- Факторы патогенности бактерий

Содержание
- 2. Факторы патогенности Факторы патогенности – это молекулы или структуры микроорганизмов, наличие которых обеспечивает протекание инфекционного процесса.
- 3. Классификация Общепринятой классификации факторов патогенности бактерий не существует. Можно условно выделить следующие группы: Факторы адгезии Факторы
- 4. Часть I: Факторы адгезии
- 5. Бактериальная адгезия В большинстве случаев первым этапом взаимодействия бактериальных возбудителей с организмом человека является адгезия к
- 6. Варианты адгезинов Необходимость связывания с определенными молекулами определяет видовой тропизм и тканевый тропизм бактерий. Фимбриальные адгезины
- 7. Пертактин, филаментозный гемагглютинин (ФГА), и фимбрии на поверхности клеток Bordetella ФГА - экзопротеин (синий), секретируется через
- 8. Пили IV типа Пили IV типа (на примере Pil-белков представителей рода Neisseria) Пили IV типа способны
- 9. Адгезия как механизм инвазии Связывание бактерий с некоторыми поверхностными белками (интегрины, E-кадгерин) ведет к реорганизации цитоскелета
- 10. Часть II: Факторы инвазии, агрессии и добычи питательных веществ
- 11. Факторы инвазии, агрессии и добычи питательных веществ Данная группа факторов преимущественно представлена набором ферментов, которые расщепляют
- 12. Ферменты инвазии Коллагеназа – расщепление коллагена – основного белкового компонента соединительной ткани. Гиалуронидаза – расщепление гиалуроновой
- 13. Ферменты инвазии Фосфолипазы – расщепление фосфолипидов мембран, приводящее к гибели клеток. Данные ферменты рассматриваются не только
- 14. Ферменты инвазии Уреаза – гидролизует мочевину до аммиака и углекислого газа. Это приводит к: возможности использования
- 15. Железо как дефицитный ресурс Неотъемлемым компонентом белков дыхательной цепи и многих ферментов являются ионы железа Свободные
- 16. Системы перехвата железа Cидерофоры (пиовердин Pseudomonas aeruginosa) Рецепторы к железосодержащим белкам человека (трансферрин-связывающий белок Neisseria meningitidis)
- 17. Часть III: Антифагоцитарные факторы
- 18. Фагоцитоз Ключевую роль в инициации фагоцитоза играет распознавание – образование комплекса бактериальная структура – опсонин –
- 19. Капсулы Капсулы – наиболее распространенный защитный фактор бактерий. Для ряда патогенов (например, пневмококков) потеря способности синтезировать
- 20. Химическое строение капсул Как правило, капсулы состоят из полисахаридов или, реже, полипептидов. У многих бактерий капсулы
- 21. Слизистые чехлы Некоторые бактерии (например, Pseudomonas aeruginosa) образуют не плотные капсулы, а легко отделяющиеся от клетки
- 22. Укрытие белками организма Плазмокоагулаза Staphylococcus aureus – переводит фибриноген плазмы крови в нерастворимый фибрин, образуя подобие
- 23. Многие бактерии способны оценивать собственную численность в месте колонизации (феномен «чувства кворума», или «quorum sensing») и
- 24. Биоплёнки Биопленки представляют собой сплошные слои из бактериальных клеток и скрепляющего из каркаса из полисахаридов, белков
- 25. Биоплёнки как фактор патогенности Защита от фагоцитоза Устойчивость к антисептикам и дезинфектантам Устойчивость к антибиотикам Высокая
- 26. Клеточная стенка кислотоустойчивых бактерий Димиколат трегалозы (также известный как корд-фактор) является основным фактором патогенности кислотоустойчивых бактерий
- 27. Корд-фактор Толстый гидрофобный слой, создаваемый корд-фактором, обеспечивает защиту от многих антибактериальных веществ. Корд-фактор способен встраиваться в
- 28. Часть IV: Токсины
- 29. Бактериальные токсины Токсины – это соединения, синтезируемые бактериями и способные вызывать патологические изменения в организме. Бактериальные
- 30. Структура эндотоксина О-антиген Ядро (кор) Липид А Присутствует почти у всех грамотрицательных бактерий, за исключением спирохет
- 31. Механизм действия эндотоксина Эндотоксин не является токсином в эволюционном смысле. Его действие обусловлено связыванием липида А
- 32. Экзотоксины Действующие снаружи эукариотической клетки Действующие внутри эукариотической клетки Токсины, проникающие внутрь клетки, как правило, имеют
- 33. Экзотоксины Действующие снаружи эукариотической клетки Действующие внутри эукариотической клетки A B A B B B B
- 34. Виды экзотоксинов Большинство бактериальных токсинов можно отнести к одной из нескольких групп: Мембранотоксины Цитотоксины Нейротоксины Токсины,
- 35. Мембранотоксины Мембранотоксины – это белки, нарушающие целостность мембран эукариотических клеток, что приводит к их гибели. Это
- 36. Мембранотоксины Мембранотоксины можно разделить на группы: Порообразующие токсины – встраиваются в мембрану клетки и олигомеризуются, образуя
- 37. Мембранотоксины Мембранотоксины можно разделить на группы: Липазы – ферментативно расщепляют фосфолипиды и сфинголипиды мембран, приводя к
- 38. Цитотоксины Цитотоксины – это токсины, подавляющие жизненно важные процессы в клетках эукариот. Они приводят к нарушению
- 39. EFII EFII Цитотоксины Дифтерийный токсин вырабатывается штаммами Corynebacterium diphtheriae, содержащими tox+ профаг. Данный токсин является АДФ-рибозилтрансферазой.
- 40. Цитотоксины Шига-токсин вырабатывается Shigella dysenteriae, Шига-подобные токсины вырабатываются энтерогеморрагическими Escherichia coli. Данная группа токсинов является N-гликозидазами:
- 41. Нейротоксины Группа бактериальных белковых нейротоксинов включает в себя всего два представителя: Столбнячный токсин Clostridium tetani; Ботулинический
- 42. Нейротоксины Пресинаптическая мембрана Везикула Нейротоксины клостридий являются протеазами, специфически расщепляющими белки SNARE-комплекса, что приводит к невозможности
- 43. Нейротоксины Столбнячный токсин (тетаноспазмин) нарушает высвобождение тормозных неромедиаторов (ГАМК и глицина) из вставочных нейронов спинного мозга.
- 44. Нейротоксины Столбнячный токсин (тетаноспазмин) нарушает высвобождение тормозных неромедиаторов (ГАМК и глицина) из вставочных нейронов спинного мозга.
- 45. Токсины, нарушающие внутриклеточную передачу сигнала Действуют на уровне вторичных мессенджеров. Рецептор Gαs-белок АТФ цАМФ Аденилатциклаза Рецептор
- 46. Токсины, нарушающие внутриклеточную передачу сигнала АДФ-рибозилирование Gas белка, блокирующее его в «активном» состоянии Повышение уровня цАМФ
- 47. Токсин-опосредованная вирулентность Bordetella spp Комбинированное воздействие PT и ACT интоксикации и порообразования проявляется в ингибировании комплемент-зависимого
- 48. Токсины, нарушающие внутриклеточную передачу сигнала Рецептор факторов роста ГДФ ГТФ Ras Grb2 Sos MAPKKK MAPKK MAPK
- 49. Токсины, нарушающие внутриклеточную передачу сигнала Токсины α и β Clostridium difficile – являются гликозилтрансферазами. Они гликозилируют
- 50. Суперантигены Активация Т-лимфоцитов не может происходить при простом контакте рецептора с антигеном. Короткий фрагмент антигена должен
- 51. Суперантигены Суперантигены – это токсины бактерий, которые перекрестно связывают TCR и MHC II класса независимо от
- 52. Часть V: Инъецируемые эффекторные белки
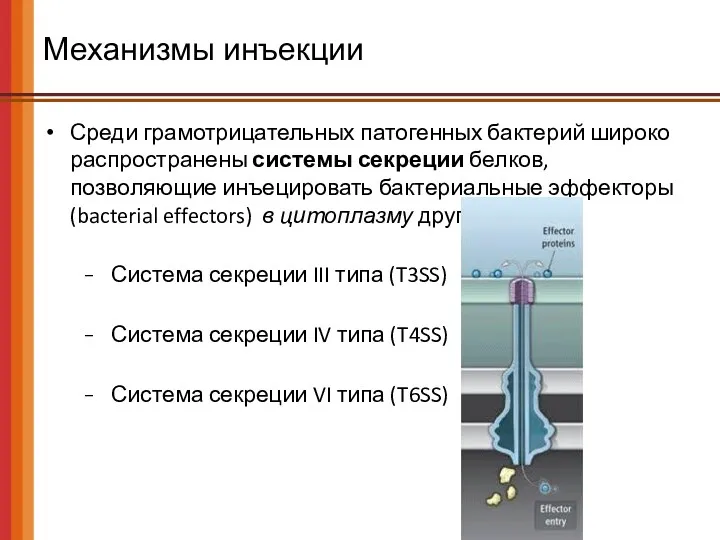
- 53. Механизмы инъекции Среди грамотрицательных патогенных бактерий широко распространены системы секреции белков, позволяющие инъецировать бактериальные эффекторы (bacterial
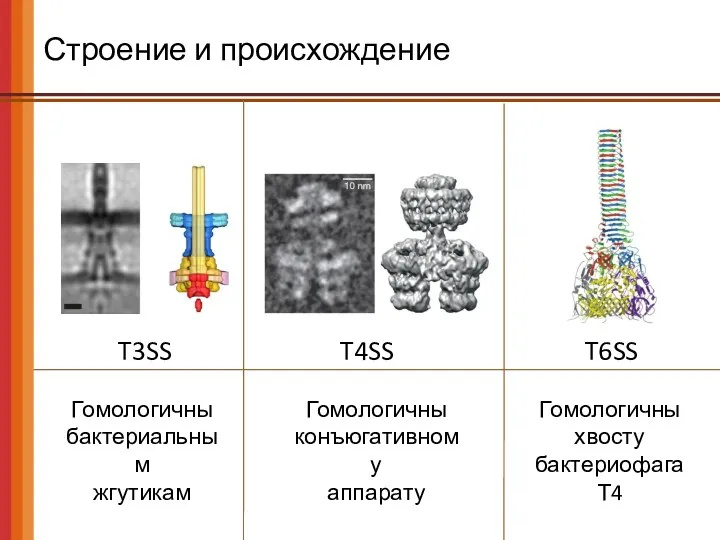
- 54. Строение и происхождение T3SS T4SS T6SS Гомологичны бактериальным жгутикам Гомологичны конъюгативному аппарату Гомологичны хвосту бактериофага Т4
- 55. Схематическое строение бактериофага Т4 и предполагаемой структуры впрыска у бактерий - системы секреции VI типа (T6SS).
- 56. Бактериальные эффекторы Бактерии чаще всего невыгодно уничтожать клетку, на которой она адгезирована. Бактериальные эффекторы обычно действуют
- 57. Распространенные эффекты Самые распространенные действия бактериальных эффекторов: Захват бактерий нефагоцитирующими клетками с использованием “trigger mechanism”. Ингибирование
- 58. Участие в адгезии: энтеропатогенные Escherichia coli Энтеропатогенные E. coli способны образовывать «пьедесталы» на поверхности кишечного эпителия,
- 59. Участие в инвазии: энтероинвазивные Escherichia coli и Shigella spp. Вирулентность данных бактерий в первую очередь обеспечивается
- 60. Участие в процессе выживания в макрофагах: Legionella pneumophila Данные бактерии способны синтезировать около 300 эффекторов, доставляемых
- 62. Скачать презентацию

Факторы патогенности
Факторы патогенности – это молекулы или структуры микроорганизмов, наличие которых
Факторы патогенности
Факторы патогенности – это молекулы или структуры микроорганизмов, наличие которых

Классификация
Общепринятой классификации факторов патогенности бактерий не существует.
Можно условно выделить следующие
Классификация
Общепринятой классификации факторов патогенности бактерий не существует.
Можно условно выделить следующие
Часть I:
Факторы адгезии
Часть I:
Факторы адгезии

Бактериальная адгезия
В большинстве случаев первым этапом взаимодействия бактериальных возбудителей с организмом
Бактериальная адгезия
В большинстве случаев первым этапом взаимодействия бактериальных возбудителей с организмом
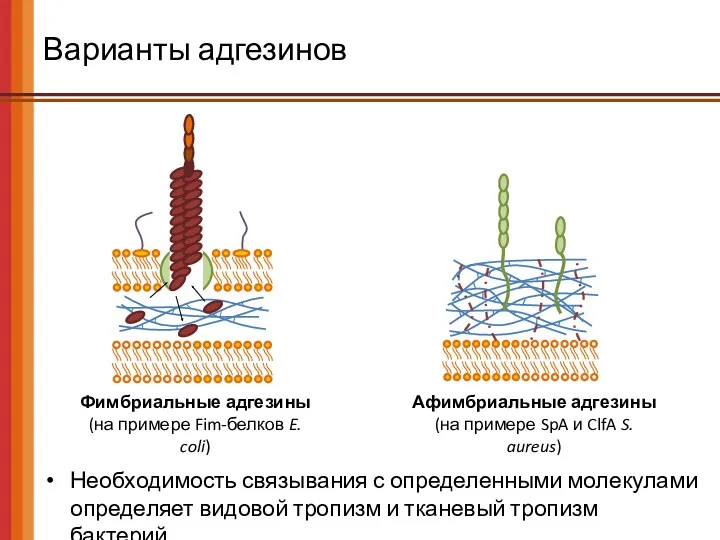
Варианты адгезинов
Необходимость связывания с определенными молекулами определяет видовой тропизм и тканевый
Варианты адгезинов
Необходимость связывания с определенными молекулами определяет видовой тропизм и тканевый
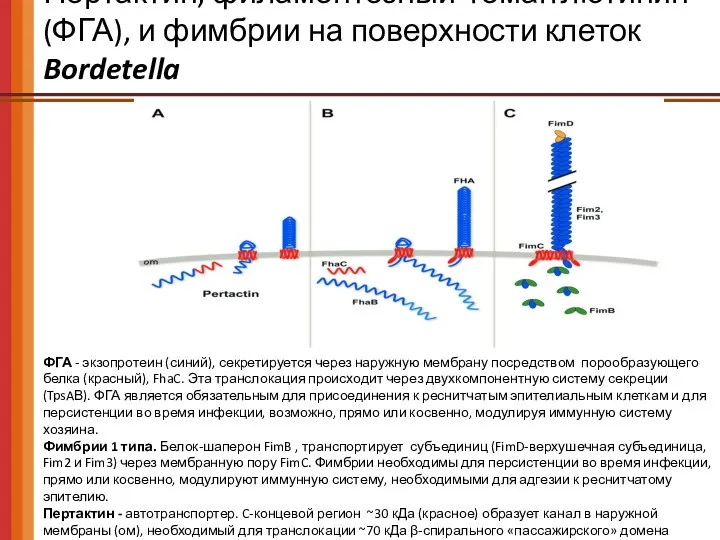
Пертактин, филаментозный гемагглютинин (ФГА), и фимбрии на поверхности клеток Bordetella
ФГА -
Пертактин, филаментозный гемагглютинин (ФГА), и фимбрии на поверхности клеток Bordetella
ФГА -
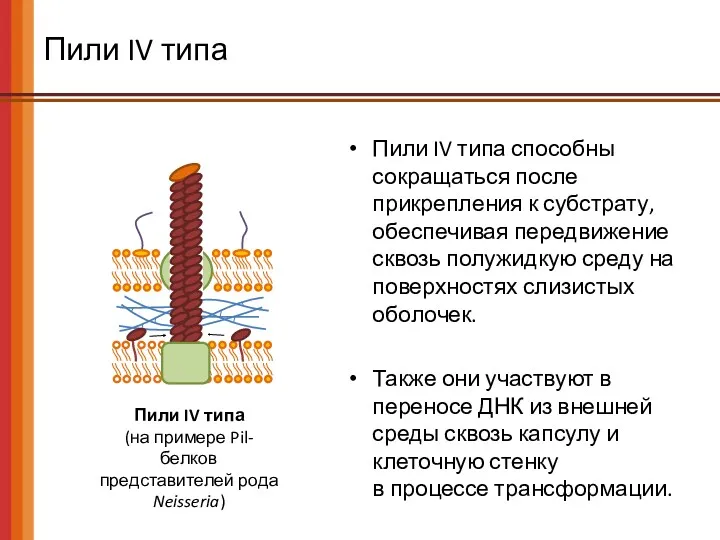
Пили IV типа
Пили IV типа
(на примере Pil-белков
представителей рода
Neisseria)
Пили IV
Пили IV типа
Пили IV типа
(на примере Pil-белков
представителей рода
Neisseria)
Пили IV
Адгезия как механизм инвазии
Связывание бактерий с некоторыми поверхностными белками (интегрины, E-кадгерин)
Адгезия как механизм инвазии
Связывание бактерий с некоторыми поверхностными белками (интегрины, E-кадгерин)

Часть II:
Факторы инвазии, агрессии и добычи питательных веществ
Часть II:
Факторы инвазии, агрессии и добычи питательных веществ

Факторы инвазии, агрессии и добычи питательных веществ
Данная группа факторов преимущественно представлена
Факторы инвазии, агрессии и добычи питательных веществ
Данная группа факторов преимущественно представлена

Ферменты инвазии
Коллагеназа – расщепление коллагена – основного белкового компонента соединительной ткани.
Ферменты инвазии
Коллагеназа – расщепление коллагена – основного белкового компонента соединительной ткани.

Ферменты инвазии
Фосфолипазы – расщепление фосфолипидов мембран, приводящее к гибели клеток. Данные
Ферменты инвазии
Фосфолипазы – расщепление фосфолипидов мембран, приводящее к гибели клеток. Данные

Ферменты инвазии
Уреаза – гидролизует мочевину до аммиака и углекислого газа.
Это приводит
Ферменты инвазии
Уреаза – гидролизует мочевину до аммиака и углекислого газа.
Это приводит
Железо как дефицитный ресурс
Неотъемлемым компонентом белков дыхательной цепи и многих ферментов
Железо как дефицитный ресурс
Неотъемлемым компонентом белков дыхательной цепи и многих ферментов
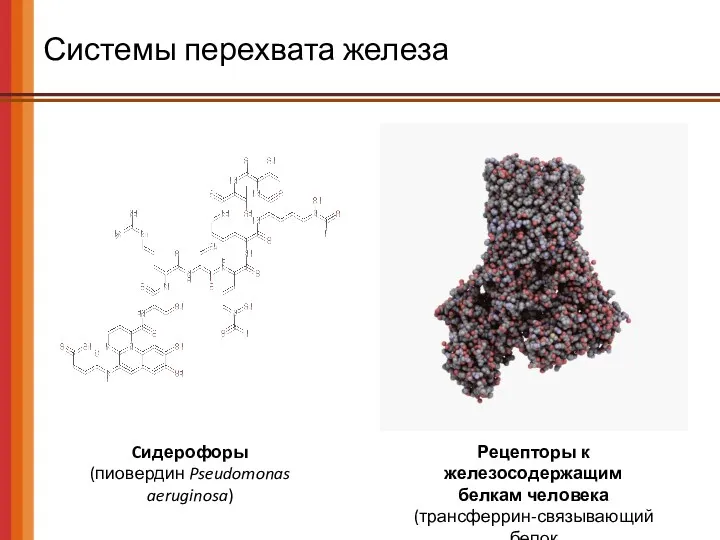
Системы перехвата железа
Cидерофоры
(пиовердин Pseudomonas aeruginosa)
Рецепторы к железосодержащим
белкам человека
(трансферрин-связывающий белок
Neisseria
Системы перехвата железа
Cидерофоры
(пиовердин Pseudomonas aeruginosa)
Рецепторы к железосодержащим белкам человека (трансферрин-связывающий белок Neisseria
Часть III:
Антифагоцитарные факторы
Часть III:
Антифагоцитарные факторы
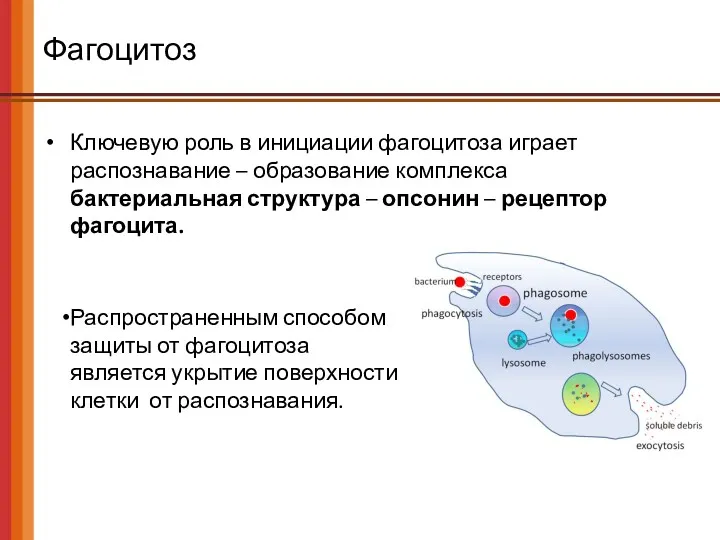
Фагоцитоз
Ключевую роль в инициации фагоцитоза играет распознавание – образование комплекса
бактериальная
Фагоцитоз
Ключевую роль в инициации фагоцитоза играет распознавание – образование комплекса бактериальная

Капсулы
Капсулы – наиболее распространенный защитный фактор бактерий. Для ряда патогенов (например,
Капсулы
Капсулы – наиболее распространенный защитный фактор бактерий. Для ряда патогенов (например,

Химическое строение капсул
Как правило, капсулы состоят из полисахаридов или, реже, полипептидов.
У
Химическое строение капсул
Как правило, капсулы состоят из полисахаридов или, реже, полипептидов.
У
Слизистые чехлы
Некоторые бактерии (например, Pseudomonas aeruginosa) образуют не плотные капсулы, а
Слизистые чехлы
Некоторые бактерии (например, Pseudomonas aeruginosa) образуют не плотные капсулы, а

Укрытие белками организма
Плазмокоагулаза Staphylococcus aureus – переводит фибриноген плазмы крови в
Укрытие белками организма
Плазмокоагулаза Staphylococcus aureus – переводит фибриноген плазмы крови в
Многие бактерии способны оценивать собственную численность в месте колонизации (феномен «чувства
Многие бактерии способны оценивать собственную численность в месте колонизации (феномен «чувства

Биоплёнки
Биопленки представляют собой сплошные слои из бактериальных клеток и скрепляющего из
Биоплёнки
Биопленки представляют собой сплошные слои из бактериальных клеток и скрепляющего из

Биоплёнки как фактор патогенности
Защита от фагоцитоза
Устойчивость к антисептикам и дезинфектантам
Устойчивость к
Биоплёнки как фактор патогенности
Защита от фагоцитоза
Устойчивость к антисептикам и дезинфектантам
Устойчивость к
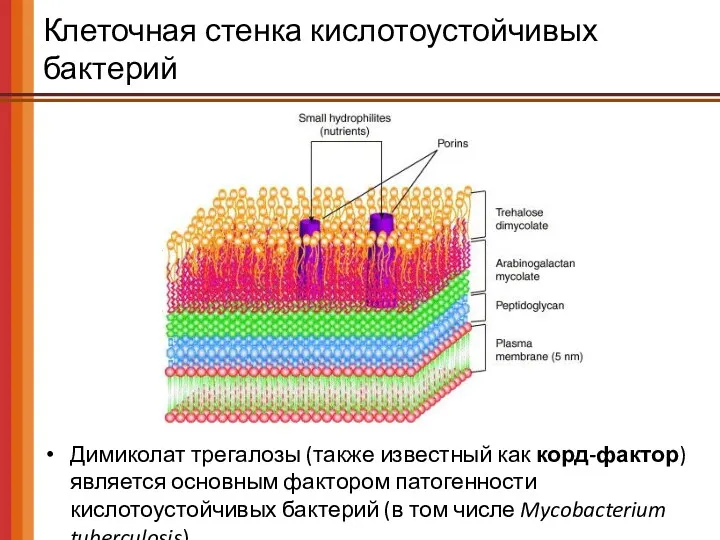
Клеточная стенка кислотоустойчивых бактерий
Димиколат трегалозы (также известный как корд-фактор) является основным
Клеточная стенка кислотоустойчивых бактерий
Димиколат трегалозы (также известный как корд-фактор) является основным

Корд-фактор
Толстый гидрофобный слой, создаваемый корд-фактором, обеспечивает защиту от многих антибактериальных веществ.
Корд-фактор
Корд-фактор
Толстый гидрофобный слой, создаваемый корд-фактором, обеспечивает защиту от многих антибактериальных веществ.
Корд-фактор

Часть IV:
Токсины
Часть IV:
Токсины
Бактериальные токсины
Токсины – это соединения, синтезируемые бактериями и способные вызывать патологические
Бактериальные токсины
Токсины – это соединения, синтезируемые бактериями и способные вызывать патологические
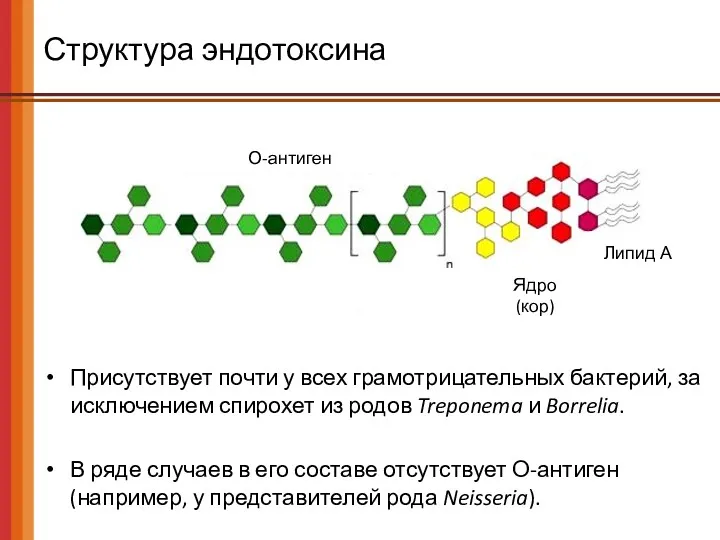
Структура эндотоксина
О-антиген
Ядро
(кор)
Липид А
Присутствует почти у всех грамотрицательных бактерий, за исключением спирохет
Структура эндотоксина
О-антиген
Ядро
(кор)
Липид А
Присутствует почти у всех грамотрицательных бактерий, за исключением спирохет

Механизм действия эндотоксина
Эндотоксин не является токсином в эволюционном смысле.
Его действие обусловлено
Механизм действия эндотоксина
Эндотоксин не является токсином в эволюционном смысле.
Его действие обусловлено

Экзотоксины
Действующие снаружи эукариотической клетки
Действующие внутри эукариотической клетки
Токсины, проникающие внутрь клетки, как
Экзотоксины
Действующие снаружи эукариотической клетки
Действующие внутри эукариотической клетки
Токсины, проникающие внутрь клетки, как
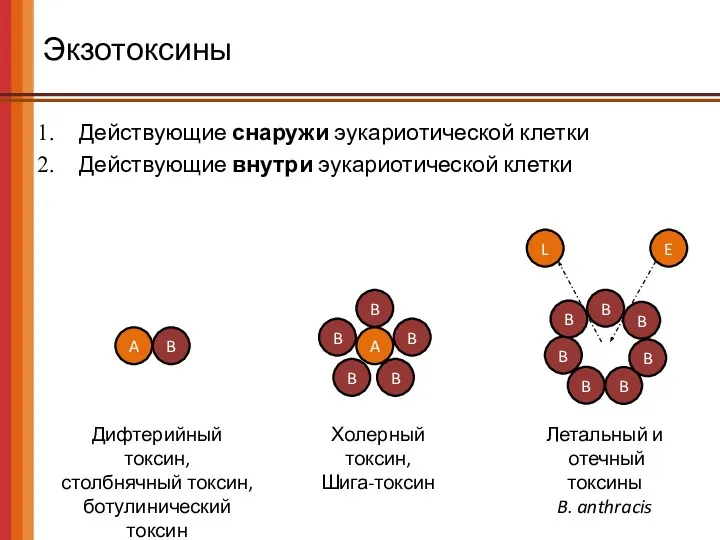
Экзотоксины
Действующие снаружи эукариотической клетки
Действующие внутри эукариотической клетки
A
B
A
B
B
B
B
B
Дифтерийный токсин,
столбнячный токсин,
ботулинический токсин
Холерный токсин,
Шига-токсин
Летальный
Экзотоксины
Действующие снаружи эукариотической клетки
Действующие внутри эукариотической клетки
A
B
A
B
B
B
B
B
Дифтерийный токсин,
столбнячный токсин,
ботулинический токсин
Холерный токсин,
Шига-токсин
Летальный

Виды экзотоксинов
Большинство бактериальных токсинов можно отнести к одной из нескольких групп:
Мембранотоксины
Цитотоксины
Нейротоксины
Токсины,
Виды экзотоксинов
Большинство бактериальных токсинов можно отнести к одной из нескольких групп:
Мембранотоксины
Цитотоксины
Нейротоксины
Токсины,

Мембранотоксины
Мембранотоксины – это белки, нарушающие целостность мембран эукариотических клеток, что приводит
Мембранотоксины
Мембранотоксины – это белки, нарушающие целостность мембран эукариотических клеток, что приводит
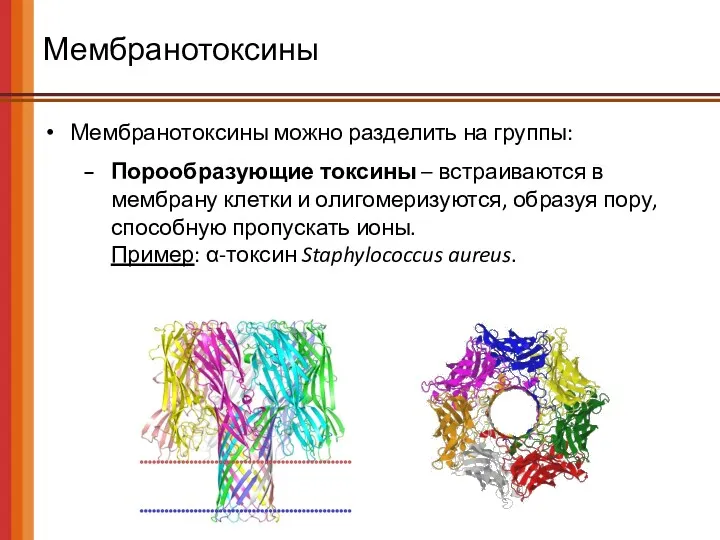
Мембранотоксины
Мембранотоксины можно разделить на группы:
Порообразующие токсины – встраиваются в мембрану клетки
Мембранотоксины
Мембранотоксины можно разделить на группы:
Порообразующие токсины – встраиваются в мембрану клетки
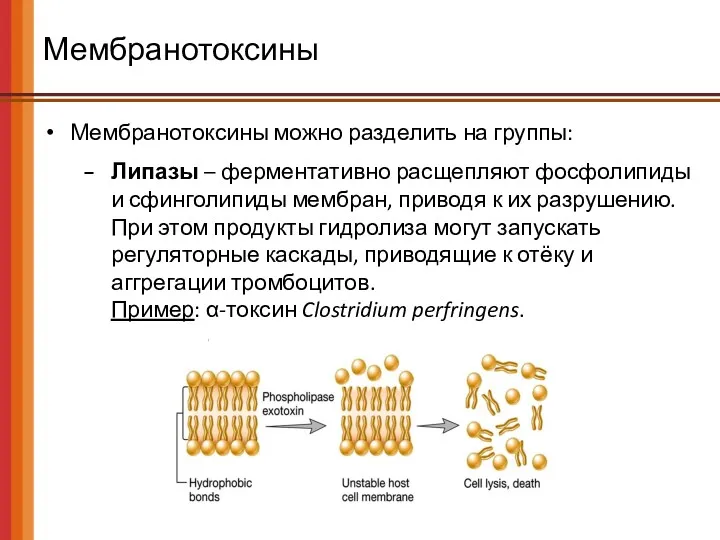
Мембранотоксины
Мембранотоксины можно разделить на группы:
Липазы – ферментативно расщепляют фосфолипиды и сфинголипиды
Мембранотоксины
Мембранотоксины можно разделить на группы:
Липазы – ферментативно расщепляют фосфолипиды и сфинголипиды

Цитотоксины
Цитотоксины – это токсины, подавляющие жизненно важные процессы в клетках эукариот.
Цитотоксины
Цитотоксины – это токсины, подавляющие жизненно важные процессы в клетках эукариот.
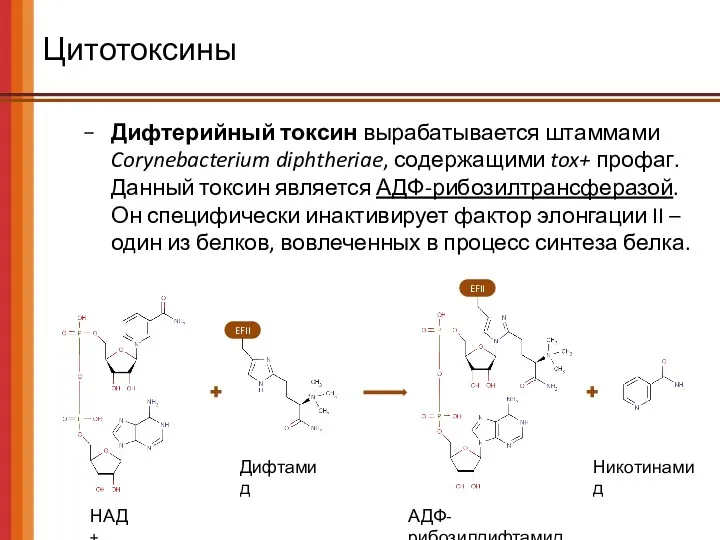
EFII
EFII
Цитотоксины
Дифтерийный токсин вырабатывается штаммами Corynebacterium diphtheriae, содержащими tox+ профаг.
Данный токсин
EFII
EFII
Цитотоксины
Дифтерийный токсин вырабатывается штаммами Corynebacterium diphtheriae, содержащими tox+ профаг. Данный токсин

Цитотоксины
Шига-токсин вырабатывается
Shigella dysenteriae,
Шига-подобные токсины
вырабатываются
энтерогеморрагическими
Escherichia coli.
Данная группа токсинов
Цитотоксины
Шига-токсин вырабатывается Shigella dysenteriae, Шига-подобные токсины вырабатываются энтерогеморрагическими Escherichia coli. Данная группа токсинов

Нейротоксины
Группа бактериальных белковых нейротоксинов включает в себя всего два представителя:
Столбнячный токсин
Нейротоксины
Группа бактериальных белковых нейротоксинов включает в себя всего два представителя:
Столбнячный токсин
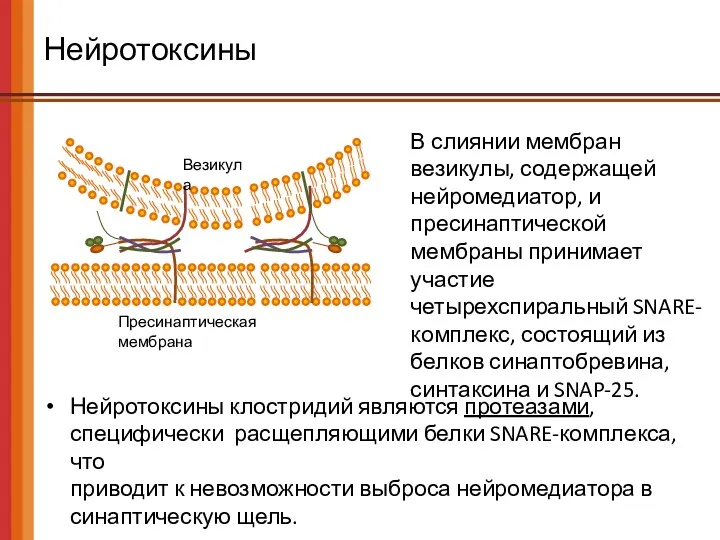
Нейротоксины
Пресинаптическая мембрана
Везикула
Нейротоксины клостридий являются протеазами, специфически расщепляющими белки SNARE-комплекса, что
приводит
Нейротоксины
Пресинаптическая мембрана
Везикула
Нейротоксины клостридий являются протеазами, специфически расщепляющими белки SNARE-комплекса, что приводит
Нейротоксины
Столбнячный токсин (тетаноспазмин) нарушает высвобождение тормозных неромедиаторов (ГАМК и глицина) из
Нейротоксины
Столбнячный токсин (тетаноспазмин) нарушает высвобождение тормозных неромедиаторов (ГАМК и глицина) из
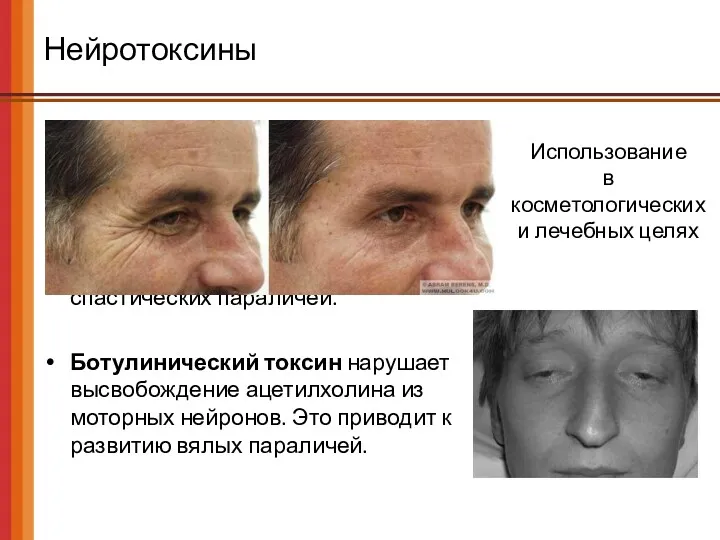
Нейротоксины
Столбнячный токсин (тетаноспазмин) нарушает высвобождение тормозных неромедиаторов (ГАМК и глицина) из
Нейротоксины
Столбнячный токсин (тетаноспазмин) нарушает высвобождение тормозных неромедиаторов (ГАМК и глицина) из
Токсины, нарушающие внутриклеточную передачу сигнала
Действуют на уровне вторичных мессенджеров.
Рецептор
Gαs-белок
АТФ
цАМФ
Аденилатциклаза
Рецептор
Gαi-белок
ГДФ
ГТФ
ГДФ
ГТФ
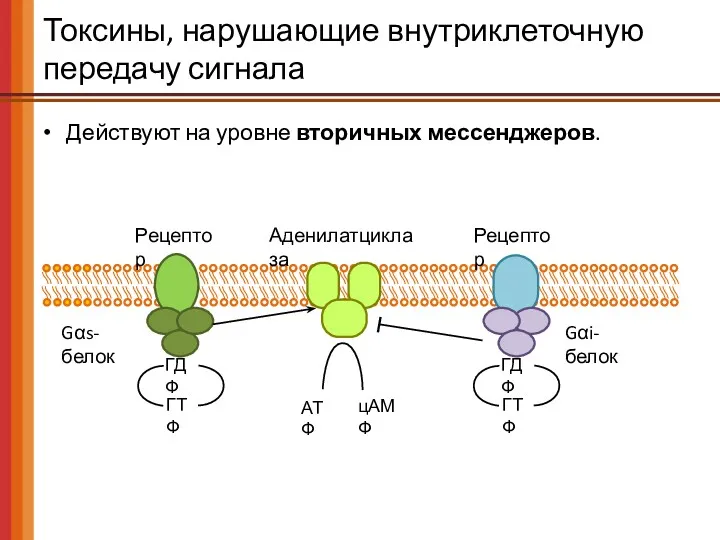
Токсины, нарушающие внутриклеточную передачу сигнала
Действуют на уровне вторичных мессенджеров.
Рецептор
Gαs-белок
АТФ
цАМФ
Аденилатциклаза
Рецептор
Gαi-белок
ГДФ
ГТФ
ГДФ
ГТФ
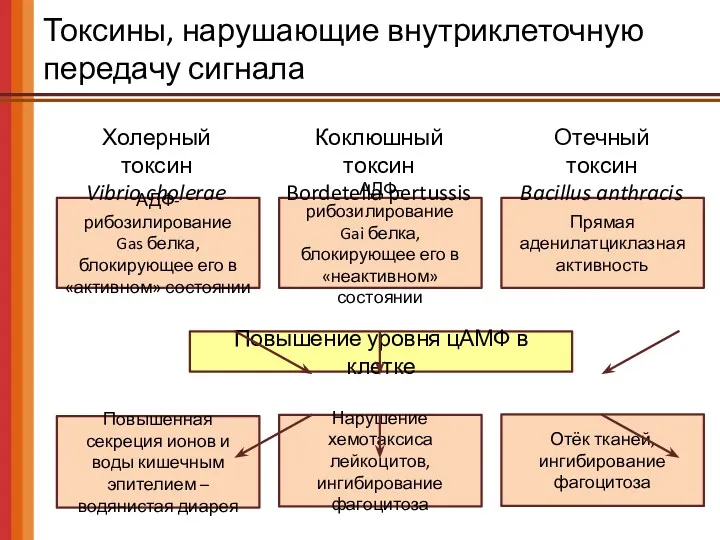
Токсины, нарушающие внутриклеточную передачу сигнала
АДФ-рибозилирование
Gas белка, блокирующее его в «активном» состоянии
Повышение
Токсины, нарушающие внутриклеточную передачу сигнала
АДФ-рибозилирование
Gas белка, блокирующее его в «активном» состоянии
Повышение
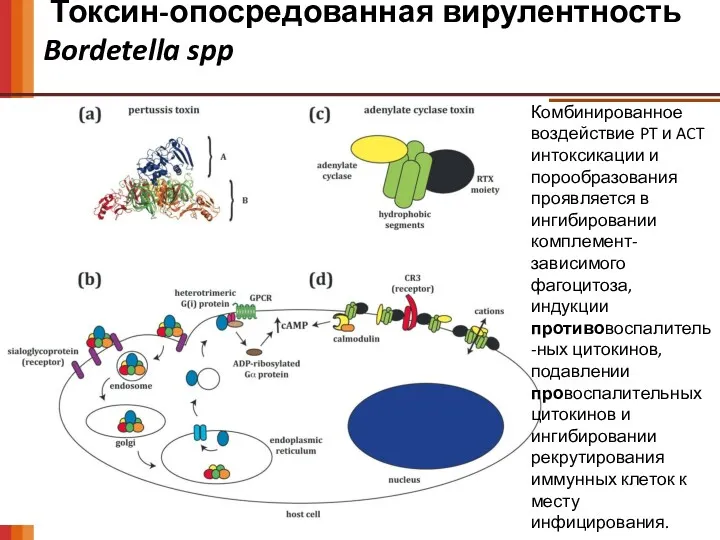
Токсин-опосредованная вирулентность
Bordetella spp
Комбинированное воздействие PT и ACT интоксикации и
Токсин-опосредованная вирулентность
Bordetella spp
Комбинированное воздействие PT и ACT интоксикации и
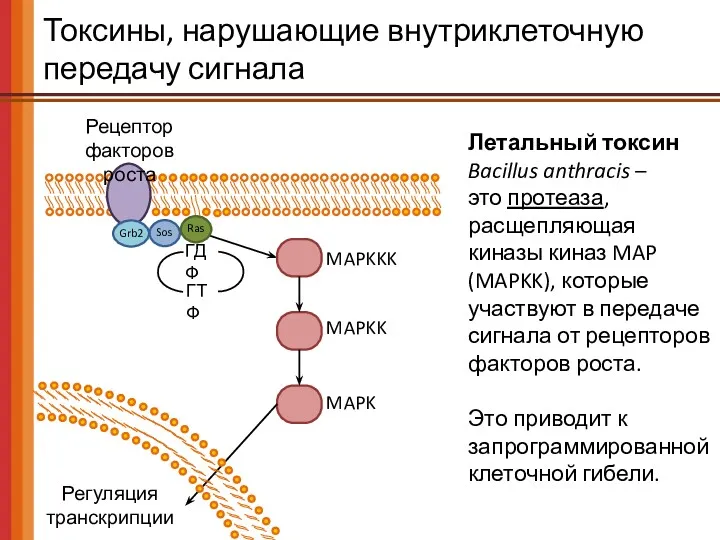
Токсины, нарушающие внутриклеточную передачу сигнала
Рецептор
факторов роста
ГДФ
ГТФ
Ras
Grb2
Sos
MAPKKK
MAPKK
MAPK
Регуляция
транскрипции
Летальный токсин
Bacillus anthracis –
Токсины, нарушающие внутриклеточную передачу сигнала
Рецептор
факторов роста
ГДФ
ГТФ
Ras
Grb2
Sos
MAPKKK
MAPKK
MAPK
Регуляция
транскрипции
Летальный токсин Bacillus anthracis –
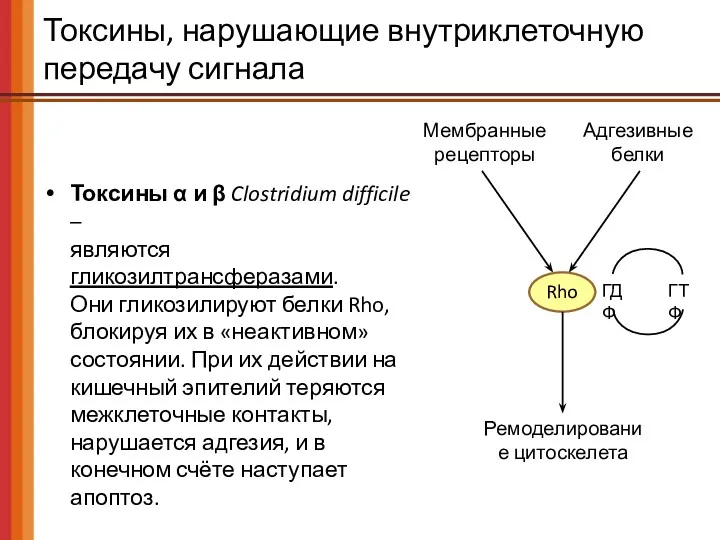
Токсины, нарушающие внутриклеточную передачу сигнала
Токсины α и β Clostridium difficile –
Токсины, нарушающие внутриклеточную передачу сигнала
Токсины α и β Clostridium difficile –
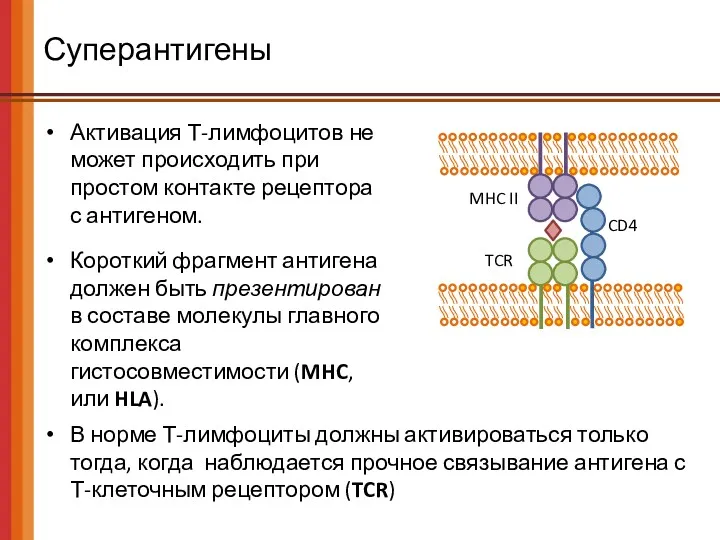
Суперантигены
Активация Т-лимфоцитов не может происходить при простом контакте рецептора с антигеном.
Суперантигены
Активация Т-лимфоцитов не может происходить при простом контакте рецептора с антигеном.
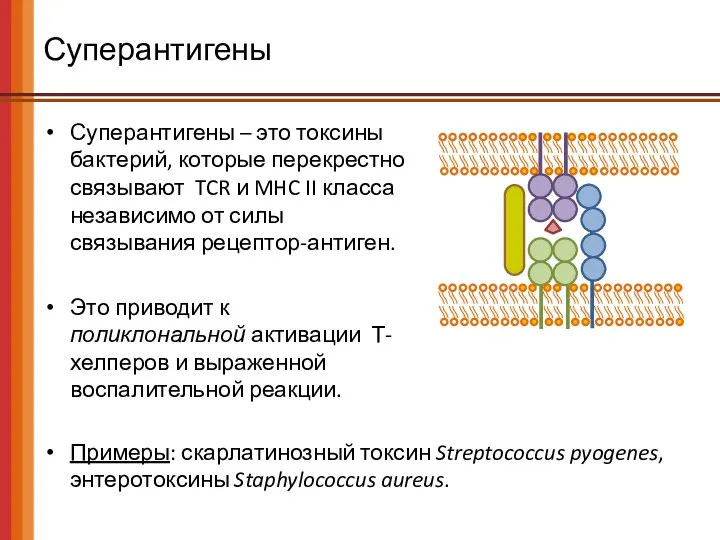
Суперантигены
Суперантигены – это токсины бактерий, которые перекрестно связывают TCR и MHC
Суперантигены
Суперантигены – это токсины бактерий, которые перекрестно связывают TCR и MHC

Часть V:
Инъецируемые эффекторные белки
Часть V:
Инъецируемые эффекторные белки
Механизмы инъекции
Среди грамотрицательных патогенных бактерий широко распространены системы секреции белков, позволяющие
Механизмы инъекции
Среди грамотрицательных патогенных бактерий широко распространены системы секреции белков, позволяющие
Строение и происхождение
T3SS
T4SS
T6SS
Гомологичны
бактериальным
жгутикам
Гомологичны
конъюгативному
аппарату
Гомологичны
хвосту бактериофага Т4
Строение и происхождение
T3SS
T4SS
T6SS
Гомологичны
бактериальным
жгутикам
Гомологичны
конъюгативному
аппарату
Гомологичны
хвосту бактериофага Т4
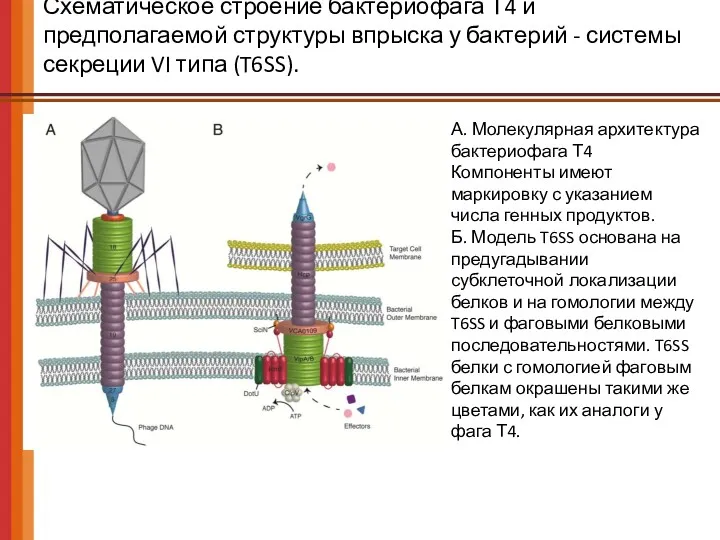
Схематическое строение бактериофага Т4 и предполагаемой структуры впрыска у бактерий -
Схематическое строение бактериофага Т4 и предполагаемой структуры впрыска у бактерий -

Бактериальные эффекторы
Бактерии чаще всего невыгодно уничтожать клетку, на которой она адгезирована.
Бактериальные
Бактериальные эффекторы
Бактерии чаще всего невыгодно уничтожать клетку, на которой она адгезирована.
Бактериальные

Распространенные эффекты
Самые распространенные действия
бактериальных эффекторов:
Захват бактерий
нефагоцитирующими
клетками с использованием
“trigger mechanism”.
Ингибирование
Распространенные эффекты
Самые распространенные действия
бактериальных эффекторов:
Захват бактерий
нефагоцитирующими
клетками с использованием
“trigger mechanism”.
Ингибирование
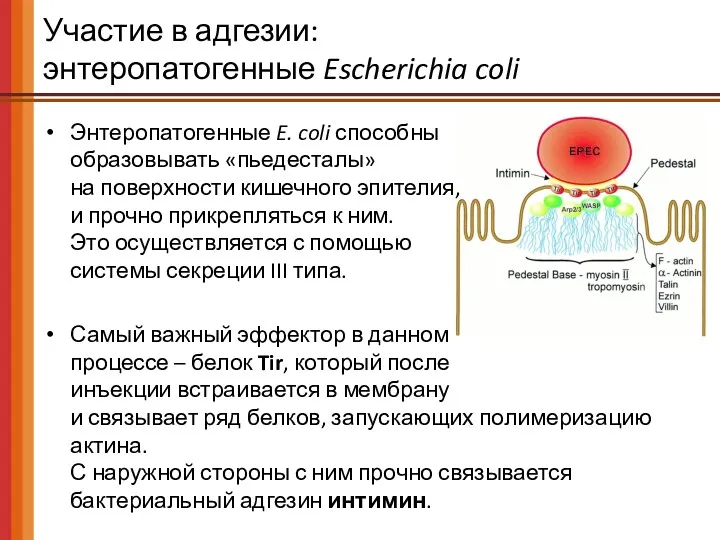
Участие в адгезии:
энтеропатогенные Escherichia coli
Энтеропатогенные E. coli способны
образовывать «пьедесталы»
на
Участие в адгезии:
энтеропатогенные Escherichia coli
Энтеропатогенные E. coli способны образовывать «пьедесталы» на
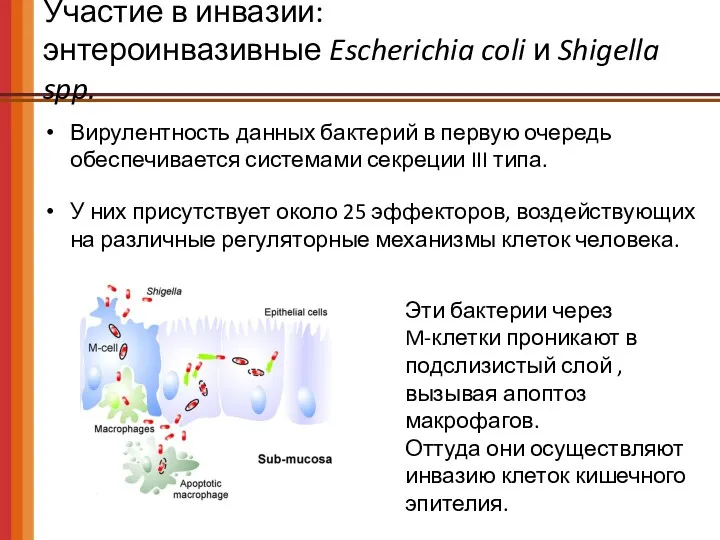
Участие в инвазии:
энтероинвазивные Escherichia coli и Shigella spp.
Вирулентность данных бактерий
Участие в инвазии:
энтероинвазивные Escherichia coli и Shigella spp.
Вирулентность данных бактерий

Участие в процессе выживания в макрофагах:
Legionella pneumophila
Данные бактерии способны
синтезировать около
Участие в процессе выживания в макрофагах:
Legionella pneumophila
Данные бактерии способны синтезировать около
 Заболевания ЛОР-органов
Заболевания ЛОР-органов Комбинированные оральные контрацептивы (КОК)
Комбинированные оральные контрацептивы (КОК) СПИД и ВИЧ. Риск и способы заражения
СПИД и ВИЧ. Риск и способы заражения Острые воспалительные заболевания матки и придатков как причина развития клиники острого живота в гинекологии
Острые воспалительные заболевания матки и придатков как причина развития клиники острого живота в гинекологии Гигиена детей и подростков
Гигиена детей и подростков Физиология сосудов
Физиология сосудов Рак шейки матки. Основные ошибки гинекологов с позиции онколога. Органосохраняющие операции
Рак шейки матки. Основные ошибки гинекологов с позиции онколога. Органосохраняющие операции Лучевая семиотика повреждений и заболеваний опорно-двигательного аппарата
Лучевая семиотика повреждений и заболеваний опорно-двигательного аппарата Повреждения предплечья и кисти
Повреждения предплечья и кисти Діагностичні труднощі ведення пацієнта з диспептичним синдромом
Діагностичні труднощі ведення пацієнта з диспептичним синдромом Сифилис. Общее течение сифилиса
Сифилис. Общее течение сифилиса Медико-генетическое консультирование
Медико-генетическое консультирование Жабық бассүйек-ми жарақаты
Жабық бассүйек-ми жарақаты Внутриутробные инфекции у детей
Внутриутробные инфекции у детей Позвоночник. Функции позвоночника
Позвоночник. Функции позвоночника Общая характеристика учетной политики аптечной организации
Общая характеристика учетной политики аптечной организации Планирование семьи. Контрацепция
Планирование семьи. Контрацепция АҚШҚ аурулары. АҚШҚ аурулары кезінде протездеу түрлері. АҚШҚ аурулары кезінде протездеу ерекшеліктері
АҚШҚ аурулары. АҚШҚ аурулары кезінде протездеу түрлері. АҚШҚ аурулары кезінде протездеу ерекшеліктері Повреждения мягких тканей, сухожилий и суставов. Вывихи. Классификация, клиника, диагностика
Повреждения мягких тканей, сухожилий и суставов. Вывихи. Классификация, клиника, диагностика Аэроб және анаэроб бактериалардың ажырату және таза дақылын бөліп алу сатылары
Аэроб және анаэроб бактериалардың ажырату және таза дақылын бөліп алу сатылары Техника и последовательность выполнения приемов плоскостного массажа
Техника и последовательность выполнения приемов плоскостного массажа Топографическая анатомия и оперативная хирургия печени, желчного пузыря
Топографическая анатомия и оперативная хирургия печени, желчного пузыря Роль TLRs в патогенезе преждевременных родов
Роль TLRs в патогенезе преждевременных родов Пиодермии. Стафилококки
Пиодермии. Стафилококки Антибиотики. Антибактериальные химиотерапевтические средства
Антибиотики. Антибактериальные химиотерапевтические средства Ларингит
Ларингит Оппортунистические заболевания при ВИЧ‐инфекции
Оппортунистические заболевания при ВИЧ‐инфекции Первая помощь при пищевом отравлении
Первая помощь при пищевом отравлении