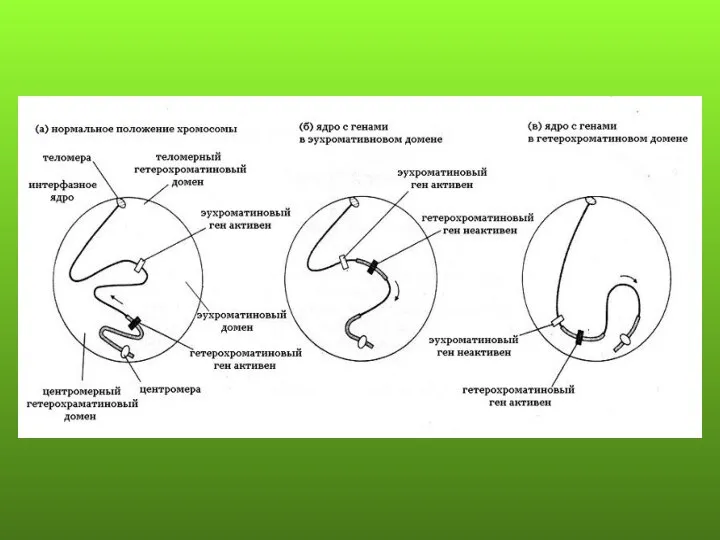
ПАТОЛОГИЯ ТИПА «ЭФФЕКТ ПОЛОЖЕНИЯ» У ЧЕЛОВЕКА

Эффект положения - нарушение или изменение уровня экспрессии гена в
Эффект положения - нарушение или изменение уровня экспрессии гена в

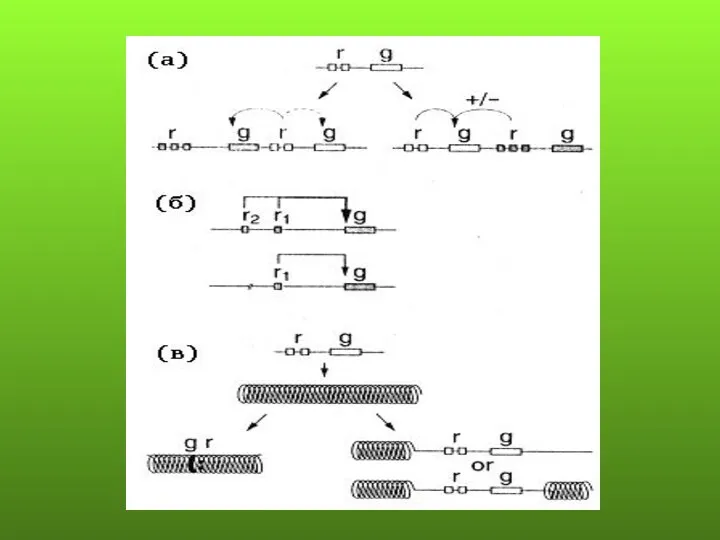
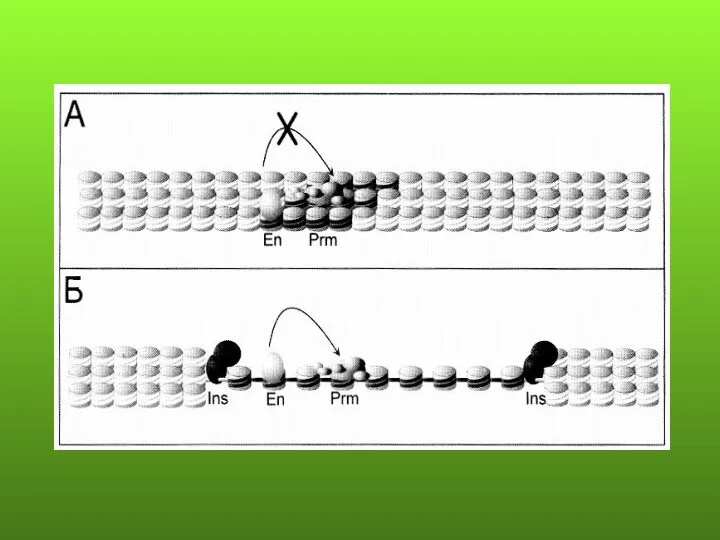
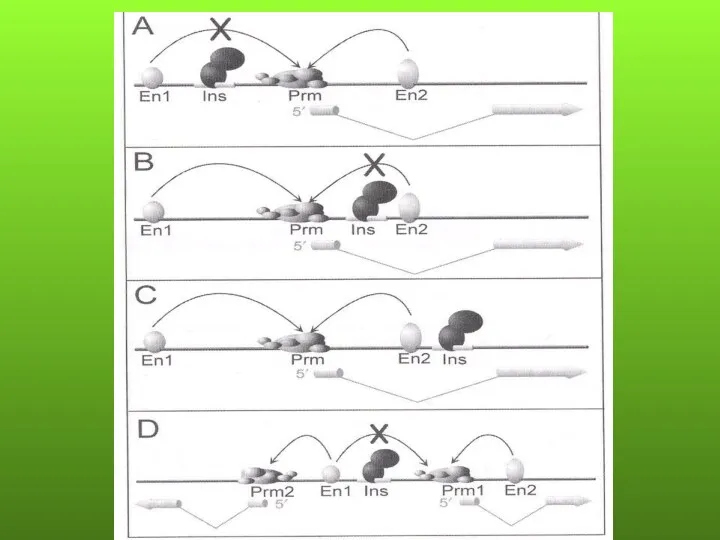
ПРЕДПОЛАГАЕМЫЕ МЕХАНИЗМЫ ЭФФЕКТА ПОЛОЖЕНИЯ
Перемещение гена из района с определенной структурой
ПРЕДПОЛАГАЕМЫЕ МЕХАНИЗМЫ ЭФФЕКТА ПОЛОЖЕНИЯ
Перемещение гена из района с определенной структурой


Доказанный эффект положения
Ген PAX6 - транскрипционный фактор. Хромосома 11p13.
Аниридия - врожденная
Доказанный эффект положения
Ген PAX6 - транскрипционный фактор. Хромосома 11p13.
Аниридия - врожденная

Ген PITX2 - транскрипционный фактор. Хромосома 4(q26). Синдром Ригера -
Ген PITX2 - транскрипционный фактор. Хромосома 4(q26). Синдром Ригера -
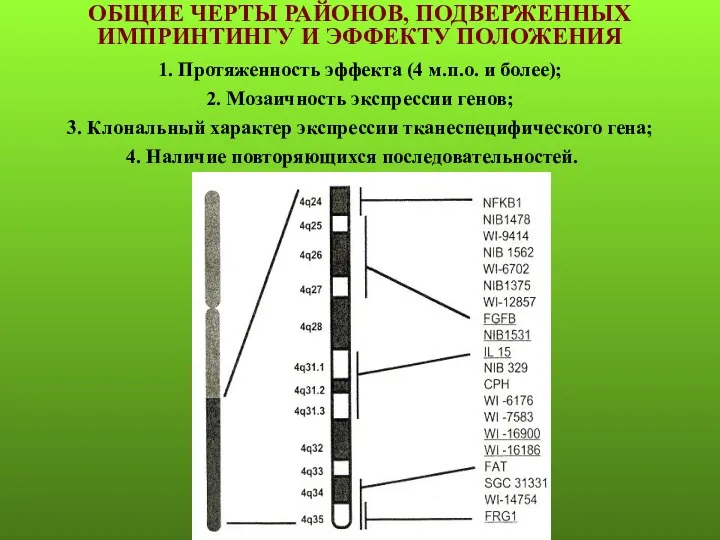
ОБЩИЕ ЧЕРТЫ РАЙОНОВ, ПОДВЕРЖЕННЫХ ИМПРИНТИНГУ И ЭФФЕКТУ ПОЛОЖЕНИЯ
1. Протяженность эффекта
ОБЩИЕ ЧЕРТЫ РАЙОНОВ, ПОДВЕРЖЕННЫХ ИМПРИНТИНГУ И ЭФФЕКТУ ПОЛОЖЕНИЯ
1. Протяженность эффекта



«белок-кодирующий район с определенными регуляторными последовательностями»
«транскрипционная единица» или «фрагмент ДНК,
«белок-кодирующий район с определенными регуляторными последовательностями»
«транскрипционная единица» или «фрагмент ДНК,

1860 – 1900х: «Ген – это дискретная единица наследственности» (Wilhelm Johannsen);
1860 – 1900х: «Ген – это дискретная единица наследственности» (Wilhelm Johannsen);
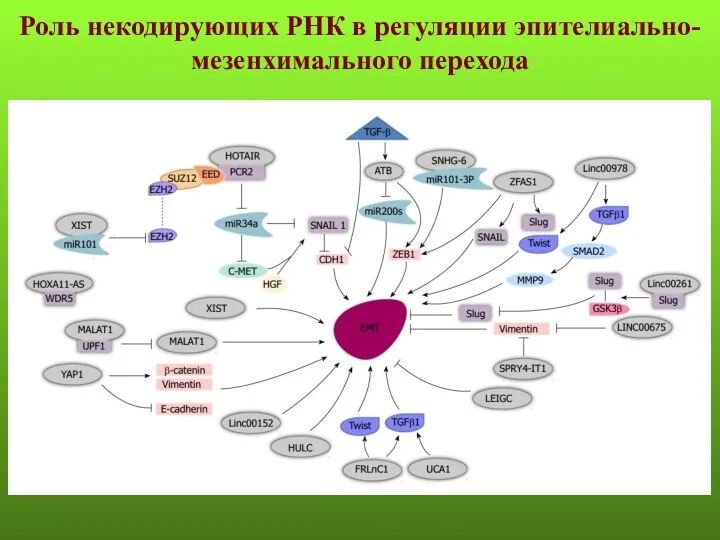
Роль некодирующих РНК в регуляции эпителиально-мезенхимального перехода
Роль некодирующих РНК в регуляции эпителиально-мезенхимального перехода

Система молекулярно-генетических маркеров для карцином желудка
Эпигенетические
Генетические
(структурные)
Делеции и МН
16q22.1 (CDH1)
17p13
Система молекулярно-генетических маркеров для карцином желудка
Эпигенетические
Генетические
(структурные)
Делеции и МН
16q22.1 (CDH1)
17p13

Различия в поведении молекулярных маркеров в клинических группах можно использовать
Различия в поведении молекулярных маркеров в клинических группах можно использовать

Методы поиска дифференциально метилированных CpG-островков
Рестрикционно-ориентированное геномное сканирование
Репрезентативно-дифференциальный анализ
Микрочипы
Метил-чувствительный фингерпринтинг
- наименьшая себестоимость
Методы поиска дифференциально метилированных CpG-островков
Рестрикционно-ориентированное геномное сканирование
Репрезентативно-дифференциальный анализ
Микрочипы
Метил-чувствительный фингерпринтинг
- наименьшая себестоимость
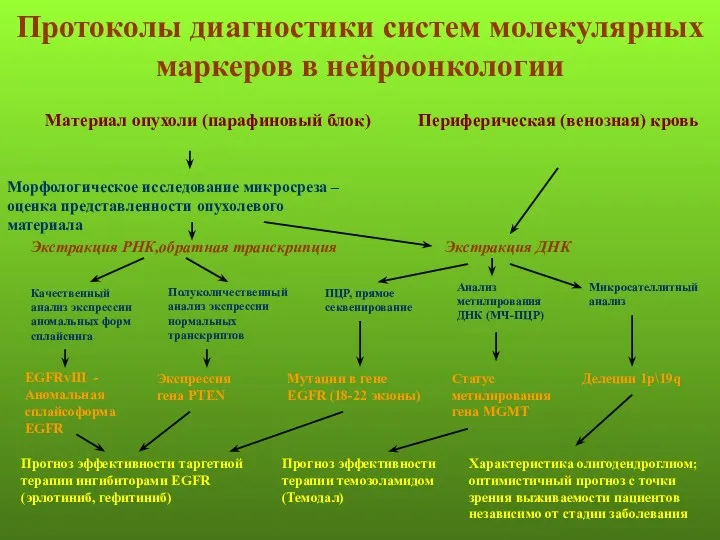
Протоколы диагностики систем молекулярных маркеров в нейроонкологии
Материал опухоли (парафиновый блок)
Периферическая (венозная)
Протоколы диагностики систем молекулярных маркеров в нейроонкологии
Материал опухоли (парафиновый блок)
Периферическая (венозная)
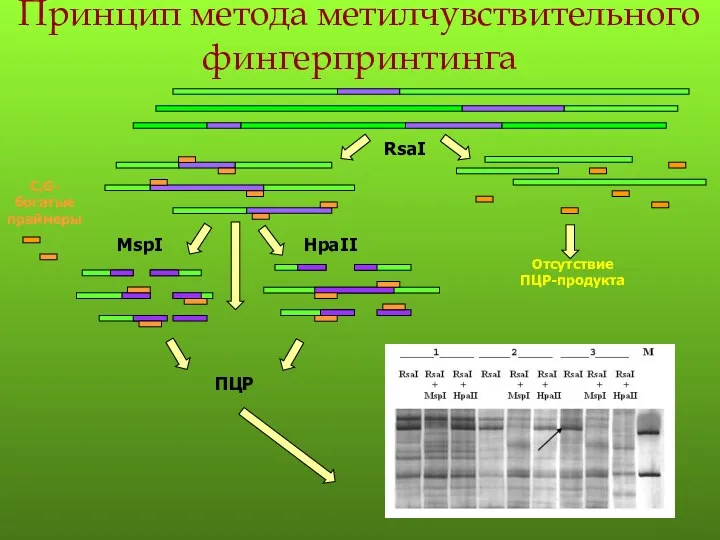
Принцип метода метилчувствительного фингерпринтинга
MspI
HpaII
Принцип метода метилчувствительного фингерпринтинга
MspI
HpaII
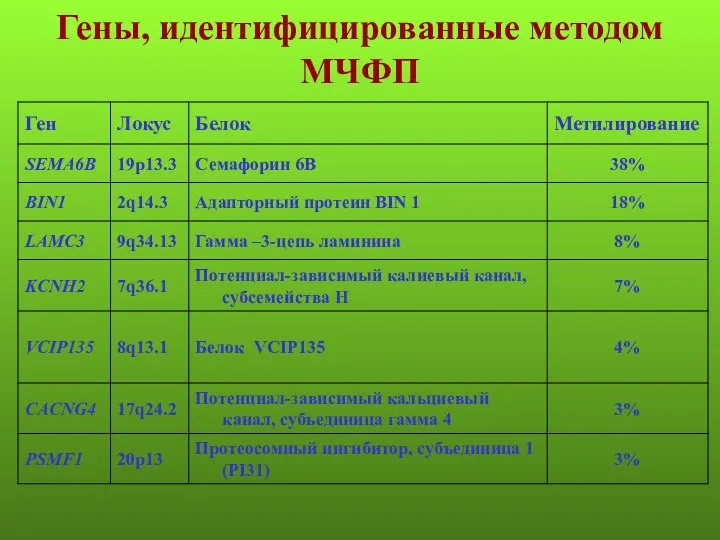
Гены, идентифицированные методом МЧФП
Гены, идентифицированные методом МЧФП

Принцип АИМС
Геномную ДНК обрабатывают метилчувствительной рестриктазой SmаI, которая формирует фрагменты с
Принцип АИМС
Геномную ДНК обрабатывают метилчувствительной рестриктазой SmаI, которая формирует фрагменты с
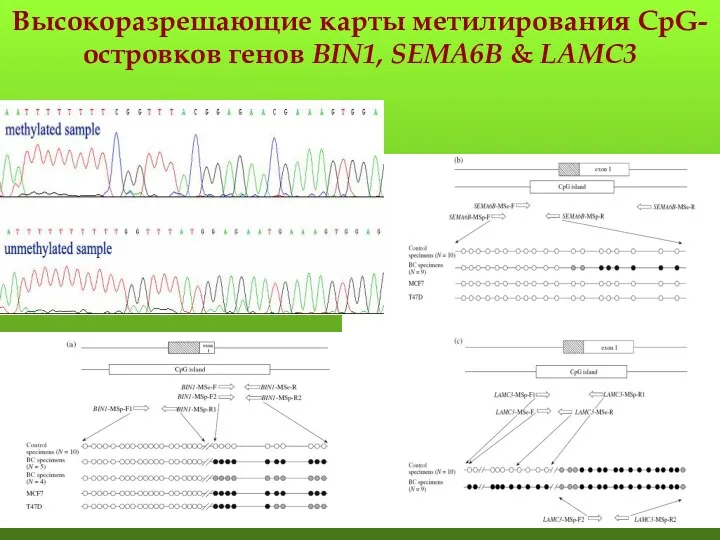
Высокоразрешающие карты метилирования CpG-островков генов BIN1, SEMA6B & LAMC3
Высокоразрешающие карты метилирования CpG-островков генов BIN1, SEMA6B & LAMC3
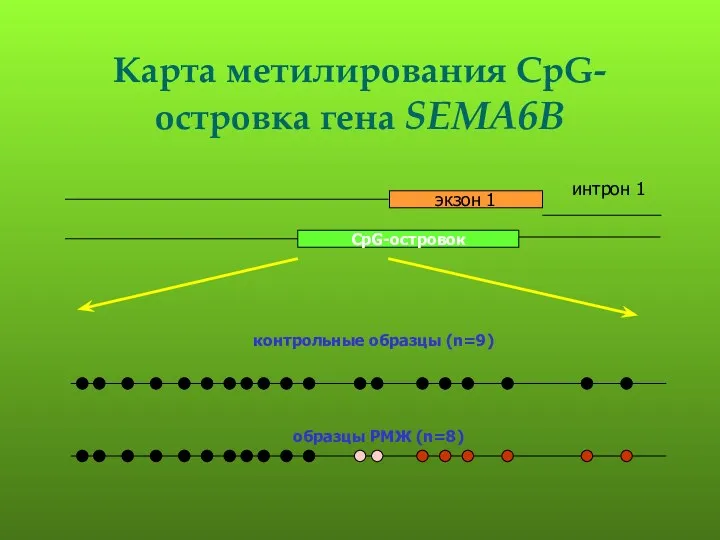
Карта метилирования CpG-островка гена SEMA6B
контрольные образцы (n=9)
образцы РМЖ (n=8)
Карта метилирования CpG-островка гена SEMA6B
контрольные образцы (n=9)
образцы РМЖ (n=8)


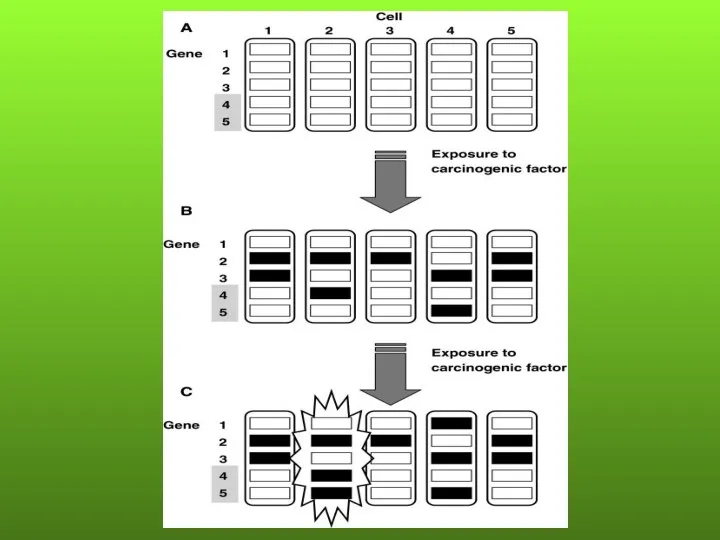
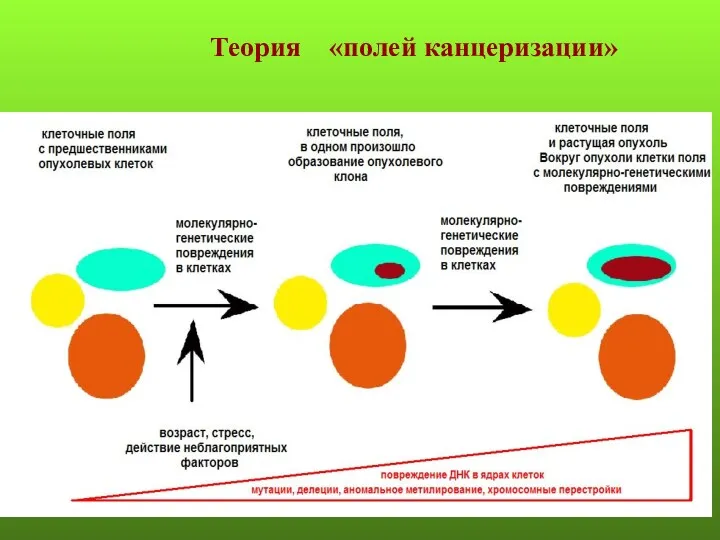
Теория «полей канцеризации»
Теория «полей канцеризации»
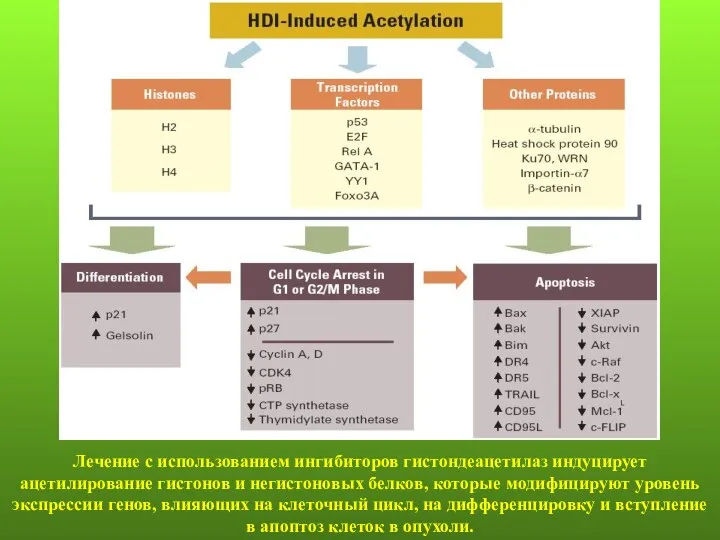
Лечение с использованием ингибиторов гистондеацетилаз индуцирует ацетилирование гистонов и негистоновых белков,
Лечение с использованием ингибиторов гистондеацетилаз индуцирует ацетилирование гистонов и негистоновых белков,

Вопросы и методы науки
Геномика
Эпигеномика
Транскриптомика
Протеомика
Сколько и каких изменений достаточно для классификации и
Вопросы и методы науки
Геномика
Эпигеномика
Транскриптомика
Протеомика
Сколько и каких изменений достаточно для классификации и
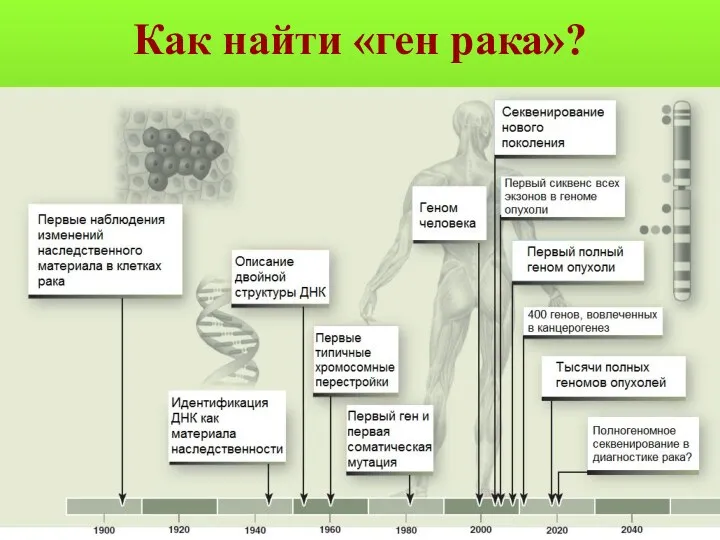
Как найти «ген рака»?
Как найти «ген рака»?

Сколько и какие гены несут мутации в опухоли?
До внедрения полногеномного секвенирования
Сколько и какие гены несут мутации в опухоли?
До внедрения полногеномного секвенирования

Соматические мутации в опухоли:
драйверные и пассажирские
Драйверные мутации возникают в ключевых
Соматические мутации в опухоли:
драйверные и пассажирские
Драйверные мутации возникают в ключевых

Соматические мутации в опухоли:
драйверные и пассажирские
Прочие соматические мутации, составляющие в
Соматические мутации в опухоли:
драйверные и пассажирские
Прочие соматические мутации, составляющие в
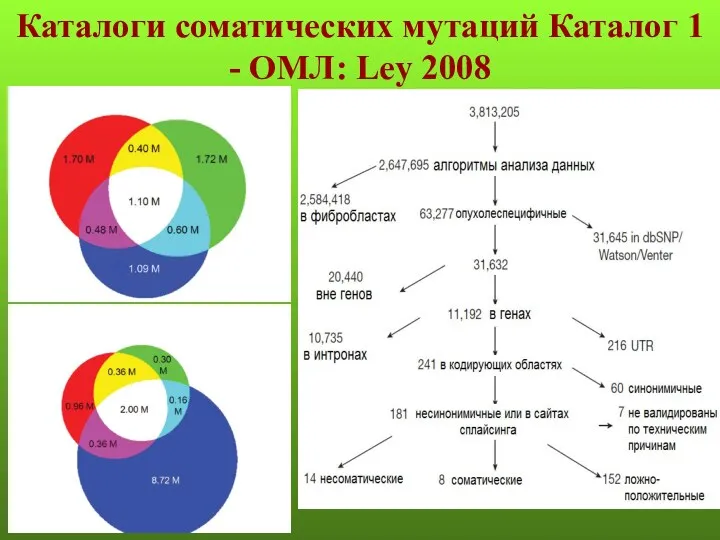
Каталоги соматических мутаций Каталог 1 - ОМЛ: Ley 2008
Каталоги соматических мутаций Каталог 1 - ОМЛ: Ley 2008

Каталог 2 - ОМЛ: Mardis 2009
Уровень ложноположительных результатов снижен с 96%
Каталог 2 - ОМЛ: Mardis 2009
Уровень ложноположительных результатов снижен с 96%

микроРНК – прогноз и лечение
Выживаемость – корреляция с экспрессией miR-21, miR-155,
микроРНК – прогноз и лечение
Выживаемость – корреляция с экспрессией miR-21, miR-155,

Методы анализа метилирования
1. Метилчувствительная ПЦР (Not1, Eag1, SacII, HpaII, HhaI)
аналитическая
Методы анализа метилирования
1. Метилчувствительная ПЦР (Not1, Eag1, SacII, HpaII, HhaI)
аналитическая
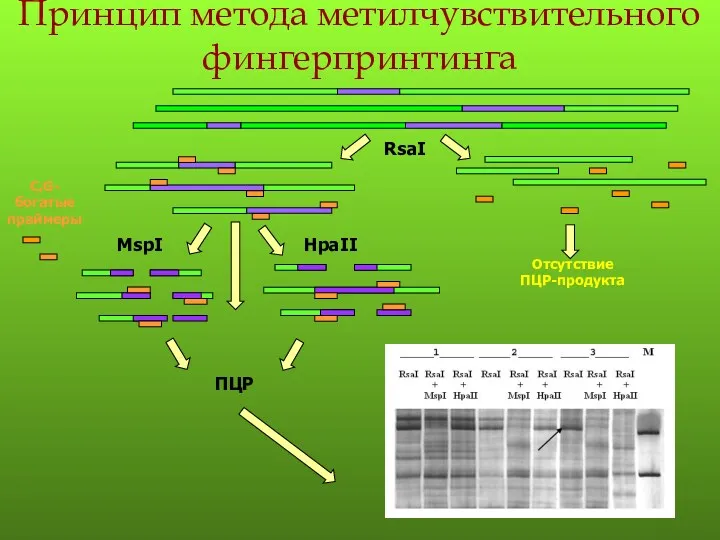
Принцип метода метилчувствительного фингерпринтинга
MspI
HpaII
Принцип метода метилчувствительного фингерпринтинга
MspI
HpaII
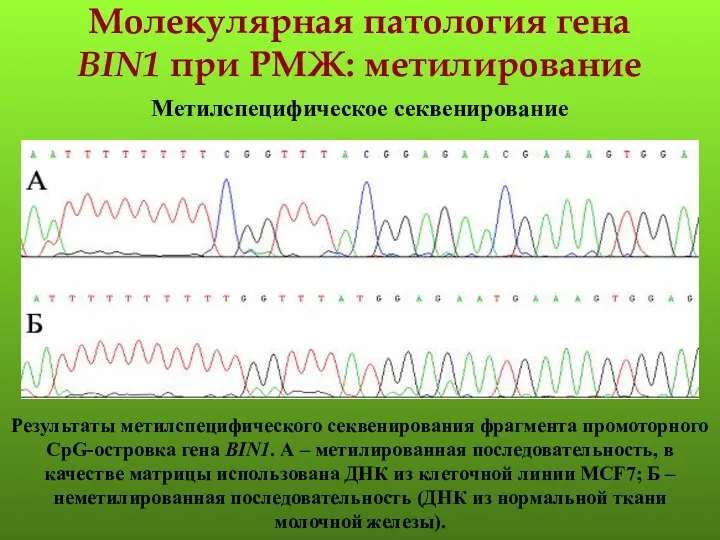
Молекулярная патология гена BIN1 при РМЖ: метилирование
Метилспецифическое секвенирование
Результаты метилспецифического секвенирования фрагмента
Молекулярная патология гена BIN1 при РМЖ: метилирование
Метилспецифическое секвенирование
Результаты метилспецифического секвенирования фрагмента

УЧАСТИЕ В КАНЦЕРОГЕНЕЗЕ НЕКОТОРЫХ ИЗ ВЫЯВЛЕННЫХ ГЕНОВ
УЧАСТИЕ В КАНЦЕРОГЕНЕЗЕ НЕКОТОРЫХ ИЗ ВЫЯВЛЕННЫХ ГЕНОВ
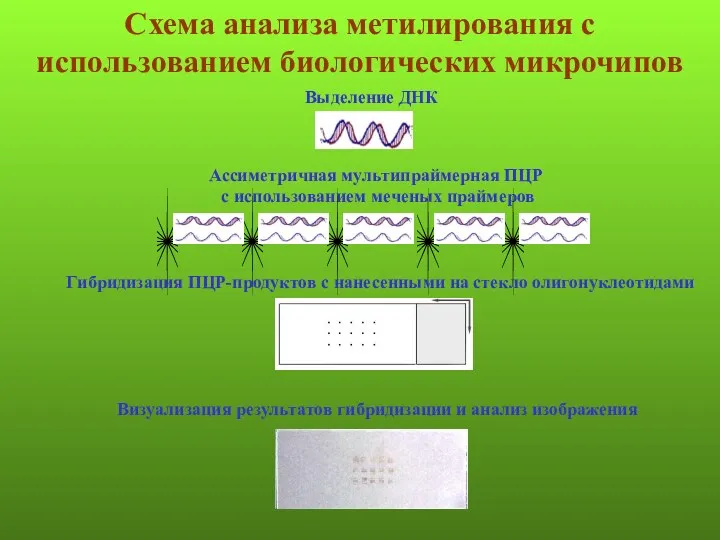
Схема анализа метилирования с использованием биологических микрочипов
Схема анализа метилирования с использованием биологических микрочипов
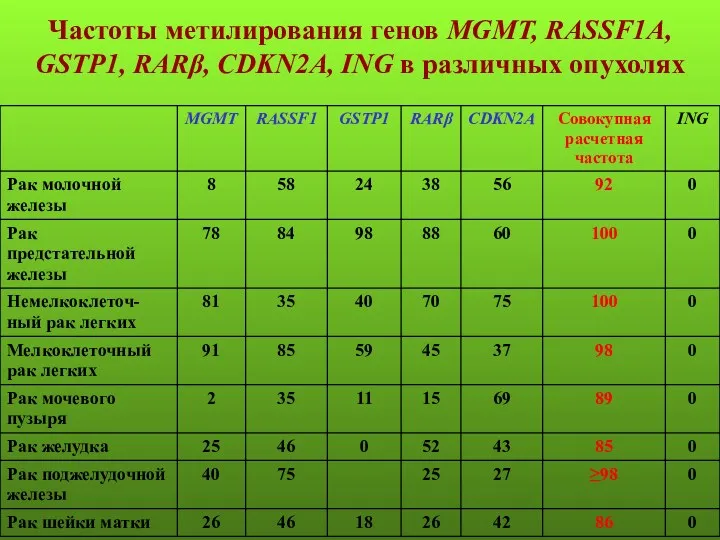
Частоты метилирования генов MGMT, RASSF1A, GSTP1, RARβ, CDKN2A, ING в различных
Частоты метилирования генов MGMT, RASSF1A, GSTP1, RARβ, CDKN2A, ING в различных

Определение метилирования генов, вовлеченных в канцерогенез, методом метилчувствительной ПЦР
Определение метилирования генов, вовлеченных в канцерогенез, методом метилчувствительной ПЦР
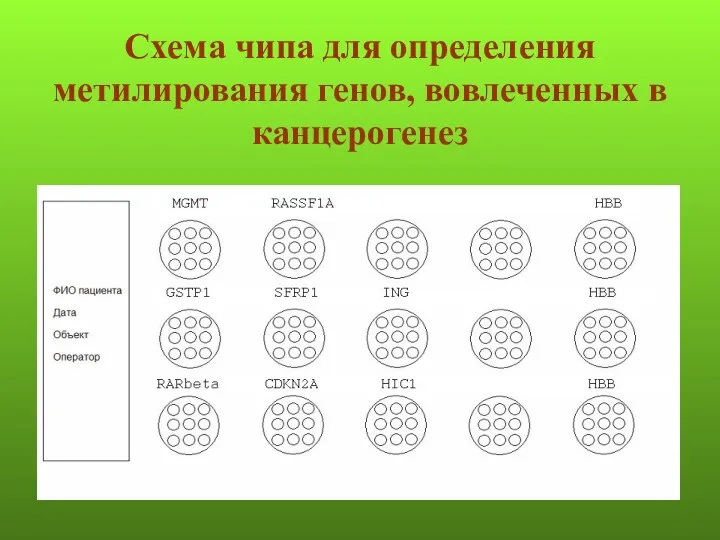
Схема чипа для определения метилирования генов, вовлеченных в канцерогенез
Схема чипа для определения метилирования генов, вовлеченных в канцерогенез
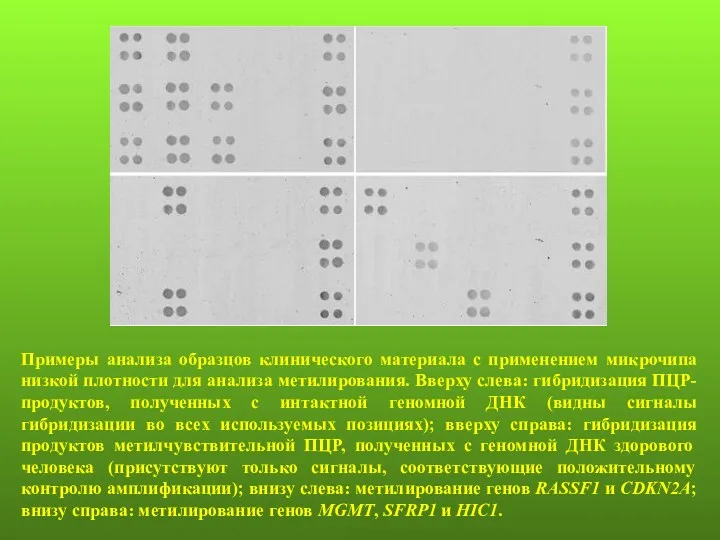
Примеры анализа образцов клинического материала с применением микрочипа низкой плотности для
Примеры анализа образцов клинического материала с применением микрочипа низкой плотности для

Среднее значение экспрессии let-7a, miR-155, miR-205 в образцах рака легкого
Среднее значение экспрессии let-7a, miR-155, miR-205 в образцах рака легкого

Примеры генов-супрессоров опухолевого роста,
CpG-островки которых могут быть метилированы при
Примеры генов-супрессоров опухолевого роста, CpG-островки которых могут быть метилированы при

Примеры генов-супрессоров опухолевого роста,
CpG-островки которых могут быть метилированы при различных злокачественных
Примеры генов-супрессоров опухолевого роста, CpG-островки которых могут быть метилированы при различных злокачественных

Метилирование позволяет предсказать поведение опухоли (эффективность терапии, метастазирование):
- метилирование WIT1
Метилирование позволяет предсказать поведение опухоли (эффективность терапии, метастазирование):
- метилирование WIT1
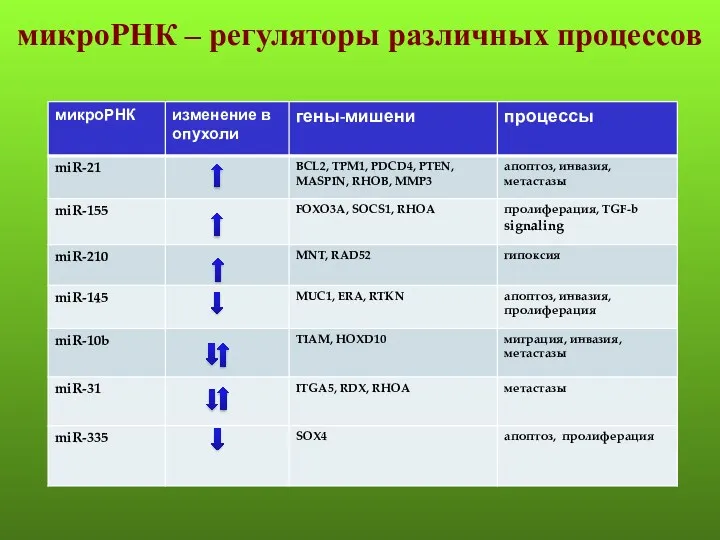
микроРНК – регуляторы различных процессов
микроРНК – регуляторы различных процессов

Лекарственные средства для аномальных эпигенетических событий
Ингибиторы ДНК-метилтрансфераз
5-Azacytidine Vidaza (лейкемия, МДС)
5-Aza-2’-deoxycytidine
Лекарственные средства для аномальных эпигенетических событий
Ингибиторы ДНК-метилтрансфераз
5-Azacytidine Vidaza (лейкемия, МДС)
5-Aza-2’-deoxycytidine
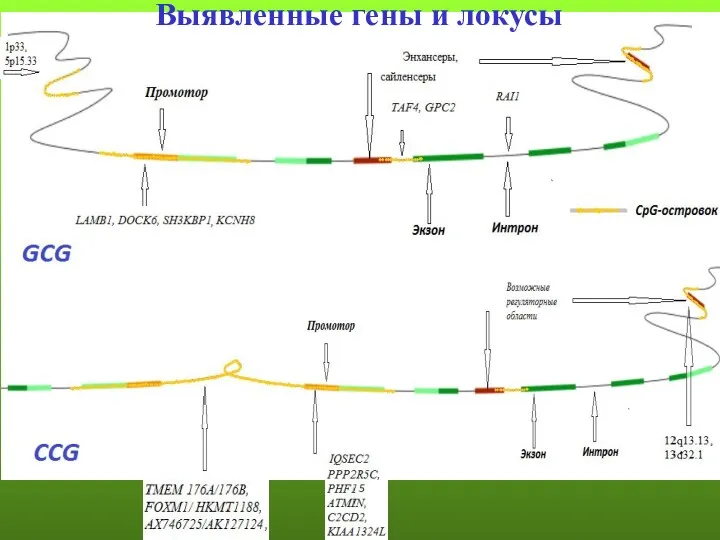
Выявленные гены и локусы
Выявленные гены и локусы
 Патология эмоциональной сферы. Расстройства влечений. Патология двигательной и волевой сферы
Патология эмоциональной сферы. Расстройства влечений. Патология двигательной и волевой сферы Выделительная система
Выделительная система Ауыз қуыс кілегей қабық ауруларына тағайындалатын дәрілік терпияның салыстырмалы сипаттамасы
Ауыз қуыс кілегей қабық ауруларына тағайындалатын дәрілік терпияның салыстырмалы сипаттамасы Дисциркуляторная энцефалопатия и болезнь мелких сосудов
Дисциркуляторная энцефалопатия и болезнь мелких сосудов Жүкті әйелдердегі ауруханадан тыс пневмония
Жүкті әйелдердегі ауруханадан тыс пневмония Анафилаксикалық шок кезіндегі жедел көмек
Анафилаксикалық шок кезіндегі жедел көмек Рак тела матки
Рак тела матки Краснуха - острая вирусная антропонозная инфекция
Краснуха - острая вирусная антропонозная инфекция Нематодозы. Аскаридоз
Нематодозы. Аскаридоз Аномалии количества зубов
Аномалии количества зубов Операции на органах шеи
Операции на органах шеи Методы диагностики гортани
Методы диагностики гортани Легочная гипертензия
Легочная гипертензия Сестринский уход за пациентами с хронической сердечной недостаточностью
Сестринский уход за пациентами с хронической сердечной недостаточностью Лечение открытых переломов
Лечение открытых переломов Збудники шигельозів, холери
Збудники шигельозів, холери Роль гормонов в регуляции роста, развития и гомеостаза
Роль гормонов в регуляции роста, развития и гомеостаза Тыныс алу ағзаларының аурулары
Тыныс алу ағзаларының аурулары Тері физиологиясы
Тері физиологиясы Бустеры Expert Boost. Уход за кожей
Бустеры Expert Boost. Уход за кожей Таным туралы ілім. Медициналық, ғылыми танымның ерекшелігі
Таным туралы ілім. Медициналық, ғылыми танымның ерекшелігі Placental abruption
Placental abruption Альбинизм. Типы альбинизма
Альбинизм. Типы альбинизма Современные аспекты сердечно-легочной и церебральной реанимации в стоматологической практике
Современные аспекты сердечно-легочной и церебральной реанимации в стоматологической практике Факторы риска при инфакте миокарда в развитии низкой физической активности найти информацию научных статей
Факторы риска при инфакте миокарда в развитии низкой физической активности найти информацию научных статей Балалардағы бұлшық ет жүйесінің анатомо-физиологиялық ерекшеліктері
Балалардағы бұлшық ет жүйесінің анатомо-физиологиялық ерекшеліктері Профилактическая медицина
Профилактическая медицина Аккредитация медицинских работников январь 2023
Аккредитация медицинских работников январь 2023