- Ферменты. Свойства, строение, медико-биологическая значимость

Содержание
- 2. История вопроса о ферментах Термин фермент предложен в XVII веке химиком ван Гельмонтом при обсуждении механизмов
- 3. История вопроса о ферментах Через два года после смерти Л. Пастера в 1897 году Э. Бухнер
- 4. История вопроса о ферментах Каталитическая активность РНК впервые была обнаружена в 1980-е годы у пре-рРНК Томасом
- 5. Ферменты или энзимы (от лат. fermentum, греч. ζύμη, ἔνζυμον — закваска) — обычно белковые молекулы или
- 6. Наука о ферментах называется энзимологией, а не ферментологией (чтобы не смешивать корни слов латинского и греческого
- 7. Классификация ферментов По типу катализируемых реакций ферменты подразделяются на 6 классов согласно иерархической классификации ферментов (КФ,
- 8. КФ1: Оксидоредуктазы, катализируют перенос электронов, то есть окисление или восстановление. Систематические названия ферментов класса образуются по
- 9. КФ 2: Трансферазы Катализируют перенос химических групп с одной молекулы субстрата на другую. Среди трансфераз особо
- 10. КФ 3: Гидролазы Катализируют гидролиз химических связей. Это класс ферментов, катализирующий гидролиз ковалентной связи. Общий вид
- 11. КФ 4: Лиазы Отдельный класс ферментов, катализирующих реакции негидролитического и неокислительного разрыва различных химических связей (C—C,
- 12. КФ 5: Изомеразы Ферменты, катализирующие структурные превращения изомеров (рацемизация или эпимеризация). Изомеразы – ферменты, катализирующие изомерные
- 13. КФ 5: Изомеразы Если рассматривать все подклассы, то изомеразы делятся по типу изомеризации на: рацемазы и
- 14. КФ 6: Лигазы Ферменты, катализирующие соединение двух молекул с образованием новой химической связи (лигирование). При этом
- 15. Лигазы в генной инженерии Иными словами, ДНК-лигазы сшивают рядом расположенные нуклеотиды, образуя связь между остатками сахаров.
- 16. Информация о ферментах в сети Internet
- 17. Информация о ферментах в сети Internet
- 18. Информация о ферментах в сети Internet
- 19. Информация о ферментах в сети Internet
- 20. Лактатдегидрогеназа (ЛДГ) Лактатдегидрогеназа (ЛДГ; L-лактат-НАД-оксидоредуктаза, КФ 1.1.1.27) - цинксодержаший фермент, катализирует обратимую реакцию восстановления пировиноградной кислоты
- 21. Значимость в диагностике Клиническое значение определения активности ЛДГ Активность ЛДГ в сыворотке крови повышается при многих
- 22. .Креатинкиназа (КК) Креатинкиназа-АТФ: креатин-1,4-фосфотрансфераза (КФ 2.7.3.2) — катализирует обратимый перенос фосфатного остатка между АТФ и креатином
- 23. у-Глутамилтранспептидаза ГГТ; у-глутамил-транспептидазы, КФ 2.3.2.2). ГГТ катализирует перенос у-глутамила на аминокислоту или пептид, на другую молекулу
- 24. Значение в диагностике Наиболее частая причина повышения активности ГГТ в сыворотке крови — патология печени. Слабое
- 26. Скачать презентацию

История вопроса о ферментах
Термин фермент предложен в XVII веке химиком ван Гельмонтом при обсуждении механизмов пищеварения.
В
История вопроса о ферментах
Термин фермент предложен в XVII веке химиком ван Гельмонтом при обсуждении механизмов пищеварения.
В

История вопроса о ферментах
Через два года после смерти Л. Пастера в 1897
История вопроса о ферментах
Через два года после смерти Л. Пастера в 1897

История вопроса о ферментах
Каталитическая активность РНК впервые была обнаружена в 1980-е
История вопроса о ферментах
Каталитическая активность РНК впервые была обнаружена в 1980-е

Ферменты или энзимы
(от лат. fermentum, греч. ζύμη, ἔνζυμον — закваска) — обычно белковые молекулы или молекулы РНК (рибозимы) или их комплексы, ускоряющие (катализирующие) химические
Ферменты или энзимы
(от лат. fermentum, греч. ζύμη, ἔνζυμον — закваска) — обычно белковые молекулы или молекулы РНК (рибозимы) или их комплексы, ускоряющие (катализирующие) химические

Наука о ферментах называется энзимологией, а не ферментологией (чтобы не смешивать корни
Наука о ферментах называется энзимологией, а не ферментологией (чтобы не смешивать корни

Классификация ферментов
По типу катализируемых реакций ферменты подразделяются на 6 классов согласно
Классификация ферментов
По типу катализируемых реакций ферменты подразделяются на 6 классов согласно

КФ1: Оксидоредуктазы, катализируют перенос электронов, то есть окисление или восстановление.
Систематические
КФ1: Оксидоредуктазы, катализируют перенос электронов, то есть окисление или восстановление.
Систематические

КФ 2: Трансферазы
Катализируют перенос химических групп с одной молекулы субстрата на другую.
Среди трансфераз
КФ 2: Трансферазы
Катализируют перенос химических групп с одной молекулы субстрата на другую.
Среди трансфераз

КФ 3: Гидролазы
Катализируют гидролиз химических связей.
Это класс ферментов, катализирующий гидролиз ковалентной связи. Общий вид реакции, катализируемой гидролазой
КФ 3: Гидролазы
Катализируют гидролиз химических связей.
Это класс ферментов, катализирующий гидролиз ковалентной связи. Общий вид реакции, катализируемой гидролазой

КФ 4: Лиазы
Отдельный класс ферментов, катализирующих реакции негидролитического и неокислительного разрыва различных химических
КФ 4: Лиазы
Отдельный класс ферментов, катализирующих реакции негидролитического и неокислительного разрыва различных химических
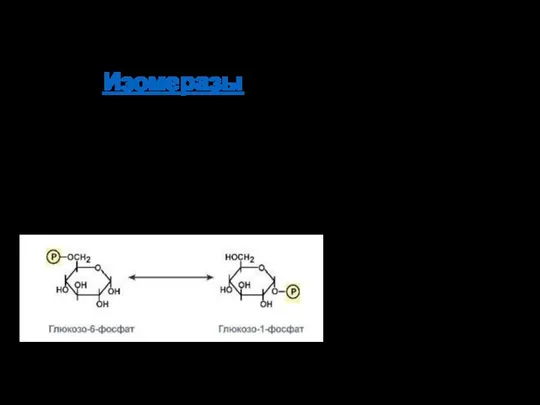
КФ 5: Изомеразы
Ферменты, катализирующие структурные превращения изомеров (рацемизация или эпимеризация).
Изомеразы – ферменты, катализирующие изомерные превращения
КФ 5: Изомеразы
Ферменты, катализирующие структурные превращения изомеров (рацемизация или эпимеризация).
Изомеразы – ферменты, катализирующие изомерные превращения

КФ 5: Изомеразы
Если рассматривать все подклассы, то изомеразы делятся по типу изомеризации на:
КФ 5: Изомеразы
Если рассматривать все подклассы, то изомеразы делятся по типу изомеризации на:

КФ 6: Лигазы
Ферменты, катализирующие соединение двух молекул с образованием новой химической связи (лигирование).
При
КФ 6: Лигазы
Ферменты, катализирующие соединение двух молекул с образованием новой химической связи (лигирование).
При

Лигазы в генной инженерии
Иными словами, ДНК-лигазы сшивают рядом расположенные нуклеотиды, образуя
Лигазы в генной инженерии
Иными словами, ДНК-лигазы сшивают рядом расположенные нуклеотиды, образуя
Информация о ферментах в сети Internet
Информация о ферментах в сети Internet
Информация о ферментах в сети Internet
Информация о ферментах в сети Internet
Информация о ферментах в сети Internet
Информация о ферментах в сети Internet
Информация о ферментах в сети Internet
Информация о ферментах в сети Internet

Лактатдегидрогеназа (ЛДГ)
Лактатдегидрогеназа (ЛДГ; L-лактат-НАД-оксидоредуктаза, КФ 1.1.1.27) - цинксодержаший фермент, катализирует обратимую
Лактатдегидрогеназа (ЛДГ)
Лактатдегидрогеназа (ЛДГ; L-лактат-НАД-оксидоредуктаза, КФ 1.1.1.27) - цинксодержаший фермент, катализирует обратимую

Значимость в диагностике
Клиническое значение определения активности ЛДГ
Активность ЛДГ в сыворотке крови
Значимость в диагностике
Клиническое значение определения активности ЛДГ Активность ЛДГ в сыворотке крови

.Креатинкиназа (КК)
Креатинкиназа-АТФ: креатин-1,4-фосфотрансфераза (КФ 2.7.3.2) — катализирует обратимый перенос фосфатного остатка
.Креатинкиназа (КК)
Креатинкиназа-АТФ: креатин-1,4-фосфотрансфераза (КФ 2.7.3.2) — катализирует обратимый перенос фосфатного остатка

у-Глутамилтранспептидаза
ГГТ; у-глутамил-транспептидазы, КФ 2.3.2.2).
ГГТ катализирует перенос у-глутамила на аминокислоту или пептид,
у-Глутамилтранспептидаза
ГГТ; у-глутамил-транспептидазы, КФ 2.3.2.2). ГГТ катализирует перенос у-глутамила на аминокислоту или пептид,

Значение в диагностике
Наиболее частая причина повышения активности ГГТ в сыворотке крови
Значение в диагностике
Наиболее частая причина повышения активности ГГТ в сыворотке крови
 Социальное медицинское страхование в Республике Казахстан
Социальное медицинское страхование в Республике Казахстан Балалар мен жасөспірімдердің дене тәрбиесі мен шынығуының физикалық-гигиеналық негіздері
Балалар мен жасөспірімдердің дене тәрбиесі мен шынығуының физикалық-гигиеналық негіздері Метаболикалық синдромның биохимиясы
Метаболикалық синдромның биохимиясы ppt
ppt District sales manager, Novo Nordisk
District sales manager, Novo Nordisk 3D принтер в медицине
3D принтер в медицине Методы определения подлинности лекарственного растительного сырья. Занятие №2
Методы определения подлинности лекарственного растительного сырья. Занятие №2 Ампутация в хирургической практике
Ампутация в хирургической практике Лямблиоз. Клиника
Лямблиоз. Клиника Ғалымдар қоршаған ортадан келетін токсиндер ағзаның қартаюының жеделдетілуінің негізгі себептерінің бірі екенін анықтады
Ғалымдар қоршаған ортадан келетін токсиндер ағзаның қартаюының жеделдетілуінің негізгі себептерінің бірі екенін анықтады Инструментальные методы исследования сердца
Инструментальные методы исследования сердца Патронаж здорового новорожденного на дому с оформлением учебной амбулаторной карты
Патронаж здорового новорожденного на дому с оформлением учебной амбулаторной карты Аритмии сердца: ЭКГ-диагностика
Аритмии сердца: ЭКГ-диагностика Эстрогендерді қабылдау мен әйелдерде эдометрийдің қатерлі ісігінің дамуы арасындағы байланыс
Эстрогендерді қабылдау мен әйелдерде эдометрийдің қатерлі ісігінің дамуы арасындағы байланыс Точечный массаж
Точечный массаж Microscopic colitis
Microscopic colitis Интенсивная терапия осложненных форм СД, основы КЩС
Интенсивная терапия осложненных форм СД, основы КЩС Қазақстан республикасындағы психиатриялық көмекті ұйымдастыру
Қазақстан республикасындағы психиатриялық көмекті ұйымдастыру Порядок проведения обязательных медицинских осмотров
Порядок проведения обязательных медицинских осмотров Пренатальное развитие ребёнка
Пренатальное развитие ребёнка Шылым шегу
Шылым шегу Догляд за хворими із захворюваннями та ушкодженнями органів черевної порожнини
Догляд за хворими із захворюваннями та ушкодженнями органів черевної порожнини Синдром слабости синусового узла
Синдром слабости синусового узла Цереброваскулярные заболевания. Причины роста сосудистых заболеваний
Цереброваскулярные заболевания. Причины роста сосудистых заболеваний Балалардың физикалық критерийлер анықтау
Балалардың физикалық критерийлер анықтау Клеточные факторы врождённого иммунитета. Фагоцитоз и его стадии
Клеточные факторы врождённого иммунитета. Фагоцитоз и его стадии Тік ішектің және анустың атрезиясы
Тік ішектің және анустың атрезиясы Конусно-лучевая компьютерная томография в терапевтической стоматологии
Конусно-лучевая компьютерная томография в терапевтической стоматологии