- Физиология нервной системы. Часть 1

Содержание
- 2. Нейрон как структурная и функциональная единица ЦНС. Нейрон состоит из тела клетки (сомы) и отростков (один
- 3. Функция дендритов - в передаче информации к телу клетки (связь с другими нейронами и афферентация с
- 4. Функции нейронов: афферентная (восприятие сигналов от других нервных клеток, из внешней и внутренней среды), эфферентная (генерация
- 5. Связи между нейронами осуществляются посредством синапсов. В ЦНС распространены химические, электрические, смешанные. По месту контакта синапсы
- 6. Химический синапс
- 7. РЕФЛЕКС КАК ОСНОВНОЙ АКТ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ Рефлекс-ответная реакция организма на раздражение из внешней и внутренней среды,
- 8. ИСТОРИЯ РАЗВИТИЯ РЕФЛЕКТОРНОЙ ТЕОРИИ Рене Декарт (1596—1650). Предположил, что при воздействии внешнего раздражителя (ожог пламенем свечи)
- 9. Классификация рефлексов По месту расположения рецепторов: интеро-, зкстеро-, проприорецептивные. По характеру рефлекторного действия: моторные, секреторные, сосудодвигательные.
- 10. Универсальные принципы рефлекторной теории По И.П.Павлову любой рефлекторный акт, независимо от его сложности, подчиняется трем принципам:
- 11. Нервный центр и его свойства Нервный центр - совокупность нейронов, участвующих в обеспечении регуляции определенной функции
- 12. СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ Односторонняя проводимость. Объясняется односторонней проводимостью химических синапсов. Задержка проведения возбуждения. Определяется суммацией синаптических
- 13. Окклюзия и центральное облегчение - разные варианты суммации возбуждения, хотя их выделяют как свойства нервных центров.
- 14. Координирующие принципы функционирования ЦНС 1.Принцип конвергенции - схождение возбуждения от многих нейронов к одной нейронной группе.
- 15. Координирующие принципы функционирования ЦНС 3. Принцип обратной связи. За счет обратной связи нейроны могут регулировать поток
- 16. СВОЙСТВА ДОМИНАНТНОГО ОЧАГА Повышенная возбудимость. Порог возбуждения доминирующего нервного центра снижается. Его возбуждение усиливается не только
- 17. ТОРМОЖЕНИЕ В ЦНС По механизму действия
- 18. Пресинаптическое торможение - тормозной нейрон образует аксоаксональный синапс на терминалах аксона возбуждающего нейрона. Медиатором в этом
- 20. Скачать презентацию
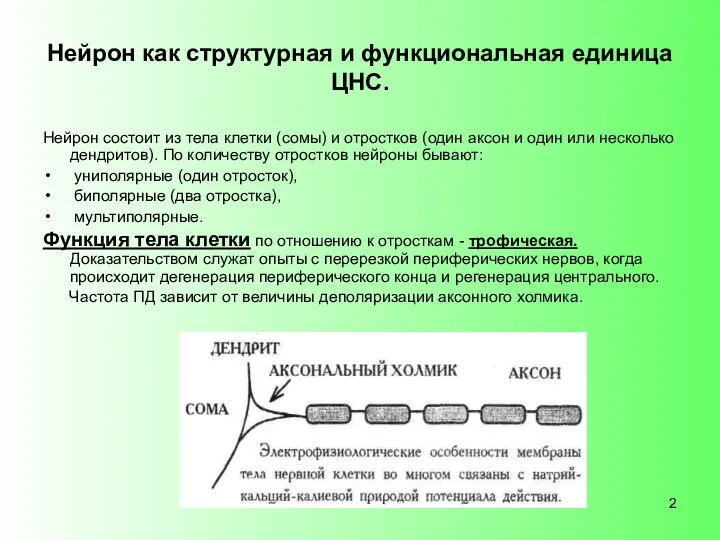
Нейрон как структурная и функциональная единица ЦНС.
Нейрон состоит из тела клетки
Нейрон как структурная и функциональная единица ЦНС.
Нейрон состоит из тела клетки

Функция дендритов - в передаче информации к телу клетки (связь с
Функция дендритов - в передаче информации к телу клетки (связь с

Функции нейронов:
афферентная (восприятие сигналов от других нервных клеток, из внешней и
Функции нейронов:
афферентная (восприятие сигналов от других нервных клеток, из внешней и

Связи между нейронами осуществляются посредством синапсов.
В ЦНС распространены химические, электрические,
Связи между нейронами осуществляются посредством синапсов.
В ЦНС распространены химические, электрические,
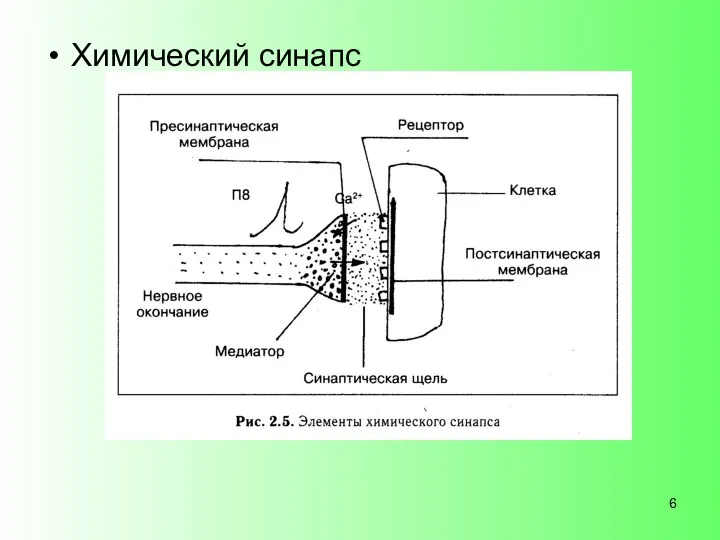
Химический синапс
Химический синапс

РЕФЛЕКС КАК ОСНОВНОЙ АКТ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
Рефлекс-ответная реакция организма на раздражение из
РЕФЛЕКС КАК ОСНОВНОЙ АКТ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
Рефлекс-ответная реакция организма на раздражение из

ИСТОРИЯ РАЗВИТИЯ РЕФЛЕКТОРНОЙ ТЕОРИИ
Рене Декарт (1596—1650). Предположил, что при воздействии внешнего
ИСТОРИЯ РАЗВИТИЯ РЕФЛЕКТОРНОЙ ТЕОРИИ
Рене Декарт (1596—1650). Предположил, что при воздействии внешнего

Классификация рефлексов
По месту расположения рецепторов:
интеро-,
зкстеро-,
проприорецептивные.
По
Классификация рефлексов
По месту расположения рецепторов:
интеро-,
зкстеро-,
проприорецептивные.
По

Универсальные принципы рефлекторной теории
По И.П.Павлову любой рефлекторный акт, независимо от его
Универсальные принципы рефлекторной теории
По И.П.Павлову любой рефлекторный акт, независимо от его

Нервный центр и его свойства
Нервный центр - совокупность нейронов, участвующих в
Нервный центр и его свойства
Нервный центр - совокупность нейронов, участвующих в

СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
Односторонняя проводимость. Объясняется односторонней проводимостью химических синапсов.
Задержка проведения возбуждения.
СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
Односторонняя проводимость. Объясняется односторонней проводимостью химических синапсов.
Задержка проведения возбуждения.
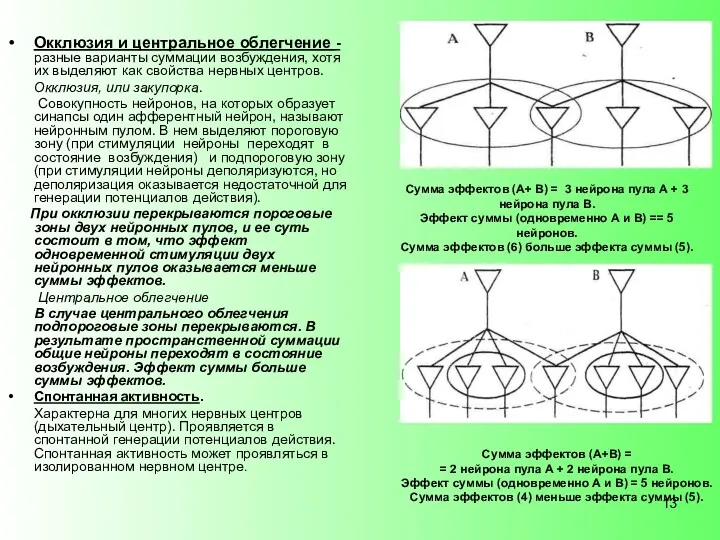
Окклюзия и центральное облегчение - разные варианты суммации возбуждения, хотя их
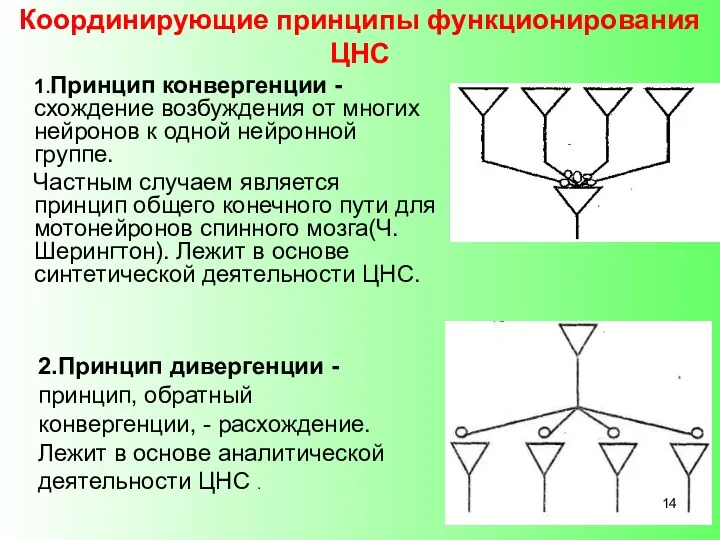
Координирующие принципы функционирования ЦНС
1.Принцип конвергенции - схождение возбуждения от
Координирующие принципы функционирования ЦНС
1.Принцип конвергенции - схождение возбуждения от

Координирующие принципы функционирования ЦНС
3. Принцип обратной связи. За счет обратной
Координирующие принципы функционирования ЦНС
3. Принцип обратной связи. За счет обратной

СВОЙСТВА ДОМИНАНТНОГО ОЧАГА
Повышенная возбудимость. Порог возбуждения доминирующего нервного центра снижается.
СВОЙСТВА ДОМИНАНТНОГО ОЧАГА
Повышенная возбудимость. Порог возбуждения доминирующего нервного центра снижается.
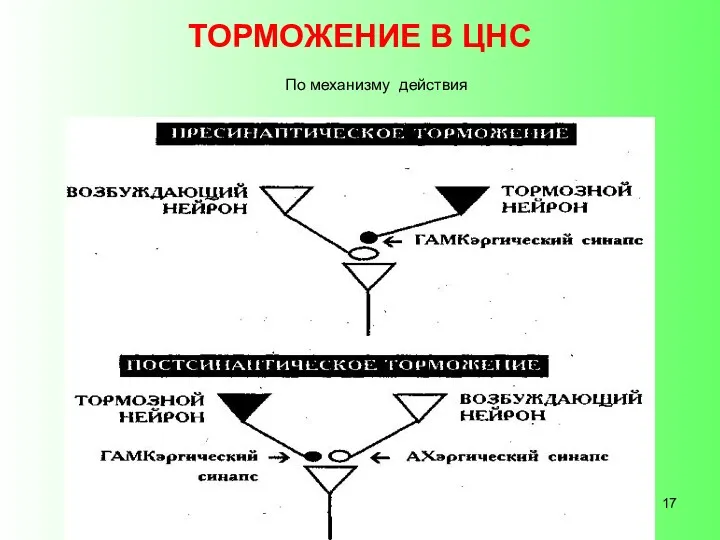
ТОРМОЖЕНИЕ В ЦНС
По механизму действия
ТОРМОЖЕНИЕ В ЦНС
По механизму действия

Пресинаптическое торможение - тормозной нейрон образует аксоаксональный синапс на терминалах аксона
Пресинаптическое торможение - тормозной нейрон образует аксоаксональный синапс на терминалах аксона
 Санитарно-эпидемиологические требования к организациям, осуществляющим медицинскую деятельность
Санитарно-эпидемиологические требования к организациям, осуществляющим медицинскую деятельность Фурункул и карбункул лица. Рожистое воспаление лица. Клиника, диагностика и лечение
Фурункул и карбункул лица. Рожистое воспаление лица. Клиника, диагностика и лечение Ранние токсикозы беременных
Ранние токсикозы беременных Побочное действие химиотерапевтических лс
Побочное действие химиотерапевтических лс Принципы организации профессиональной реабилитации. Бригадный метод в реабилитации больных и инвалидов
Принципы организации профессиональной реабилитации. Бригадный метод в реабилитации больных и инвалидов Эффективный перинатальный уход в неонатологии: основные принципы
Эффективный перинатальный уход в неонатологии: основные принципы Жасқа байланысты,асқынбаған тіс жегісінде,жүйелі аурулар мен зат алмасу ауруларында ұлпадағы өзгерістер
Жасқа байланысты,асқынбаған тіс жегісінде,жүйелі аурулар мен зат алмасу ауруларында ұлпадағы өзгерістер Врожденные пороки развития черепа
Врожденные пороки развития черепа Строение органов мочевыделительной системы
Строение органов мочевыделительной системы Интеграция в психотерапии. Использование уникальных условий океанской среды для лечения расстройств
Интеграция в психотерапии. Использование уникальных условий океанской среды для лечения расстройств Нерв жүйесінің физиологиясы. Жоғары нерв әрекеті, олардың жас ерекшелігі
Нерв жүйесінің физиологиясы. Жоғары нерв әрекеті, олардың жас ерекшелігі Развитие общественной медицины в 18-19 вв
Развитие общественной медицины в 18-19 вв Вірусні дерматози. Папулосквамозні дерматози
Вірусні дерматози. Папулосквамозні дерматози Актуальные проблемы комплексной реабилитации инвалидов на современном этапе
Актуальные проблемы комплексной реабилитации инвалидов на современном этапе Өкпе рагы
Өкпе рагы Первая медицинская помощь при отравлениях
Первая медицинская помощь при отравлениях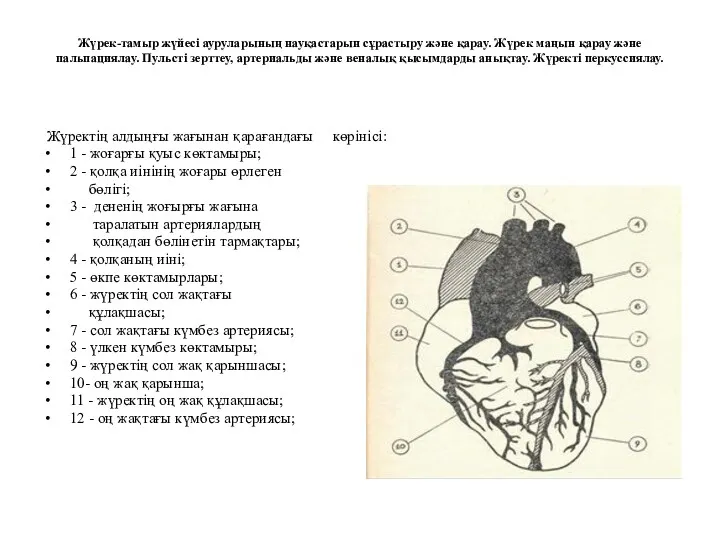 Жүрек-тамыр жүйесі ауруларының науқастарын сұрастыру және қарау. Жүрек маңын қарау және пальпациялау
Жүрек-тамыр жүйесі ауруларының науқастарын сұрастыру және қарау. Жүрек маңын қарау және пальпациялау Детский церебральный паралич. Реабилитационный маршрут ребёнка с ДЦП. Цели реабилитации
Детский церебральный паралич. Реабилитационный маршрут ребёнка с ДЦП. Цели реабилитации Заикание. Этиология заикания
Заикание. Этиология заикания Аффективные расстройства. Расстройства воли и влечений
Аффективные расстройства. Расстройства воли и влечений Цитостатические препараты
Цитостатические препараты Тромбоэмболия легочной артерии
Тромбоэмболия легочной артерии Лица науки. Константин Иванович Скрябин
Лица науки. Константин Иванович Скрябин Вибрация. Вибрационная патология и ее предупреждение
Вибрация. Вибрационная патология и ее предупреждение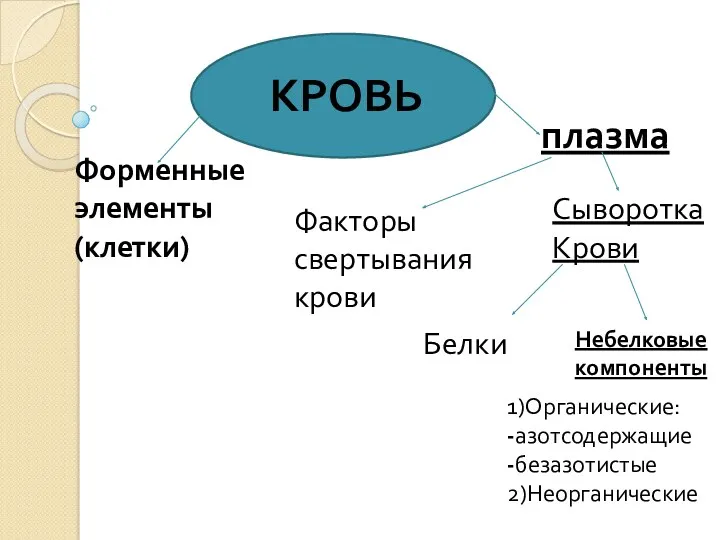 Биохимия крови
Биохимия крови Эволюция представлений о сепсисе. Сепсис-3. Современная концепция патогенеза сепсиса
Эволюция представлений о сепсисе. Сепсис-3. Современная концепция патогенеза сепсиса Развитие спортивно-оздоровительного туризма в России
Развитие спортивно-оздоровительного туризма в России Система органов кроветворения
Система органов кроветворения