- Гуморальный иммунный ответ. Основные этапы развития. Цикл 1 – иммунология

Содержание
- 2. Выход зрелых В клеток на периферию Из костного мозга выходят только те В –лимфоциты, у которых
- 3. Y Y Y Y Y Y Зрелый В-лимфоцит выходит на периферию Y Y В - клеточная
- 4. Антигенраспознающие участки рецепторов В лимфоцитов (ВCR) и рецепторов Т-лимфоцитов (TCR) Антиген-распознающие участки рецепторов как Т, так
- 5. Постулаты теории клональной селекции Каждый В лимфоцит имеет рецептор уникальной специфичности. Высокоаффинное (прочное) взаимодействие рецептора с
- 6. Антигензависимая дифференцировка В лимфоцитов Плазматические клетки синтезируют имуноглобулины –антитела, распознающие своими Fab-фрагментами антигены. Наряду с В-
- 7. Гены, кодирующие молекулы иммуноглобулинов (Ig) Синтез иммуноглобулинов значительно отличается от синтеза других белков: вариабельные (V) и
- 8. Строение молекулы иммуноглобулинов (Ig) Каждая молекула Ig состоит из: двух тяжелых (Н) и двух легких (L)
- 9. Полярность молекулы Ig: Fab и Fc -фрагменты Fab-фрагмент(antigen binding)-уникальная для каждого Ig часть молекулы Легкая цепь,
- 10. Фазы развития гуморального иммунного ответа
- 11. Периферия: расположение, строение и функция лимфатических узлов(ЛУ) Лимфоузлы находятся на стыке лимфатических и кровеносных сосудов: по
- 12. Функции вторичных лимфоидных органов «Входов» в ЛУ намного больше, чем «выходов», то есть ЛУ –своеобразные фильтры,
- 13. Строение ЛУ Афферентный (приносящий) и эфферентный (выносящий) лимфатические сосуды. Снаружи капсула, под капсулой - кора –
- 14. Жизненный цикл В - лимфоцитов на периферии - в ЛУ В - клетки покидают сосуды и
- 15. Рециркулирующие В – клетки встречаются со «своим» антигеном в лимфоузле
- 16. Биологический смысл соматических гипермутаций В зародышевых центрах интенсивная пролиферация В-клеток сочетается: с соматическими гипермутациями реаранжированных ранее
- 17. События в лимфоузле – посте встречи В лимфоцита с комплементарным его рецептору антигеном После соматических гипермутаций
- 18. J J O’Shea, and W E Paul Science 2010;327:1098-1102 Развитие Т-хелперов различных типов под влиянием цитокиновых
- 19. Тип иммунного ответа определяет цитокиновое микроокружение, которое для наивных Тхелперов (Th 0) создают активированные антигеном дендритные
- 20. Сопоставление характеристик В-лимфоцита и плазматической клетки уровень экспрессии рост соматические переключение Ig MHC II Ig секреция
- 21. Домены –компактно уложенные структуры, устойчивые к действию протеаз; по типу тяжелых цепей называются классы иммуноглобулинов α-цепь-(IgA);
- 22. Функции (Fab – antigen binding) фрагментов в молекулах Ig Распознавать антиген. Преципитировать антиген. Запускать процессинг антигенов.
- 23. Fv Гибкость иммуноглобули новых молекул
- 24. Классы иммуноглобулинов(Ig) и их содержание в сыворотке крови * Cреди класса IgG субклассы: IgG1 -65, IgG2-23,
- 25. IgМ Синтезируются первыми, составляют 6-10% от всех иммуноглобулинов сыворотки крови. Короткий период синтеза и полураспада (4-
- 26. Мономерный IgM На поверхности В клетки IgM существует только как мономер мономерный IgM имеет очень низкую
- 27. МУЛЬТИМЕРИЗАЦИЯ: из 5 молекул IgM-формируется пентамер 1. 2 мономера IgM (показаны только Fc) 2. Цистеины в
- 28. Изменение конформации молекулы IgM, вызванное связыванием Fab –фрагментов с антигеном
- 29. Иммуноглобулины класса М - IgM Тяжелая цепь: μ - Mю Время полужизни: 5 - 10 дней
- 30. IgG – мономер Самое высокое содержание в крови - до 80% от общего числа всех Ig.
- 31. Имуноглобулины класса G - IgG Тяжелые цепи: γ 1 γ 2 γ3 γ4 - гамма 1
- 32. IgA- димер Выделяют 2 субкласса: IgA1 (90%) и IgA2 (10%). Содержание IgA в сыворотке крови составляет
- 33. И.И.Мечников и А.М.Безредка - учение о местном иммунитете
- 34. IgA -мономерный, димерный и и секреторный Существует 2 субкласса IgA IgA1 в основном присутствует в сыворотке
- 35. Cекреторный IgA (sIgA) - трансцитоз комплекс pIgR разрушается, освобождается IgA с частью pIgR - секреторным компонентом
- 36. Места продукции секреторного IgA в организме человека рот бронхи кишка вагина кожа
- 37. Секреторный IgA Продукция в организме взрослого человека - 2,5-3,0 грамма в сутки. 80% всех плазматических клеток
- 38. ФУНКЦИИ sIgA Нейтрализация вирусов и токсинов Иммунная экскреция антигенов Внутриклеточная нейтрализация вирусов
- 39. Иммунное исключение (агглютинация вирусов и бактерий) в просвете пищеварительного тракта. ФУНКЦИИ sIgA sIgA, взаимодействуя с бактериями,
- 40. Иммуноглобулины класса А - IgA Тяжелые цепи: α1 или α2 - альфа 1или 2 Время полужизни:
- 41. IgE - мономер В сыворотке крови крайне мало - 0,002% от общего количества Ig (введены специальные
- 42. Высокоаффинные IgE рецепторы (FcεRI) IgE - FcεRI взаимодействия являются самыми устойчивыми , у них самая прочная
- 43. Иммуноглобулины класса Е - IgE IgE появился позже в эволюции в связи с его ролью в
- 44. . IgD - мономер Является антигенспецифичным рецептором на поверхности В-лимфоцитов. Биологическое значение до конца не определено.
- 45. Иммуноглобулины класса D - IgD IgD ко-экспрессируется вместе с IgM на В клетке в результате альтернативного
- 46. Различия в аминокислотной последовательности молекул антител Изотипические различия между иммуноглобулиновыми молекулами: у каждого изотипа Ig -одинаковая
- 47. Роль антигенпрезентирующих клеток (АПК) в активации В лимфоцитов при ответе на Т- зависимые антигены (большинство белковых
- 48. Взаимодействие Т и В лимфоцитов при ответе на Т- зависимые антигены (белковые АГ) Мембранносвязанный IgR(рецептор) на
- 49. Взаимодействие Т и В лимфоцитов при ответе на Т- зависимые антигены Комплекс MHC II +АГ пептид
- 50. 2. Связывание и интернализация через Ig вызывает экспрессию CD40 3. Антиген подвергается процессингу 4. Пептидные фрагменты
- 51. ДК презентируют РАЗЛИЧНЫЕ АНТИГЕНЫ Активация антигеном В лимфоцита – экспрессия молекулы CD40 Поверхностный Ig захватывает АГ
- 52. T клетки помогают B клеткам сигнал 1 антиген и рецептор к антигену 1. T клеточный рецептор
- 53. T клеточная помощь - сигнал 2 B клетки погибают путем апоптоза, однако сигналы1 и 2 повышают
- 54. Цитокиновая активация В клеток Активированные Т хелперы секретируют цитокины, активирующие В клетки. IL-2 –индуктор активации Т
- 55. Ответ В клеток на Т-независимые антигены При ответе на Т-независимые антигены В лимфоцитам не нужна помощь
- 56. Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y
- 57. Ответ В клеток на Т- независимые антигены В ответе В клеток на Т-независимые антигены отсутствует взаимодействие
- 58. B-1 клетки Распознают эпитопы АГ с повторяющимися структурами - фосфотидилхолин, липополисахариды и др. Не являются (?)
- 59. Эффекторные механизмы защиты против внеклеточных патогенов с помощью антител Каков биологический смысл и итог гуморального иммунного
- 60. Эффекторные механизмы защиты против внеклеточных патогенов НЕЙТРАЛИЗАЦИЯ Нейтрализующие антитела БАКТЕРИЯ ТОКСИН
- 61. Эффекторные механизмы защиты против внеклеточных патогенов ОПСОНИЗАЦИЯ ОПСОНИЗАЦИЯ
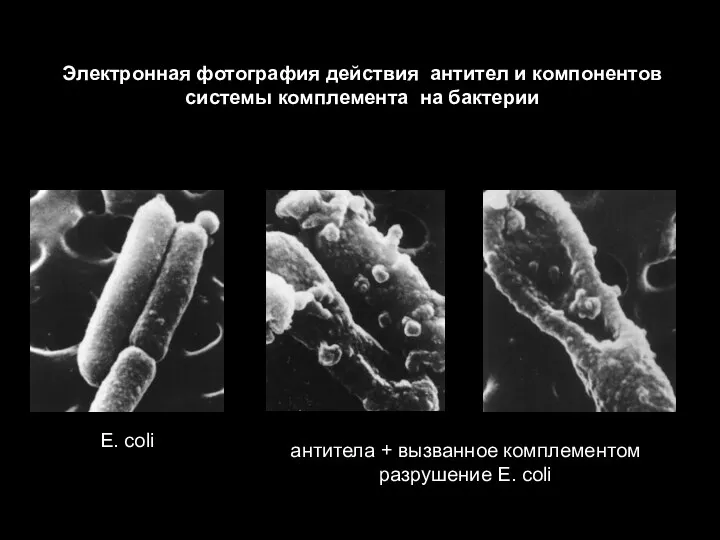
- 64. Эффекторные механизмы защиты против внеклеточных патогенов Активация системы комплемента лизис
- 66. Вопросы Фазы адаптивного иммунного ответа. Типы иммунного ответа и факторы, их определяющие. В-лимфоцит как антигенпредставляющая клетка.
- 67. Тестовые вопросы Антитела обладают способностью: Расщеплять антиген Активировать комплемент Специфически связывать антиген Фагоцитировать антиген Экспрессировать антиген
- 68. Тестовые вопросы К «цитофильным» иммуноглобулинам относятся: IgA IgG IgM IgE IgD Основными иммуноглобулинами вторичного ответа являются:
- 69. Тестовые вопросы Активировать систему комплемента по классическому пути способны: IgA IgG IgM IgE IgD Изотипы иммуноглобулинов
- 70. Тестовые вопросы Через плаценту проникает: IgA IgG IgM IgE IgD Основными иммуноглобулинами первичного ответа являются: IgA
- 72. Скачать презентацию

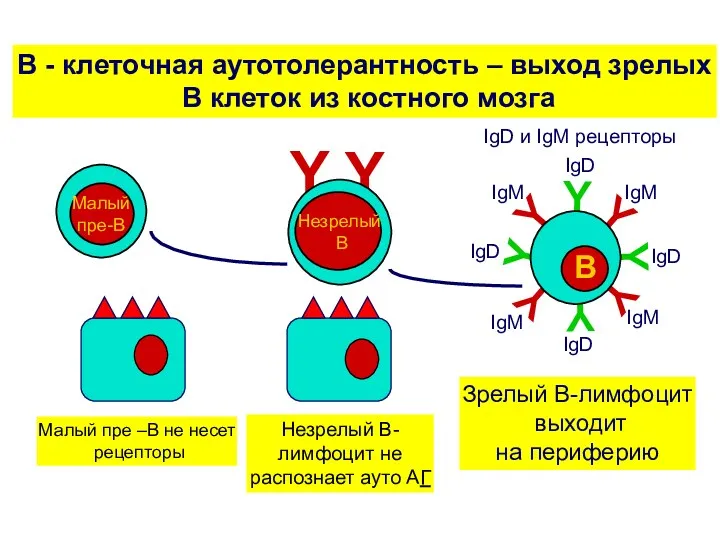
Выход зрелых В клеток на периферию
Из костного мозга выходят только те
Выход зрелых В клеток на периферию
Из костного мозга выходят только те
Y
Y
Y
Y
Y
Y
Зрелый В-лимфоцит
выходит
на периферию
Y
Y
В - клеточная аутотолерантность – выход зрелых
Y
Y
Y
Y
Y
Y
Зрелый В-лимфоцит
выходит
на периферию
Y
Y
В - клеточная аутотолерантность – выход зрелых
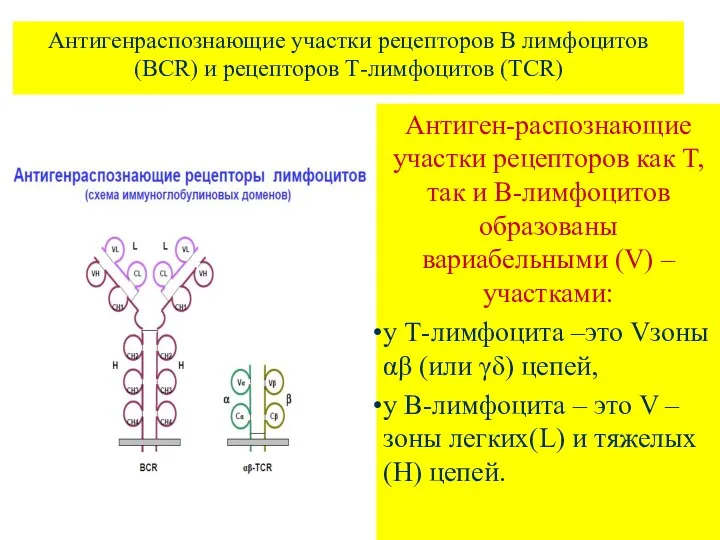
Антигенраспознающие участки рецепторов В лимфоцитов (ВCR) и рецепторов Т-лимфоцитов (TCR)
Антиген-распознающие участки
Антигенраспознающие участки рецепторов В лимфоцитов (ВCR) и рецепторов Т-лимфоцитов (TCR)
Антиген-распознающие участки

Постулаты теории клональной селекции
Каждый В лимфоцит имеет рецептор уникальной специфичности.
Высокоаффинное (прочное)
Постулаты теории клональной селекции
Каждый В лимфоцит имеет рецептор уникальной специфичности.
Высокоаффинное (прочное)

Антигензависимая дифференцировка В лимфоцитов
Плазматические клетки синтезируют имуноглобулины –антитела, распознающие своими Fab-фрагментами
Антигензависимая дифференцировка В лимфоцитов
Плазматические клетки синтезируют имуноглобулины –антитела, распознающие своими Fab-фрагментами
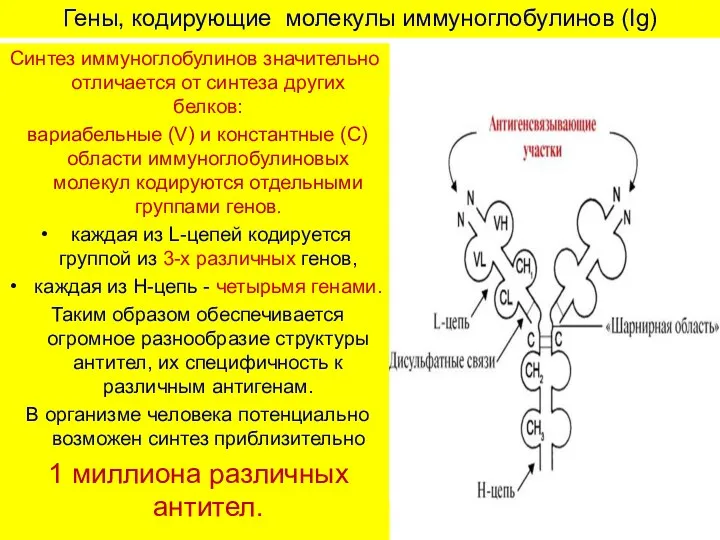
Гены, кодирующие молекулы иммуноглобулинов (Ig)
Синтез иммуноглобулинов значительно отличается от синтеза других
Гены, кодирующие молекулы иммуноглобулинов (Ig)
Синтез иммуноглобулинов значительно отличается от синтеза других
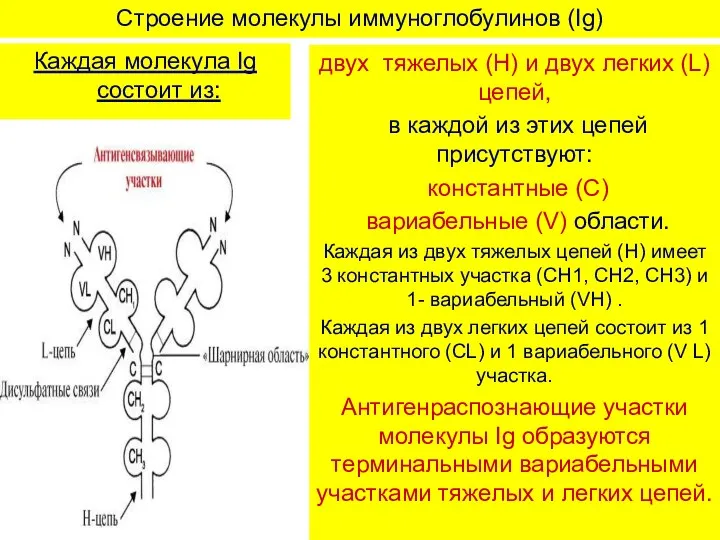
Строение молекулы иммуноглобулинов (Ig)
Каждая молекула Ig состоит из:
двух тяжелых (Н) и
Строение молекулы иммуноглобулинов (Ig)
Каждая молекула Ig состоит из:
двух тяжелых (Н) и

Полярность молекулы Ig: Fab и Fc -фрагменты
Fab-фрагмент(antigen binding)-уникальная для каждого Ig
Полярность молекулы Ig: Fab и Fc -фрагменты
Fab-фрагмент(antigen binding)-уникальная для каждого Ig
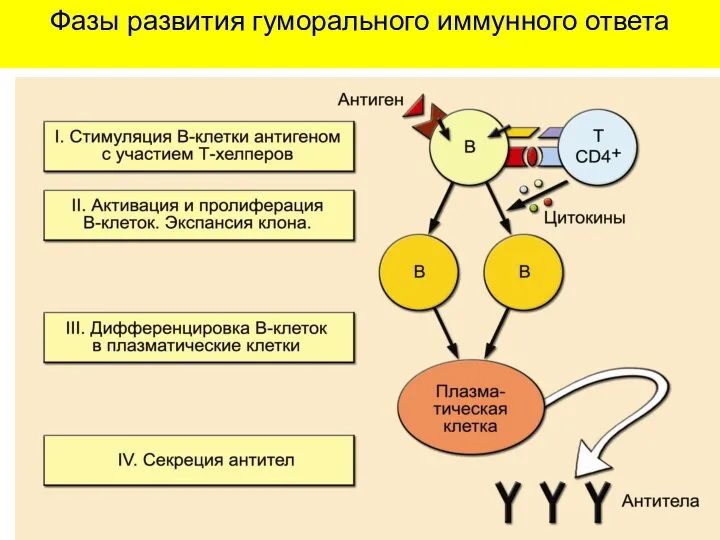
Фазы развития гуморального иммунного ответа
Фазы развития гуморального иммунного ответа

Периферия: расположение, строение и функция лимфатических узлов(ЛУ)
Лимфоузлы находятся на стыке лимфатических
Периферия: расположение, строение и функция лимфатических узлов(ЛУ)
Лимфоузлы находятся на стыке лимфатических

Функции вторичных лимфоидных органов
«Входов» в ЛУ намного больше, чем «выходов»,
Функции вторичных лимфоидных органов
«Входов» в ЛУ намного больше, чем «выходов»,
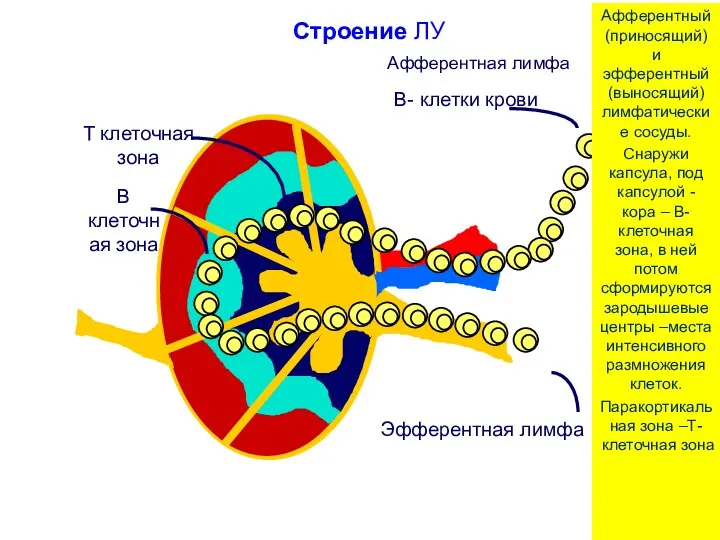
Строение ЛУ
Афферентный (приносящий) и эфферентный (выносящий) лимфатические сосуды.
Снаружи капсула, под
Строение ЛУ
Афферентный (приносящий) и эфферентный (выносящий) лимфатические сосуды.
Снаружи капсула, под

Жизненный цикл В - лимфоцитов на периферии - в ЛУ
В -
Жизненный цикл В - лимфоцитов на периферии - в ЛУ
В -
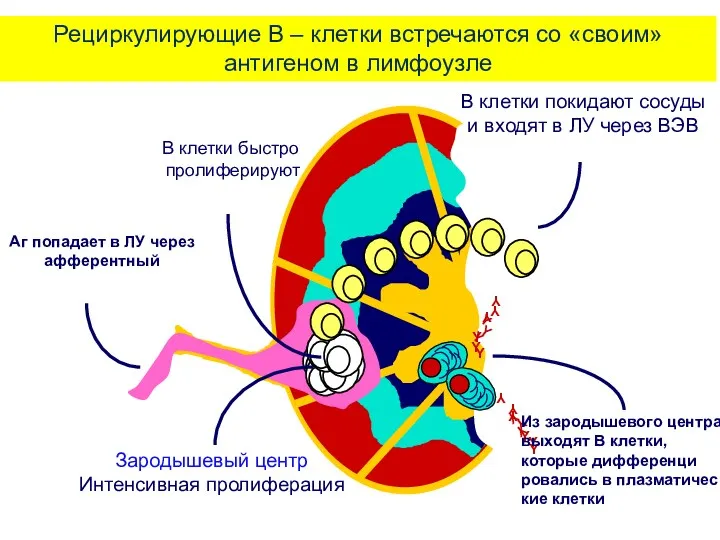
Рециркулирующие В – клетки встречаются со «своим» антигеном в лимфоузле
Рециркулирующие В – клетки встречаются со «своим» антигеном в лимфоузле

Биологический смысл соматических гипермутаций
В зародышевых центрах интенсивная пролиферация В-клеток сочетается:
с
Биологический смысл соматических гипермутаций
В зародышевых центрах интенсивная пролиферация В-клеток сочетается:
с

События в лимфоузле – посте встречи В лимфоцита с комплементарным его
События в лимфоузле – посте встречи В лимфоцита с комплементарным его
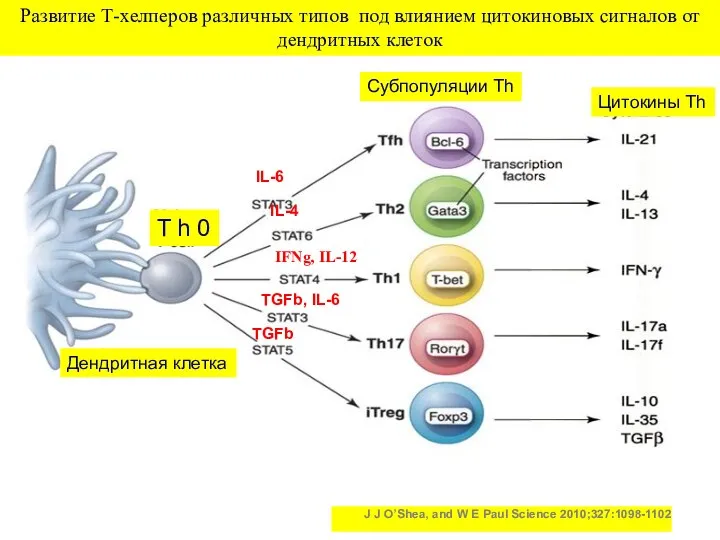
J J O’Shea, and W E Paul Science 2010;327:1098-1102
Развитие Т-хелперов различных
J J O’Shea, and W E Paul Science 2010;327:1098-1102
Развитие Т-хелперов различных
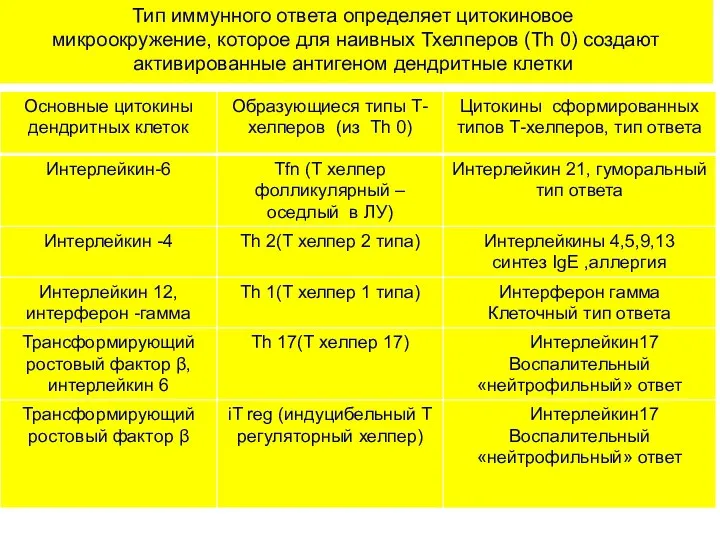
Тип иммунного ответа определяет цитокиновое микроокружение, которое для наивных Тхелперов (Th
Тип иммунного ответа определяет цитокиновое микроокружение, которое для наивных Тхелперов (Th
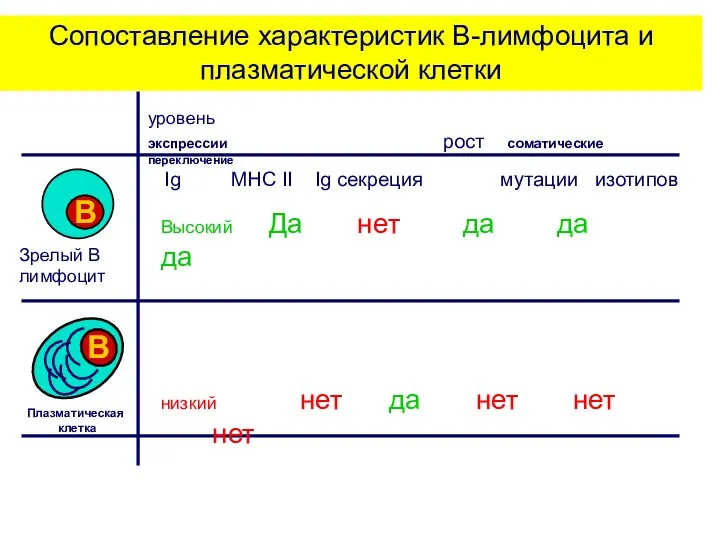
Сопоставление характеристик В-лимфоцита и плазматической клетки
уровень
экспрессии рост соматические переключение
Ig MHC
Сопоставление характеристик В-лимфоцита и плазматической клетки
уровень
экспрессии рост соматические переключение
Ig MHC
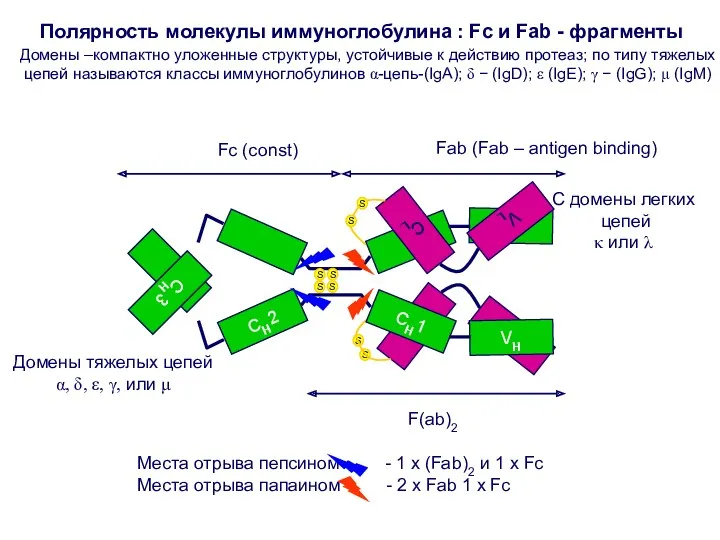
Домены –компактно уложенные структуры, устойчивые к действию протеаз; по типу тяжелых
Домены –компактно уложенные структуры, устойчивые к действию протеаз; по типу тяжелых

Функции (Fab – antigen binding) фрагментов в молекулах Ig
Распознавать антиген.
Преципитировать антиген.
Запускать
Функции (Fab – antigen binding) фрагментов в молекулах Ig
Распознавать антиген.
Преципитировать антиген.
Запускать
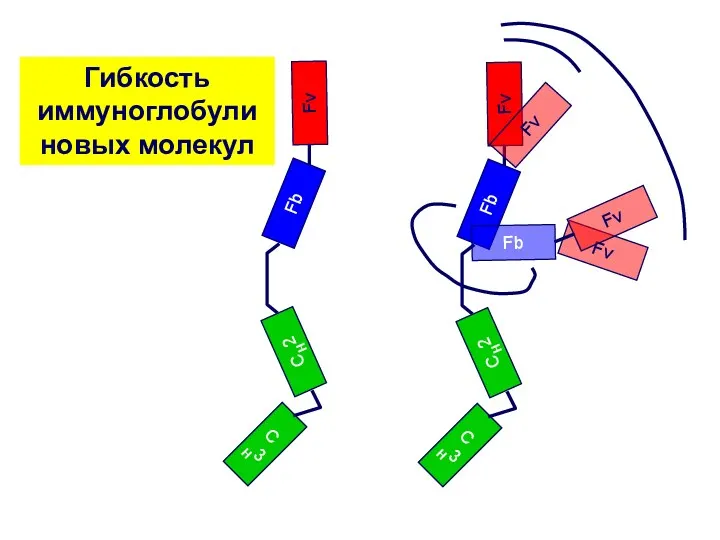
Fv
Гибкость иммуноглобули
новых молекул
Fv
Гибкость иммуноглобули
новых молекул
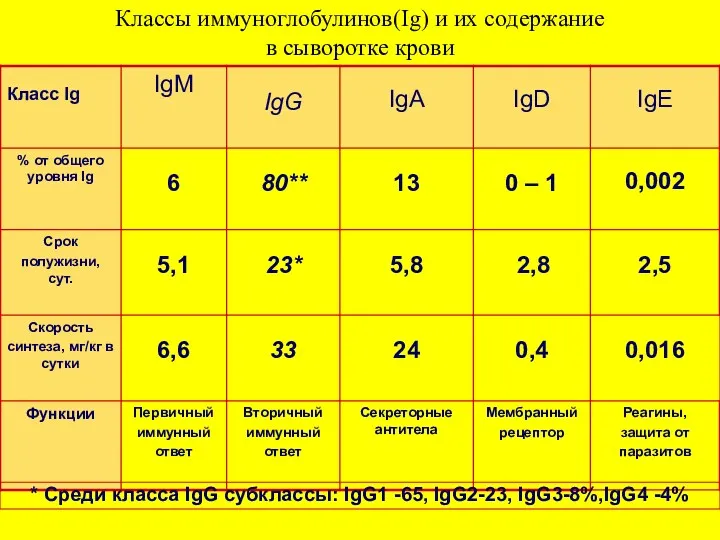
Классы иммуноглобулинов(Ig) и их содержание
в сыворотке крови
* Cреди класса IgG субклассы:
Классы иммуноглобулинов(Ig) и их содержание
в сыворотке крови
* Cреди класса IgG субклассы:

IgМ
Синтезируются первыми, составляют 6-10% от всех иммуноглобулинов сыворотки крови.
Короткий период синтеза
IgМ
Синтезируются первыми, составляют 6-10% от всех иммуноглобулинов сыворотки крови.
Короткий период синтеза
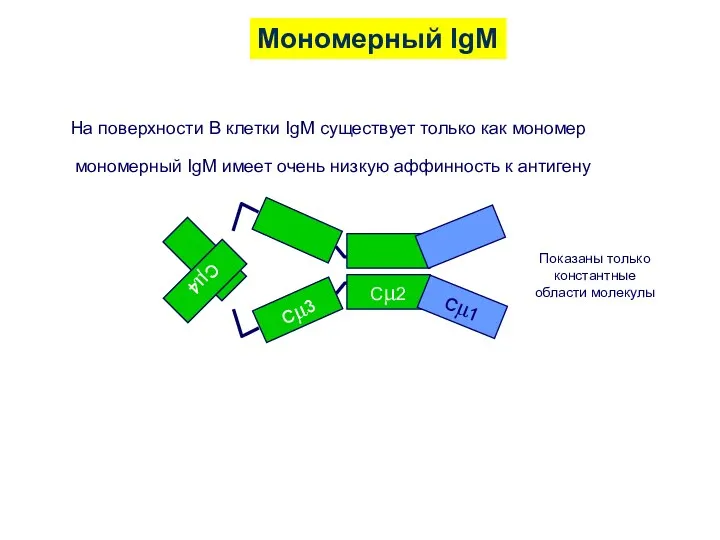
Мономерный IgM
На поверхности В клетки IgM существует только как мономер
мономерный IgM
Мономерный IgM
На поверхности В клетки IgM существует только как мономер
мономерный IgM
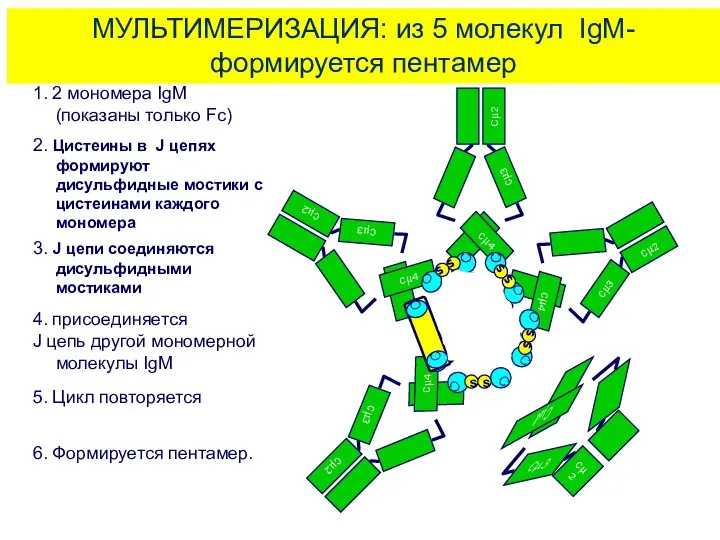
МУЛЬТИМЕРИЗАЦИЯ: из 5 молекул IgM-формируется пентамер
1. 2 мономера IgM
(показаны только Fc)
2.
МУЛЬТИМЕРИЗАЦИЯ: из 5 молекул IgM-формируется пентамер
1. 2 мономера IgM
(показаны только Fc)
2.

Изменение конформации молекулы IgM, вызванное связыванием Fab –фрагментов с антигеном
Изменение конформации молекулы IgM, вызванное связыванием Fab –фрагментов с антигеном
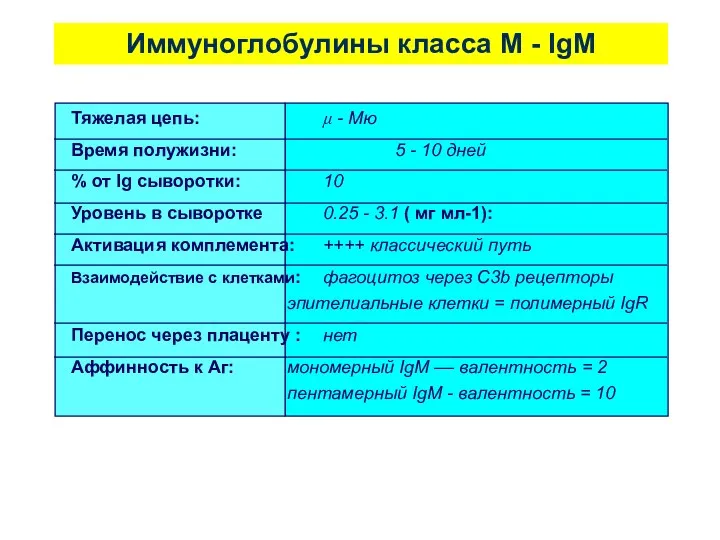
Иммуноглобулины класса М - IgM
Тяжелая цепь: μ - Mю
Время полужизни: 5 -
Иммуноглобулины класса М - IgM
Тяжелая цепь: μ - Mю
Время полужизни: 5 -

IgG – мономер
Самое высокое содержание в крови - до 80%
IgG – мономер
Самое высокое содержание в крови - до 80%
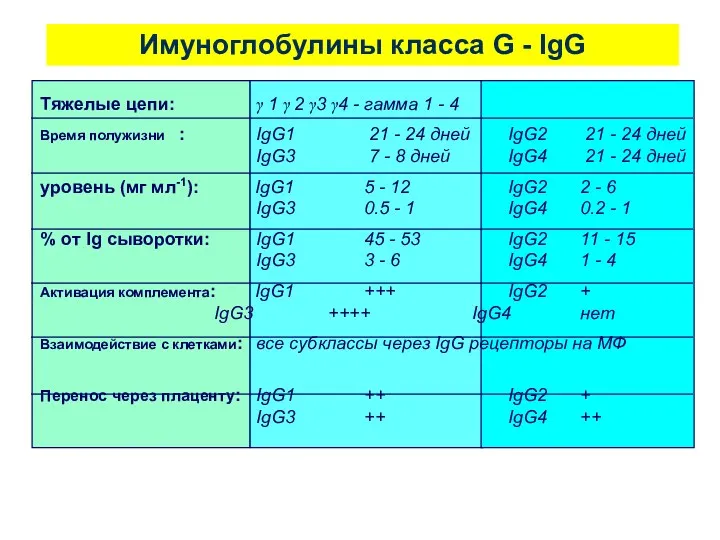
Имуноглобулины класса G - IgG
Тяжелые цепи: γ 1 γ 2 γ3 γ4
Имуноглобулины класса G - IgG
Тяжелые цепи: γ 1 γ 2 γ3 γ4

IgA- димер
Выделяют 2 субкласса: IgA1 (90%) и IgA2 (10%).
Содержание IgA в
IgA- димер
Выделяют 2 субкласса: IgA1 (90%) и IgA2 (10%).
Содержание IgA в

И.И.Мечников и А.М.Безредка - учение о местном иммунитете
И.И.Мечников и А.М.Безредка - учение о местном иммунитете
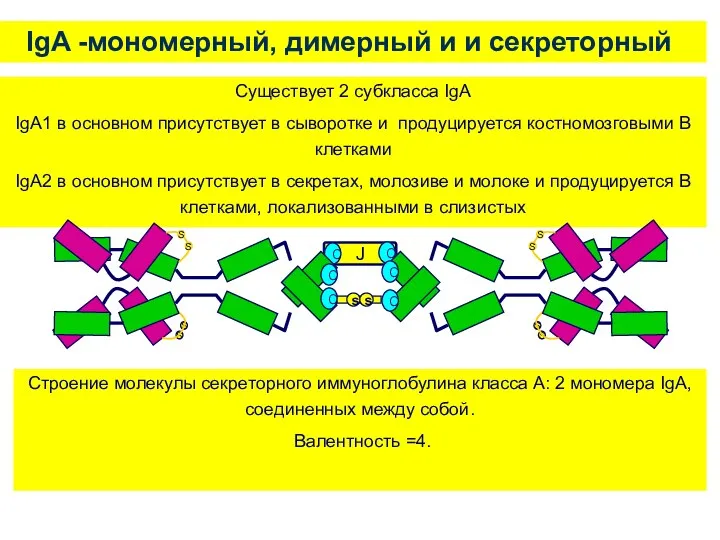
IgA -мономерный, димерный и и секреторный
Существует 2 субкласса IgA
IgA1
IgA -мономерный, димерный и и секреторный
Существует 2 субкласса IgA
IgA1
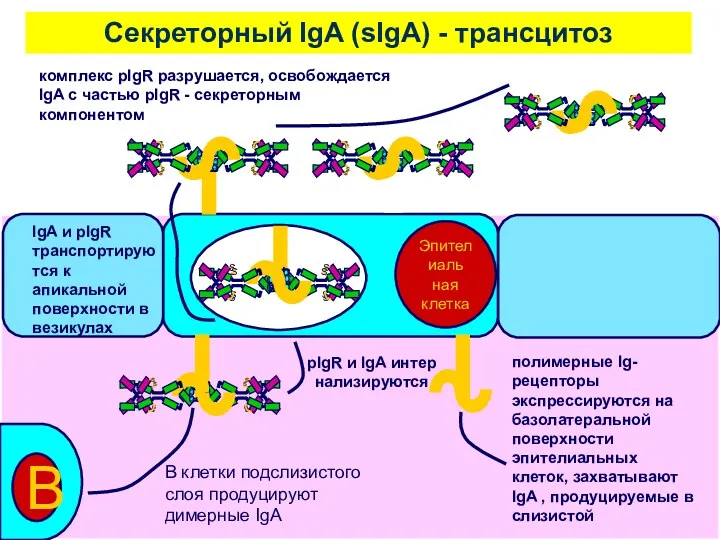
Cекреторный IgA (sIgA) - трансцитоз
комплекс pIgR разрушается, освобождается IgA с частью
Cекреторный IgA (sIgA) - трансцитоз
комплекс pIgR разрушается, освобождается IgA с частью
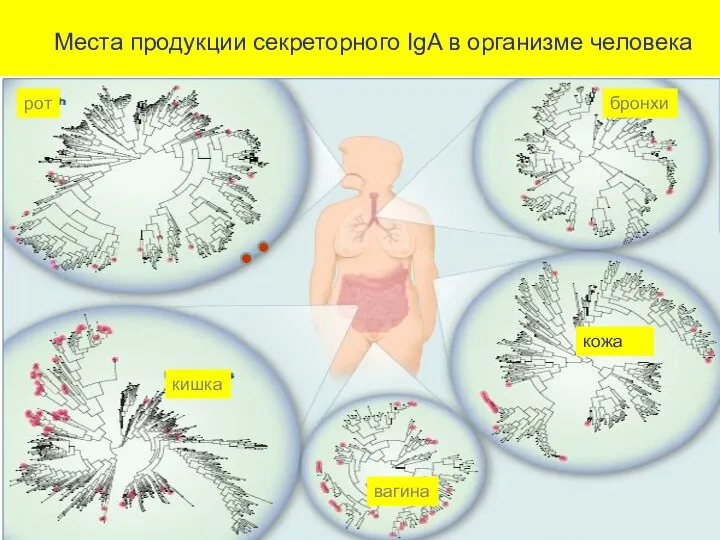
Места продукции секреторного IgA в организме человека
рот
бронхи
кишка
вагина
кожа
Места продукции секреторного IgA в организме человека
рот
бронхи
кишка
вагина
кожа
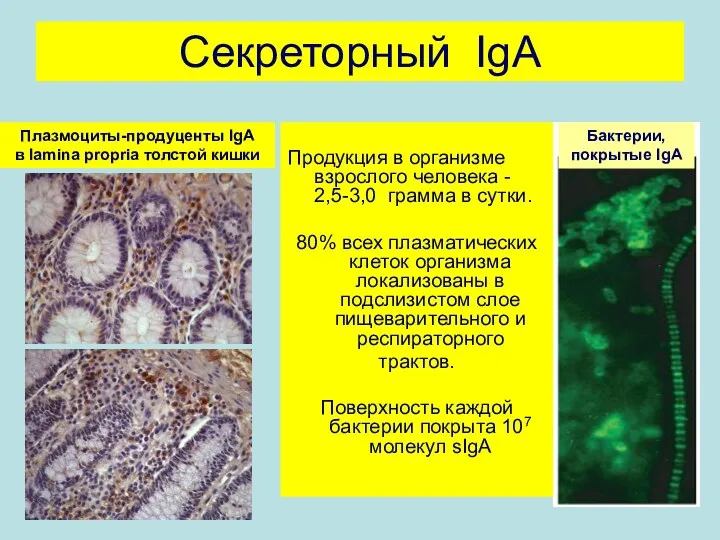
Секреторный IgA
Продукция в организме взрослого человека - 2,5-3,0 грамма в сутки.
80%
Секреторный IgA
Продукция в организме взрослого человека - 2,5-3,0 грамма в сутки.
80%
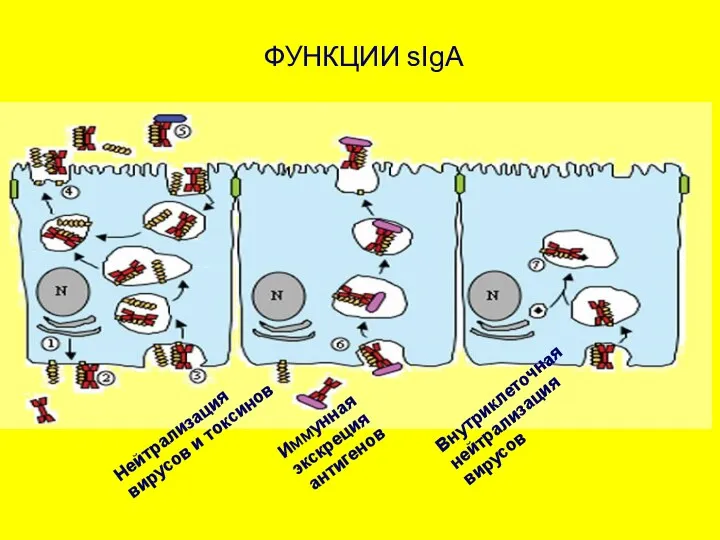
ФУНКЦИИ sIgA
Нейтрализация
вирусов и токсинов
Иммунная
экскреция
антигенов
Внутриклеточная
нейтрализация
вирусов
ФУНКЦИИ sIgA
Нейтрализация
вирусов и токсинов
Иммунная
экскреция
антигенов
Внутриклеточная
нейтрализация
вирусов
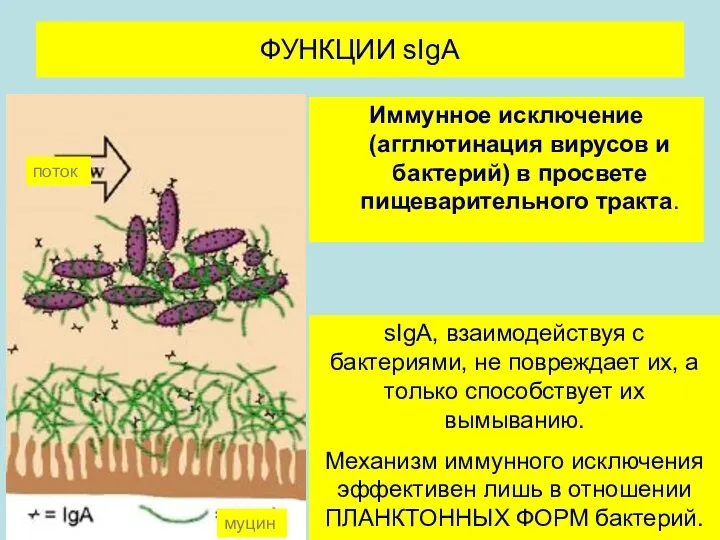
Иммунное исключение (агглютинация вирусов и бактерий) в просвете пищеварительного тракта.
ФУНКЦИИ
Иммунное исключение (агглютинация вирусов и бактерий) в просвете пищеварительного тракта.
ФУНКЦИИ

Иммуноглобулины класса А - IgA
Тяжелые цепи: α1 или α2 - альфа 1или
Иммуноглобулины класса А - IgA
Тяжелые цепи: α1 или α2 - альфа 1или

IgE - мономер
В сыворотке крови крайне мало - 0,002% от
IgE - мономер
В сыворотке крови крайне мало - 0,002% от
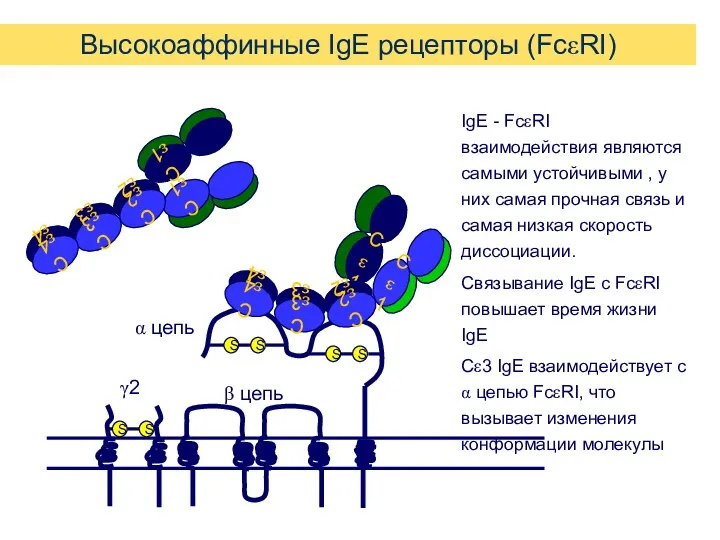
Высокоаффинные IgE рецепторы (FcεRI)
IgE - FcεRI взаимодействия являются самыми устойчивыми ,
Высокоаффинные IgE рецепторы (FcεRI)
IgE - FcεRI взаимодействия являются самыми устойчивыми ,
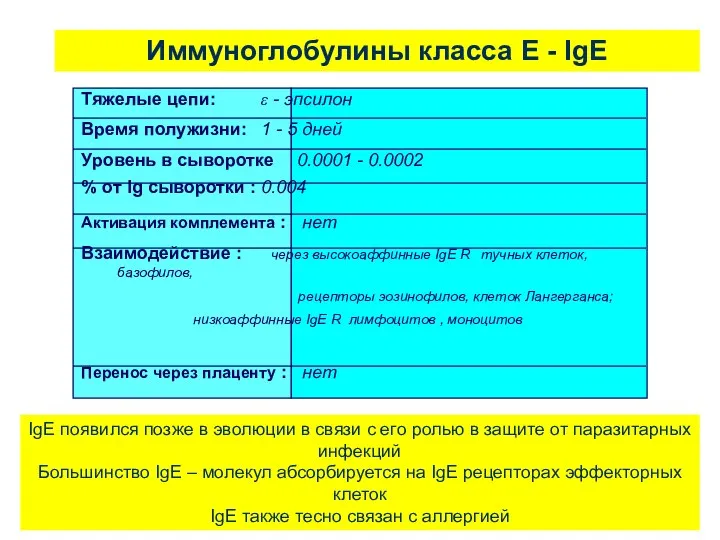
Иммуноглобулины класса Е - IgE
IgE появился позже в эволюции в связи
Иммуноглобулины класса Е - IgE
IgE появился позже в эволюции в связи

.
IgD - мономер
Является антигенспецифичным рецептором на поверхности В-лимфоцитов.
Биологическое
.
IgD - мономер
Является антигенспецифичным рецептором на поверхности В-лимфоцитов.
Биологическое

Иммуноглобулины класса D - IgD
IgD ко-экспрессируется вместе с IgM на В
Иммуноглобулины класса D - IgD
IgD ко-экспрессируется вместе с IgM на В

Различия в аминокислотной последовательности молекул антител
Изотипические различия между иммуноглобулиновыми молекулами:
Различия в аминокислотной последовательности молекул антител
Изотипические различия между иммуноглобулиновыми молекулами:

Роль антигенпрезентирующих клеток (АПК) в активации В лимфоцитов при ответе на
Роль антигенпрезентирующих клеток (АПК) в активации В лимфоцитов при ответе на

Взаимодействие Т и В лимфоцитов при ответе на Т- зависимые антигены
Взаимодействие Т и В лимфоцитов при ответе на Т- зависимые антигены

Взаимодействие Т и В лимфоцитов при ответе на Т- зависимые антигены
Комплекс
Взаимодействие Т и В лимфоцитов при ответе на Т- зависимые антигены
Комплекс
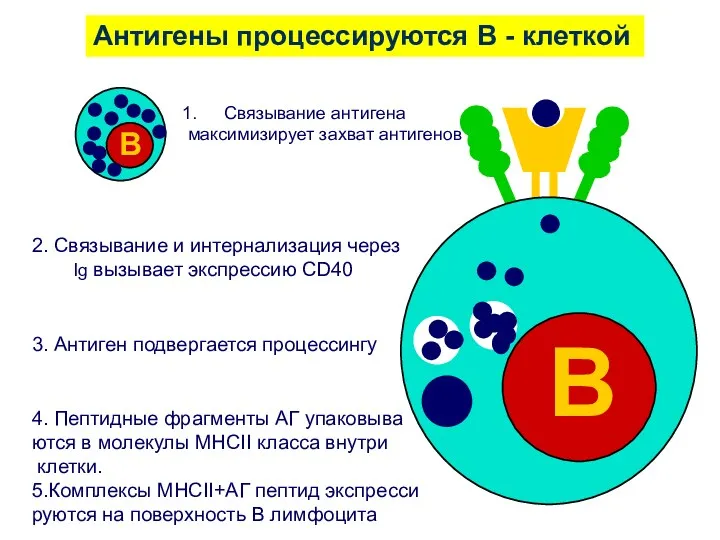
2. Связывание и интернализация через
Ig вызывает экспрессию CD40
3. Антиген
2. Связывание и интернализация через
Ig вызывает экспрессию CD40
3. Антиген
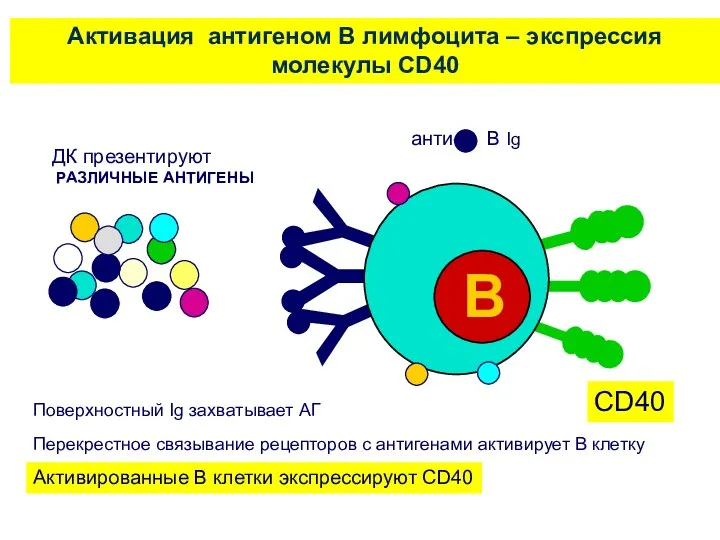
ДК презентируют
РАЗЛИЧНЫЕ АНТИГЕНЫ
Активация антигеном В лимфоцита – экспрессия молекулы CD40
Поверхностный
ДК презентируют
РАЗЛИЧНЫЕ АНТИГЕНЫ
Активация антигеном В лимфоцита – экспрессия молекулы CD40
Поверхностный
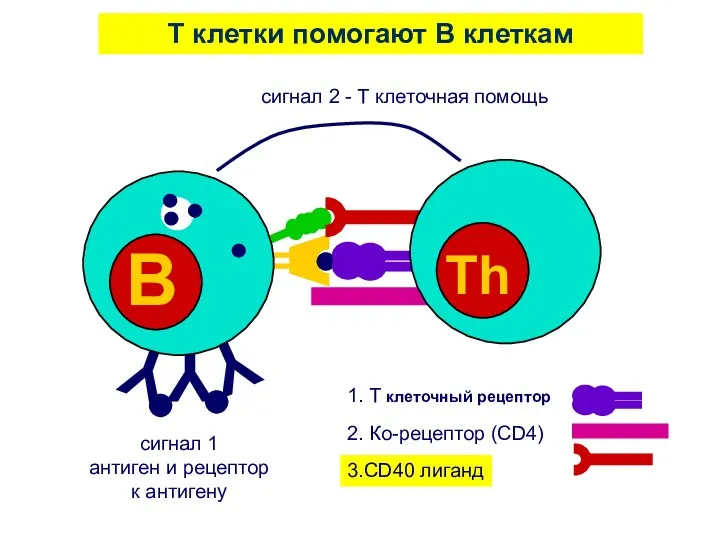
T клетки помогают B клеткам
сигнал 1
антиген и рецептор
к антигену
1. T
T клетки помогают B клеткам
сигнал 1
антиген и рецептор
к антигену
1. T
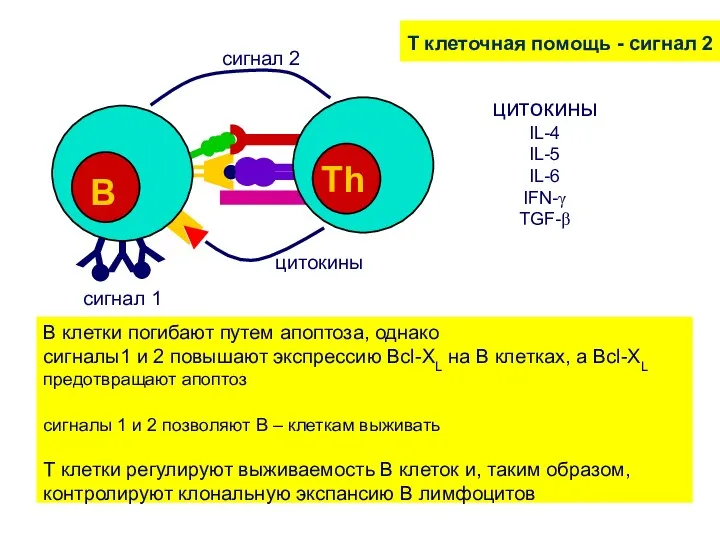
T клеточная помощь - сигнал 2
B клетки погибают путем апоптоза,
T клеточная помощь - сигнал 2
B клетки погибают путем апоптоза,

Цитокиновая активация В клеток
Активированные Т хелперы секретируют цитокины, активирующие В клетки.
IL-2
Цитокиновая активация В клеток
Активированные Т хелперы секретируют цитокины, активирующие В клетки.
IL-2

Ответ В клеток на Т-независимые антигены
При ответе на Т-независимые антигены В
Ответ В клеток на Т-независимые антигены
При ответе на Т-независимые антигены В
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
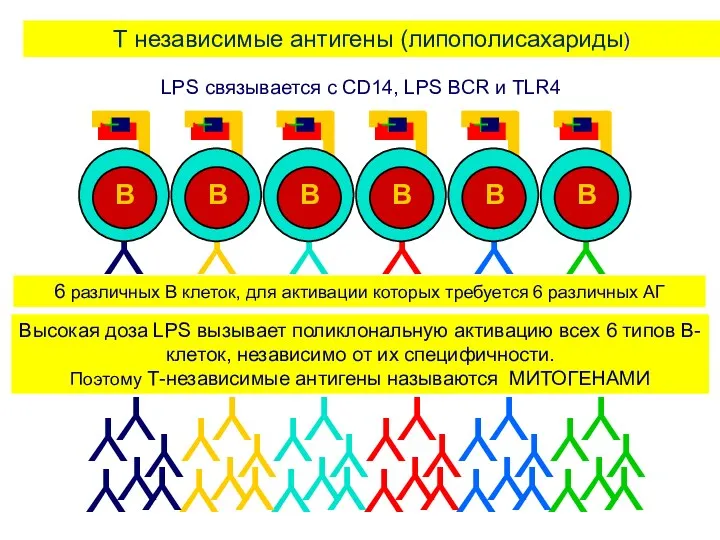
T независимые антигены (липополисахариды)
6 различных B клеток, для активации которых требуется
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
T независимые антигены (липополисахариды)
6 различных B клеток, для активации которых требуется

Ответ В клеток на Т- независимые антигены
В ответе В клеток на
Ответ В клеток на Т- независимые антигены
В ответе В клеток на
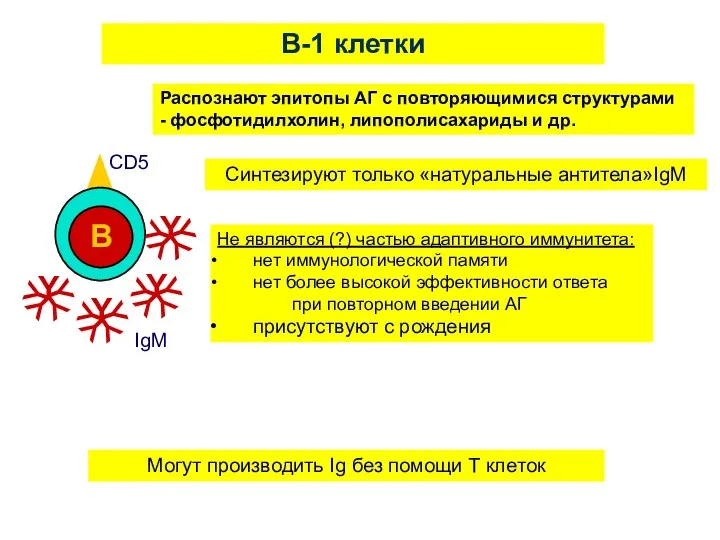
B-1 клетки
Распознают эпитопы АГ с повторяющимися структурами - фосфотидилхолин, липополисахариды и
B-1 клетки
Распознают эпитопы АГ с повторяющимися структурами - фосфотидилхолин, липополисахариды и

Эффекторные механизмы защиты против внеклеточных патогенов с помощью антител
Каков биологический смысл
Эффекторные механизмы защиты против внеклеточных патогенов с помощью антител
Каков биологический смысл
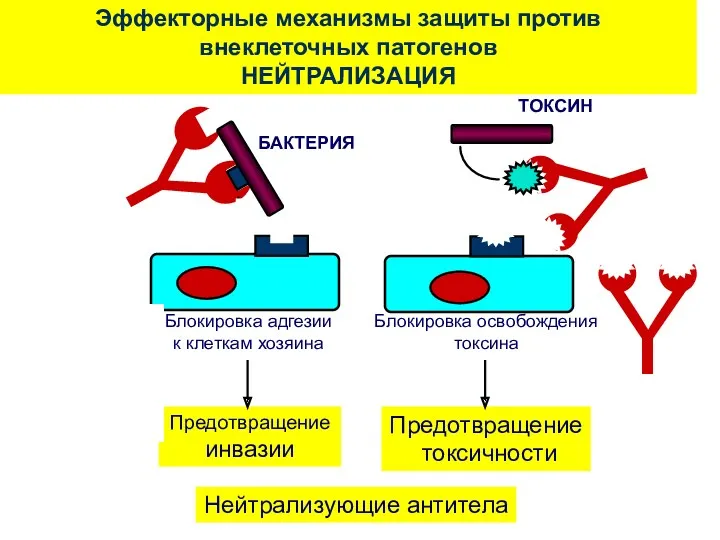
Эффекторные механизмы защиты против внеклеточных патогенов
НЕЙТРАЛИЗАЦИЯ
Нейтрализующие антитела
БАКТЕРИЯ
ТОКСИН
Эффекторные механизмы защиты против внеклеточных патогенов
НЕЙТРАЛИЗАЦИЯ
Нейтрализующие антитела
БАКТЕРИЯ
ТОКСИН
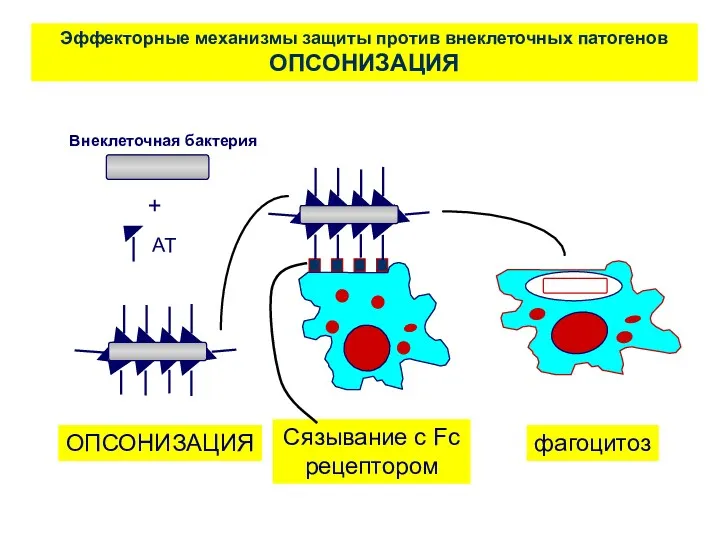
Эффекторные механизмы защиты против внеклеточных патогенов
ОПСОНИЗАЦИЯ
ОПСОНИЗАЦИЯ
Эффекторные механизмы защиты против внеклеточных патогенов
ОПСОНИЗАЦИЯ
ОПСОНИЗАЦИЯ
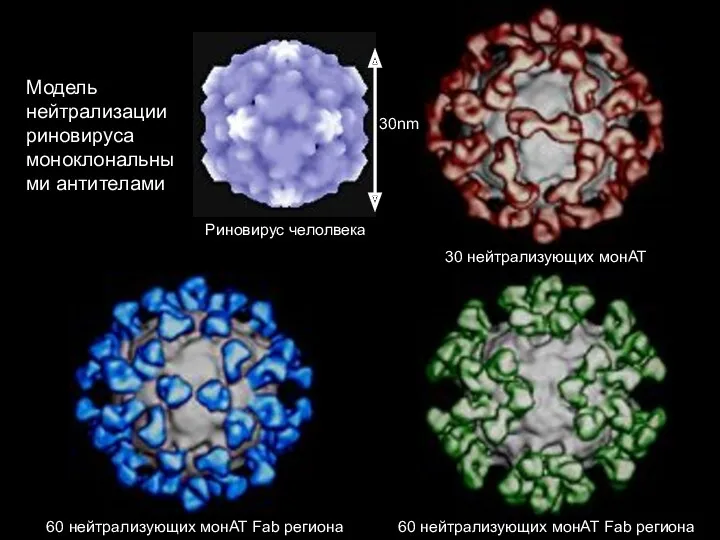
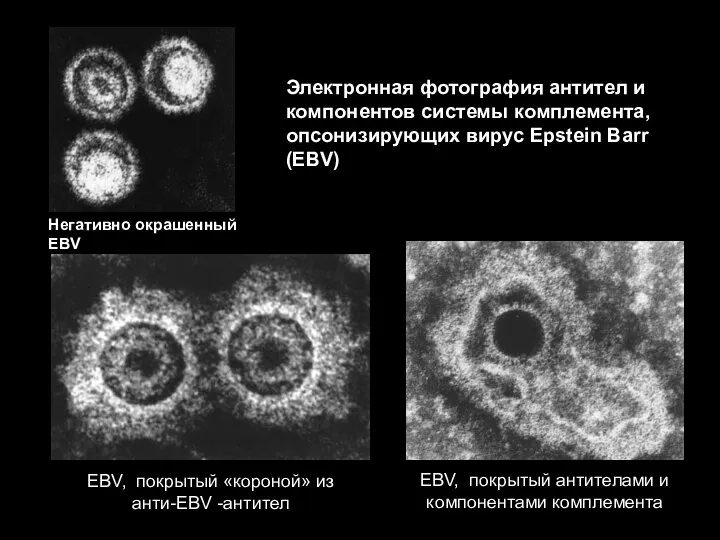
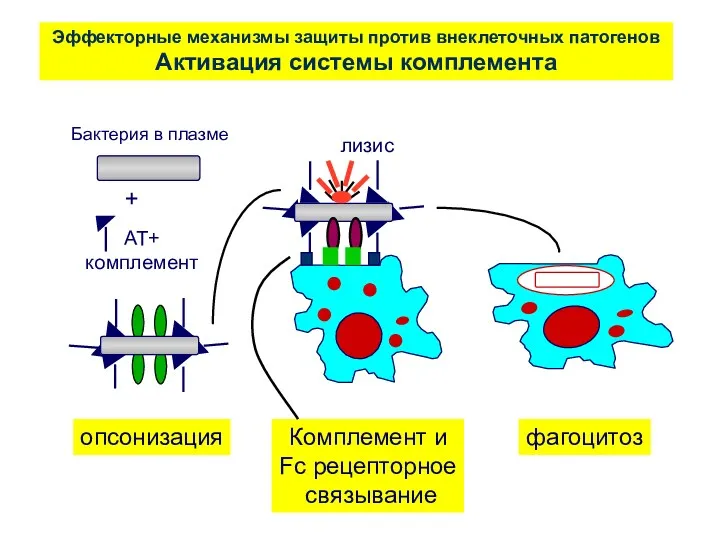
Эффекторные механизмы защиты против внеклеточных патогенов
Активация системы комплемента
лизис
Эффекторные механизмы защиты против внеклеточных патогенов
Активация системы комплемента
лизис

Вопросы
Фазы адаптивного иммунного ответа.
Типы иммунного ответа и факторы, их определяющие.
В-лимфоцит как
Вопросы
Фазы адаптивного иммунного ответа.
Типы иммунного ответа и факторы, их определяющие.
В-лимфоцит как

Тестовые вопросы
Антитела обладают способностью:
Расщеплять антиген
Активировать комплемент
Специфически связывать антиген
Фагоцитировать антиген
Экспрессировать антиген
Основным классом
Тестовые вопросы
Антитела обладают способностью:
Расщеплять антиген
Активировать комплемент
Специфически связывать антиген
Фагоцитировать антиген
Экспрессировать антиген
Основным классом

Тестовые вопросы
К «цитофильным» иммуноглобулинам относятся:
IgA
IgG
IgM
IgE
IgD
Основными иммуноглобулинами вторичного ответа являются:
IgA
IgG
IgM
IgE
IgD
Тестовые вопросы
К «цитофильным» иммуноглобулинам относятся:
IgA
IgG
IgM
IgE
IgD
Основными иммуноглобулинами вторичного ответа являются:
IgA
IgG
IgM
IgE
IgD

Тестовые вопросы
Активировать систему комплемента по классическому пути способны:
IgA
IgG
IgM
IgE
IgD
Изотипы иммуноглобулинов
Тестовые вопросы
Активировать систему комплемента по классическому пути способны:
IgA
IgG
IgM
IgE
IgD
Изотипы иммуноглобулинов

Тестовые вопросы
Через плаценту проникает:
IgA
IgG
IgM
IgE
IgD
Основными иммуноглобулинами первичного ответа являются:
IgA
IgG
IgM
IgD
IgE
Тестовые вопросы
Через плаценту проникает:
IgA
IgG
IgM
IgE
IgD
Основными иммуноглобулинами первичного ответа являются:
IgA
IgG
IgM
IgD
IgE
 Еркін қозғалыс жүйесі. Қозғалыс анализаторының анатомо-физиологиялық ерекшеліктері. Орталық және шеткі салдану белгілері
Еркін қозғалыс жүйесі. Қозғалыс анализаторының анатомо-физиологиялық ерекшеліктері. Орталық және шеткі салдану белгілері Принципы рационального питания. Лечебно-профилактическое питание, болезни связанные с характером питания
Принципы рационального питания. Лечебно-профилактическое питание, болезни связанные с характером питания Экстракорпоральные методы детоксикации
Экстракорпоральные методы детоксикации Балаларда Helicobacter pylori – инфекциясымен шақырылған асқорыту жолының жоғары бөлігін емдеу схемасы
Балаларда Helicobacter pylori – инфекциясымен шақырылған асқорыту жолының жоғары бөлігін емдеу схемасы Безсмертник
Безсмертник Онкология сегодня
Онкология сегодня Пороки развития головного мозга. Синдром Веста
Пороки развития головного мозга. Синдром Веста Дистрофии. Механизмы развития дистрофий
Дистрофии. Механизмы развития дистрофий Filling’s material: permanent & temporary
Filling’s material: permanent & temporary Гигиена зрения. Предупреждение глазных болезней
Гигиена зрения. Предупреждение глазных болезней Жүктіліктің УД зерттеу әдісі
Жүктіліктің УД зерттеу әдісі Сифилис. Лепра. Склерома. Сап. Карантинные инфекции
Сифилис. Лепра. Склерома. Сап. Карантинные инфекции Педикулез: симптомы, клиника, диагностика, лечение
Педикулез: симптомы, клиника, диагностика, лечение Описание локального статуса хирургического больного
Описание локального статуса хирургического больного Венерические заболевания
Венерические заболевания Первая медицинская помощь при неотложных состояниях
Первая медицинская помощь при неотложных состояниях Мукополисахаридоз типа I-Н (синдром Гурлер)
Мукополисахаридоз типа I-Н (синдром Гурлер) Организация акушерско-гинекологической помощи
Организация акушерско-гинекологической помощи Электромагниттердің адам өміріне зияны
Электромагниттердің адам өміріне зияны Транквилизаторы. Болеутоляющие средства. Седативные средства
Транквилизаторы. Болеутоляющие средства. Седативные средства Медикаментозная терапия при лихорадке
Медикаментозная терапия при лихорадке Понятия о ВИЧ-инфекции и СПИДе
Понятия о ВИЧ-инфекции и СПИДе Общественное здоровье и здравоохранение. Введение. (Лекция 1)
Общественное здоровье и здравоохранение. Введение. (Лекция 1) Семейства Гречишные (Polygonaceae), Буковые (Fagaceae), Берёзовые (Betulaceae)
Семейства Гречишные (Polygonaceae), Буковые (Fagaceae), Берёзовые (Betulaceae) Кардиопротективные свойства магния в комплексной терапии ишемической болезни сердца
Кардиопротективные свойства магния в комплексной терапии ишемической болезни сердца Государственное учреждение здравоохранения Севастополя Городская больница №5 – Центр охраны здоровья матери и ребенка
Государственное учреждение здравоохранения Севастополя Городская больница №5 – Центр охраны здоровья матери и ребенка Нарушение осанки и плоскостопие
Нарушение осанки и плоскостопие Дистоция плечиков
Дистоция плечиков