- Механизмы репарации ДНК

Содержание
- 2. Механизмы репарации ДНК
- 3. Объекты для изучения репарации Escherichia coli Saccharomyces cerevisiae Caenorhabditis elegans Arabidopsis Thaliana Мыши, крысы Клеточные культуры
- 4. 1. 1. Мисматч репарация (MMR) Этапы 1 - 3
- 5. MMR. Этапы 4 - 5
- 6. А. Репарация мисматчей у бактерий VSP- very short patch repair Short patch repair Long patch repair
- 7. VSP- very short patch repair – 1 В основном удаляется Т из мисматчей G/T и C/T.
- 8. VSP- very short patch repair – 2 MutY заменяет А из мисматчей C/A и G/A. Это
- 9. MMR млекопитающих 9 генов: MLH1, MLH3, PMS1-2, MSH2-6 MSH – гомолог MutS MLH – гомолог MutL
- 10. MMR человека На примере болезни HNPCC (heredity non-polyposis colorectal cancer) в 1993-1994 гг. У человека найдено
- 11. MMR человека На примере болезни HNPCC (heredity non-polyposis colorectal cancer) в 1993-1994 гг. Пациенты с HNPCC
- 12. Комбинация генов при репарации мисматчей
- 13. Показатели риска заболевания раком (Standardized incidence ratios - SIRs) на основании популяционных и клинических исследований дефекта
- 14. Механизмы, осуществляющие вклад в специфичность клеточных типов, чувствительных к дефициту MMR
- 15. 1. 2. UVR репарация
- 16. SOS-мутагенез у бактерий
- 17. 2. Прямая репарация Репарируются О6-метилгуанин и О4-метилгуанин ферментом МТаза (MGMT). У Е. coli 2 фермента (гены
- 18. Пример реакции
- 19. 3. BER-репарация. Этапы 1-2
- 20. BER-репарация. Этапы 3-4
- 21. BER-репарация. Этап 5
- 23. 4. NER-репарация 1. TCR – transcription coupled repair (преимущественная репарация траснкрибируемых цепей гена) 2. GGR –
- 24. Больные пигментной ксеродермой ( Выявлена в 1968 г. Дефект одного из 7 или более XP генов
- 25. Больные TTD трихотиодистрофией (А) и CS кокаиновым синдромом (В)
- 26. Этапы NER. 1-3
- 27. Этапы NER. 4-6
- 28. Биохимия NER (Этапы 1-3)
- 29. Специфическая активность ХР нуклеаз
- 30. Биохимия NER (Этапы 4-5)
- 31. Повреждение ХР при болезнях ХР – мутации в генах XP A-D,F,G TTD – серо-дефицитные хрупкие волосы,
- 32. Вклад NER генов в развитие сквамозной карциномы головы и шеи
- 33. Роль pol lI в репарации Когда pol II взаимодействует с промотором, она находится в гипофосфорилированном статусе
- 34. Rad26 создает NER комплекс в сайте повреждения. В то же время Rsp5 (и Ubc5, не показано)
- 35. Аддукты ДНК с цис-платином
- 36. Репарация аддуктов ДНК с цис-платином
- 37. 5. Другие виды репарации ДНК
- 38. Гомологичная репарация Продукты ГР
- 40. The double-strand break repair models through HR. Left panel: Gene conversion. After resection, the single-stranded 3′-tail
- 42. Белки ATM ATM (="ataxia telangiectasia mutated") получила название от болезни, пациенты, среди прочего, имеют высокий риск
- 43. Семейство АТМ белков
- 44. Активация АТМ
- 45. Примеры ICL, вызванных антираковыми агентами
- 46. Клеточный ответ на ICLs
- 47. Репарация ICL у млекопитающих
- 48. Fanconi Anemia (FA)путь репарации У пациентов с FA повреждено, по крайней мере, 13 генов: FANCA, B,
- 49. Сравнение FA генов у человека, Drosophila, Dictyostelium and C. elegans
- 50. FA путь у C. elegans.
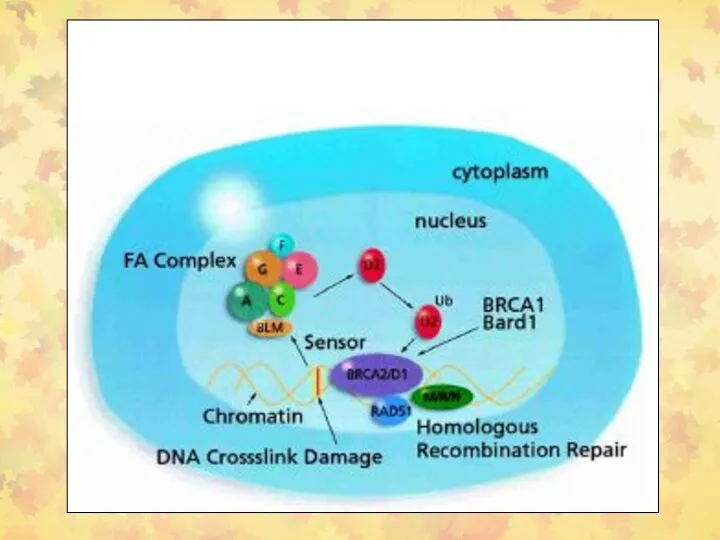
- 51. Fanconi Anemia путь регулирует репарацию ICLs ДНК с помощью гомологичной рекомбинации
- 53. BRCA1
- 54. Структура BRCA1
- 55. Ключевые шаги репарации DSB
- 56. Предполагаемая роль BRCA1 и BLM
- 57. Предполагаемая роль BRCA1 в остановке КЦ
- 58. Роль BRCA1 в репарации с гомологичной рекомбинацией
- 59. SUMO (small ubiquitin-related modifier) конъюгация Несколько SUMO E3 лигаз выявлено: SP-RING (secretory protein with a RING
- 60. Моделирование влияния SUMO конъюгации на BRCA1 Генотоксический стресс запускает SUMO модификации BRCA1 через активность UBC9–PIAS1 и
- 61. Репарация и рак
- 62. IY. Эпигенетические модификации ДНК Модификации хроматина, Метилирование ДНК, Геномный импринтинг.
- 63. Нуклеосомная организация ДНК
- 64. Регуляции транскрипции ацетилированием гистонов Гистон-деацетилаза (HDACs) деацетилирует лизиновые остатки, создавая предпосылки для метилирования HMT. ДНК может
- 65. Метилирование ДНК
- 67. Распределение метилирования
- 68. Статус метилирования и функциональные особенности промоторов, содержащих CpG-островки
- 70. Функции ДНК-МТаз
- 71. Активация транскрипции метилированием ДНК
- 73. Метилирование ДНК и рак
- 74. Морфологические изменения в хроматине (a) Нормальный эпителий кишечника: ядра разделены, одинаковы по форме и размеру (мономорфны).
- 75. Общие эпигенетические изменения при раке
- 76. Метилирование ДНК и рак
- 77. Эпигенетическая модель рака кишечника
- 78. Примеры гиперметилирования некоторых генов
- 79. Примеры гиперметилирования некоторых генов
- 81. Скачать презентацию

Механизмы репарации ДНК
Механизмы репарации ДНК

Объекты для изучения репарации
Escherichia coli
Saccharomyces cerevisiae
Caenorhabditis elegans
Arabidopsis Thaliana
Мыши, крысы
Клеточные культуры человека
Объекты для изучения репарации
Escherichia coli
Saccharomyces cerevisiae
Caenorhabditis elegans
Arabidopsis Thaliana
Мыши, крысы
Клеточные культуры человека
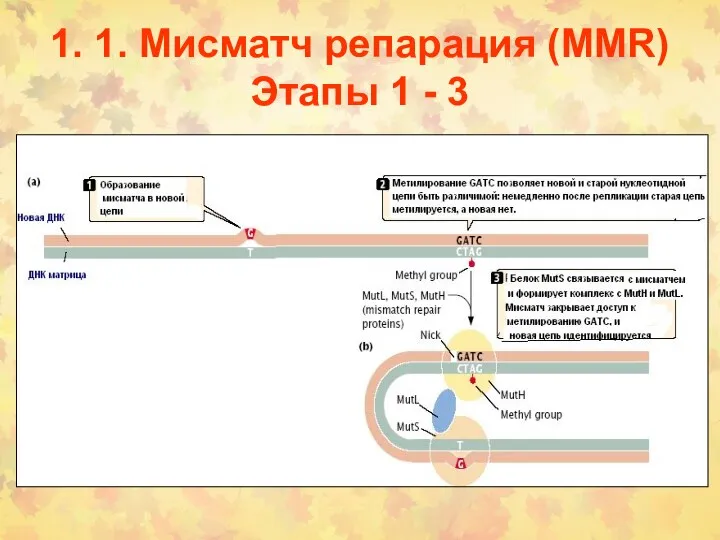
1. 1. Мисматч репарация (MMR) Этапы 1 - 3
1. 1. Мисматч репарация (MMR) Этапы 1 - 3
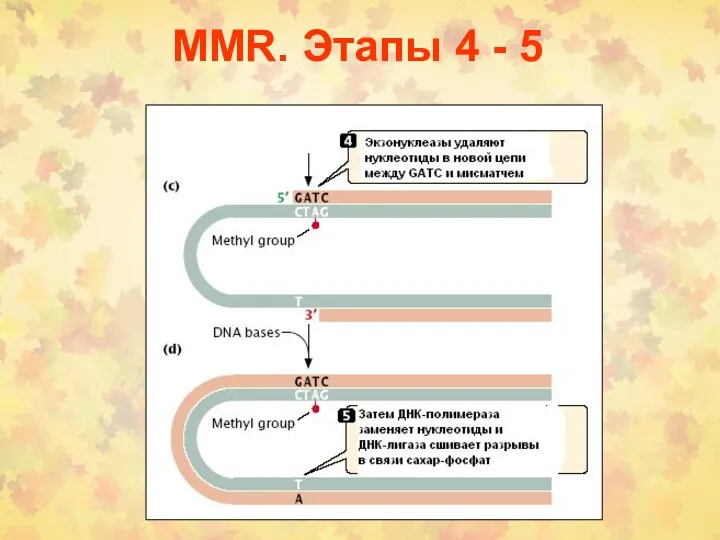
MMR. Этапы 4 - 5
MMR. Этапы 4 - 5

А. Репарация мисматчей
у бактерий
VSP- very short patch repair
Short patch repair
Long
А. Репарация мисматчей
у бактерий
VSP- very short patch repair
Short patch repair
Long

VSP- very short patch repair – 1
В основном удаляется Т из
VSP- very short patch repair – 1
В основном удаляется Т из

VSP- very short patch repair – 2
MutY заменяет А из мисматчей
VSP- very short patch repair – 2
MutY заменяет А из мисматчей

MMR млекопитающих
9 генов:
MLH1, MLH3, PMS1-2, MSH2-6
MSH – гомолог MutS
MLH – гомолог
MMR млекопитающих
9 генов:
MLH1, MLH3, PMS1-2, MSH2-6
MSH – гомолог MutS
MLH – гомолог

MMR человека
На примере болезни HNPCC (heredity
non-polyposis colorectal cancer) в 1993-1994
MMR человека
На примере болезни HNPCC (heredity
non-polyposis colorectal cancer) в 1993-1994

MMR человека
На примере болезни HNPCC (heredity non-polyposis colorectal cancer) в 1993-1994
MMR человека
На примере болезни HNPCC (heredity non-polyposis colorectal cancer) в 1993-1994
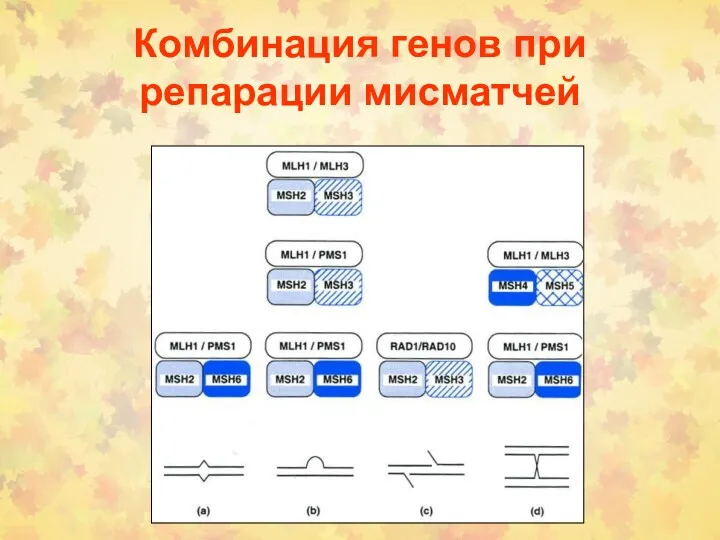
Комбинация генов при репарации мисматчей
Комбинация генов при репарации мисматчей
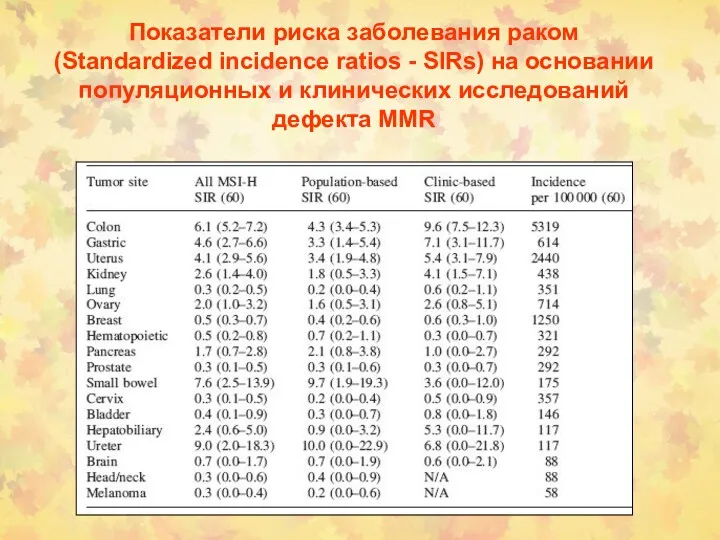
Показатели риска заболевания раком
(Standardized incidence ratios - SIRs) на основании
Показатели риска заболевания раком (Standardized incidence ratios - SIRs) на основании

Механизмы, осуществляющие вклад в специфичность клеточных типов, чувствительных к дефициту MMR
Механизмы, осуществляющие вклад в специфичность клеточных типов, чувствительных к дефициту MMR
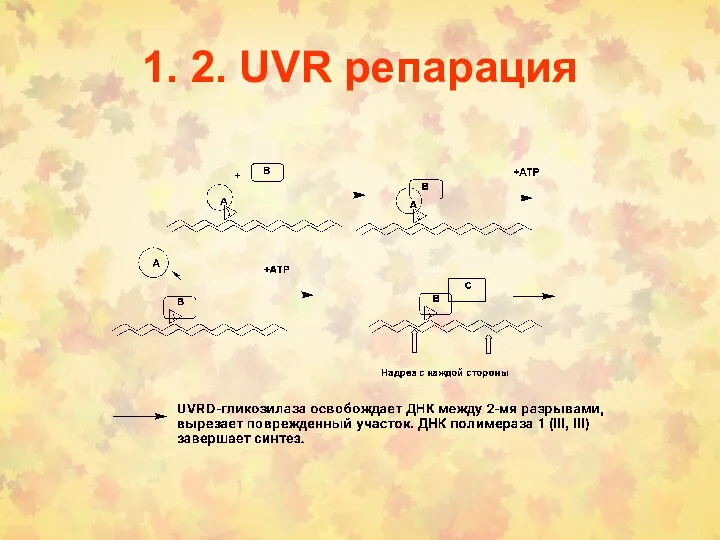
1. 2. UVR репарация
1. 2. UVR репарация
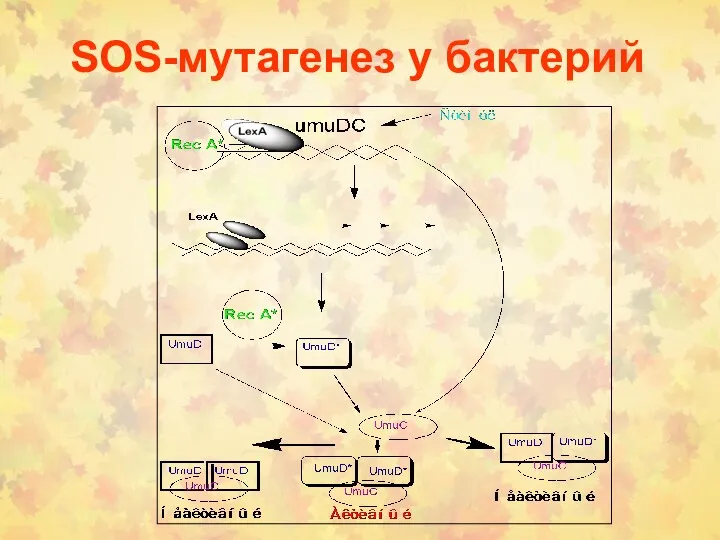
SOS-мутагенез у бактерий
SOS-мутагенез у бактерий

2. Прямая репарация
Репарируются О6-метилгуанин и О4-метилгуанин ферментом МТаза (MGMT).
У Е. coli
2. Прямая репарация
Репарируются О6-метилгуанин и О4-метилгуанин ферментом МТаза (MGMT).
У Е. coli
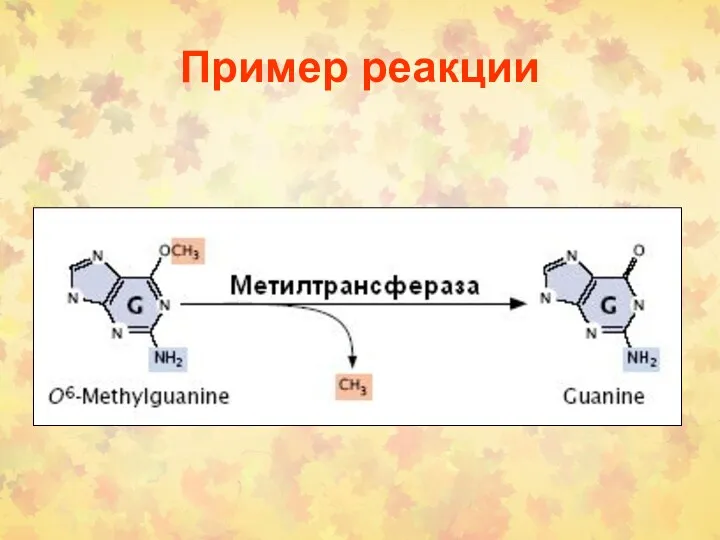
Пример реакции
Пример реакции
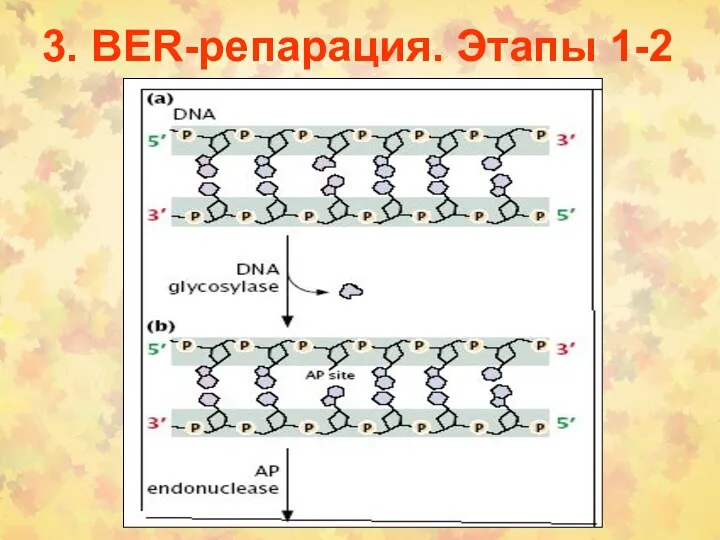
3. BER-репарация. Этапы 1-2
3. BER-репарация. Этапы 1-2
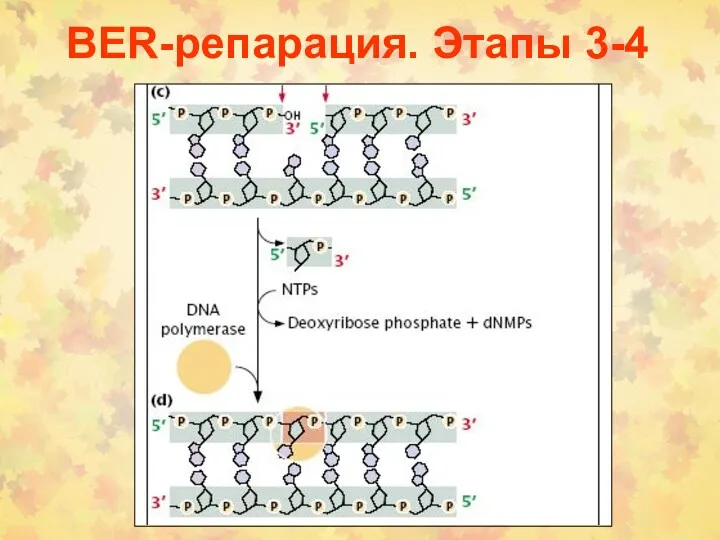
BER-репарация. Этапы 3-4
BER-репарация. Этапы 3-4
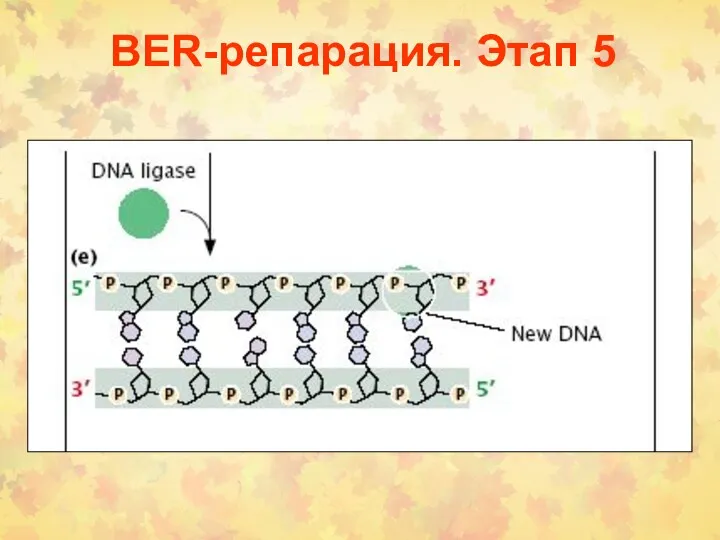
BER-репарация. Этап 5
BER-репарация. Этап 5
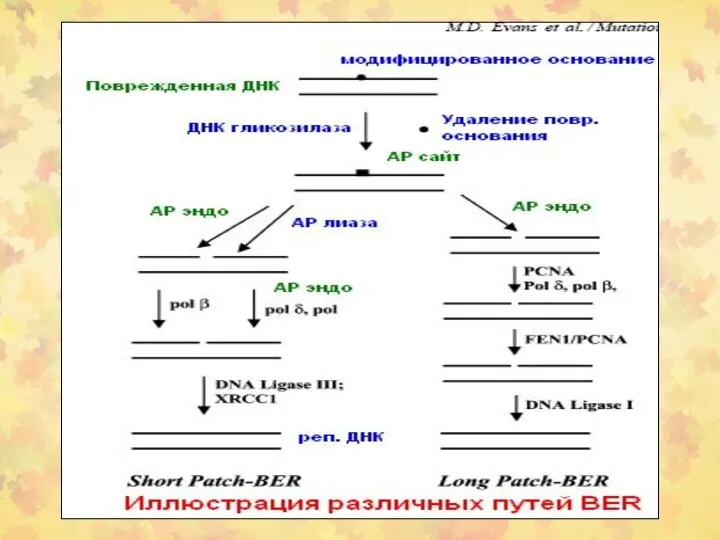

4. NER-репарация
1. TCR – transcription coupled repair (преимущественная репарация траснкрибируемых цепей
4. NER-репарация
1. TCR – transcription coupled repair (преимущественная репарация траснкрибируемых цепей
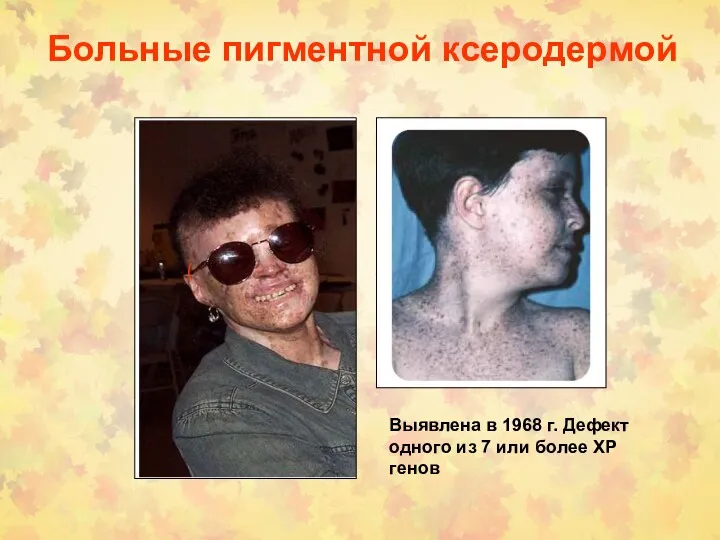
Больные пигментной ксеродермой
(
Выявлена в 1968 г. Дефект одного из 7 или
Больные пигментной ксеродермой
(
Выявлена в 1968 г. Дефект одного из 7 или
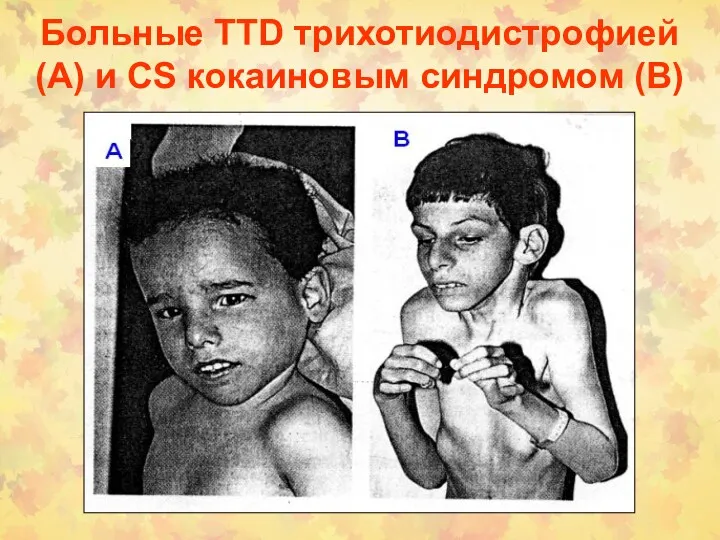
Больные TTD трихотиодистрофией (А) и CS кокаиновым синдромом (В)
Больные TTD трихотиодистрофией (А) и CS кокаиновым синдромом (В)
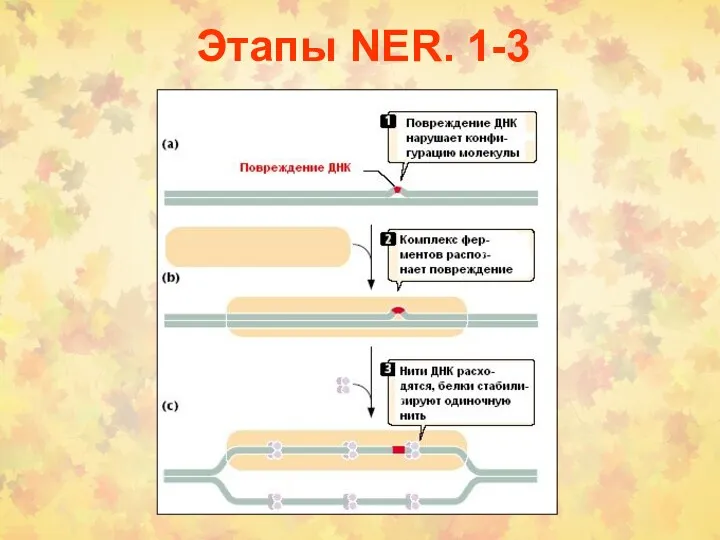
Этапы NER. 1-3
Этапы NER. 1-3
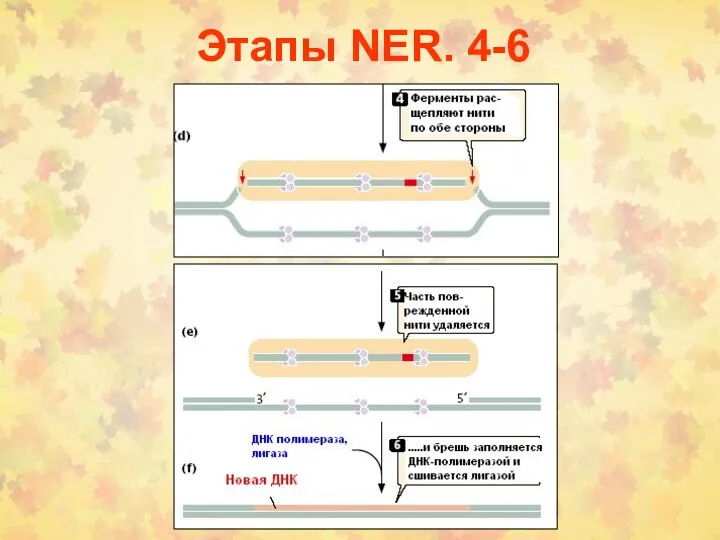
Этапы NER. 4-6
Этапы NER. 4-6
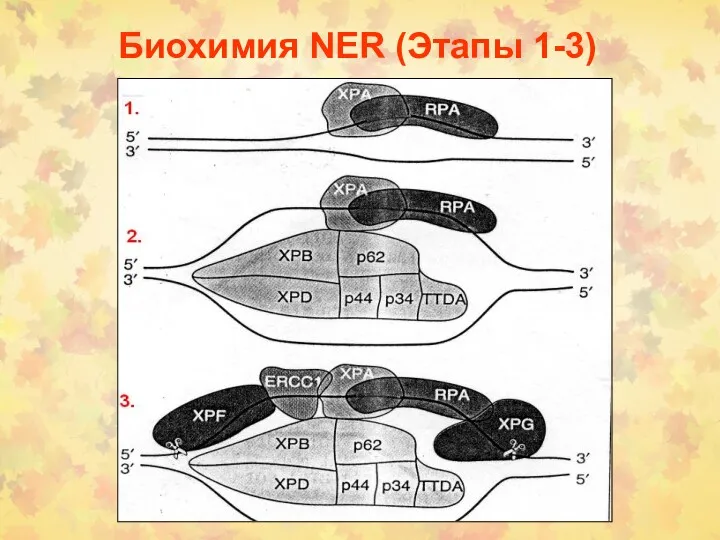
Биохимия NER (Этапы 1-3)
Биохимия NER (Этапы 1-3)
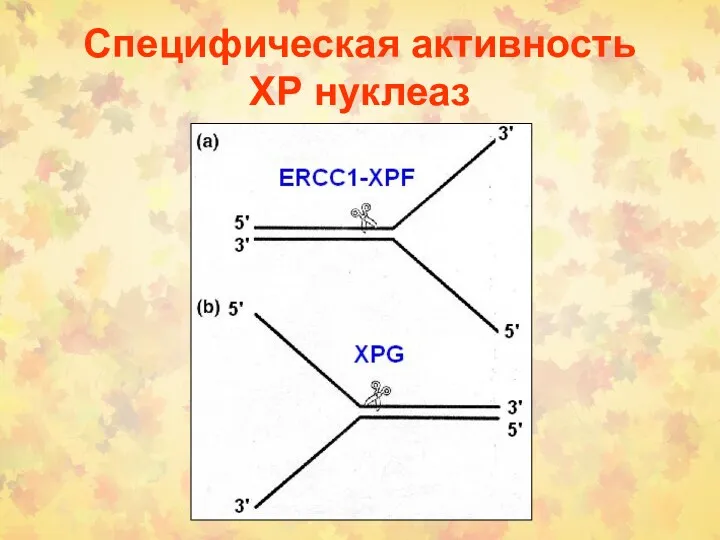
Специфическая активность
ХР нуклеаз
Специфическая активность
ХР нуклеаз

Биохимия NER (Этапы 4-5)
Биохимия NER (Этапы 4-5)

Повреждение ХР при болезнях
ХР – мутации в генах XP A-D,F,G
TTD –
Повреждение ХР при болезнях
ХР – мутации в генах XP A-D,F,G
TTD –
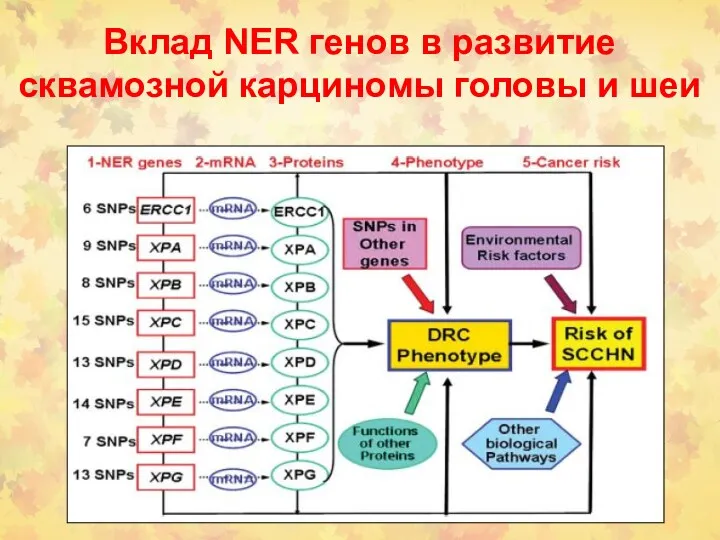
Вклад NER генов в развитие сквамозной карциномы головы и шеи
Вклад NER генов в развитие сквамозной карциномы головы и шеи
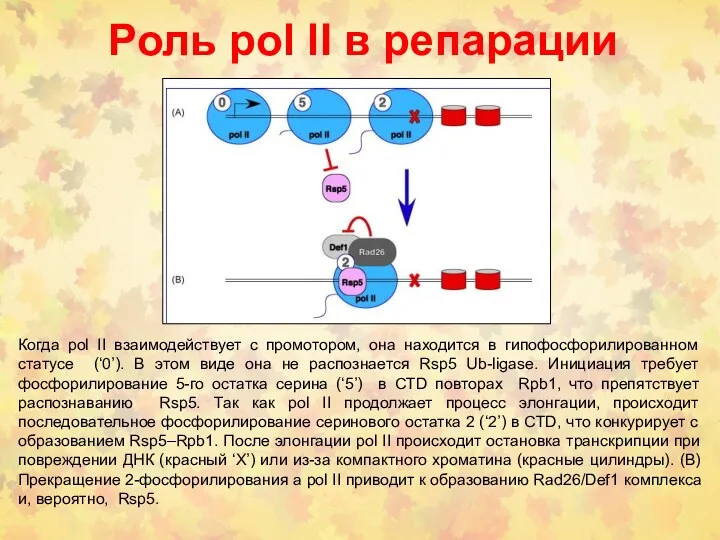
Роль pol lI в репарации
Когда pol II взаимодействует с промотором, она
Роль pol lI в репарации
Когда pol II взаимодействует с промотором, она
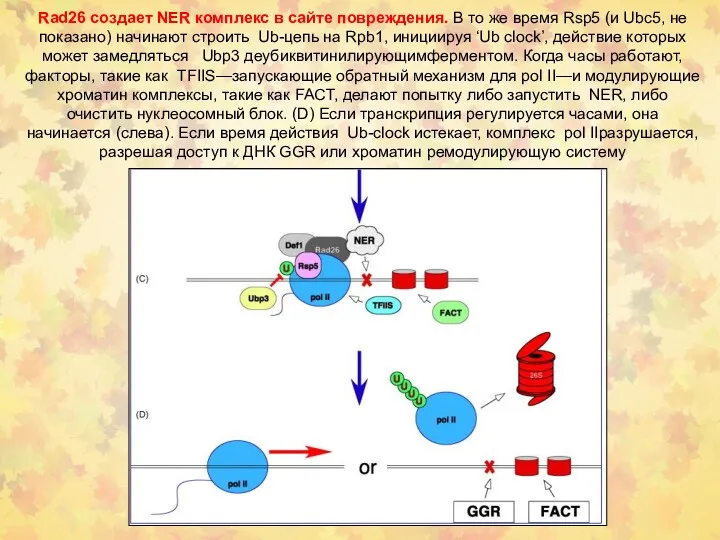
Rad26 создает NER комплекс в сайте повреждения. В то же время
Rad26 создает NER комплекс в сайте повреждения. В то же время
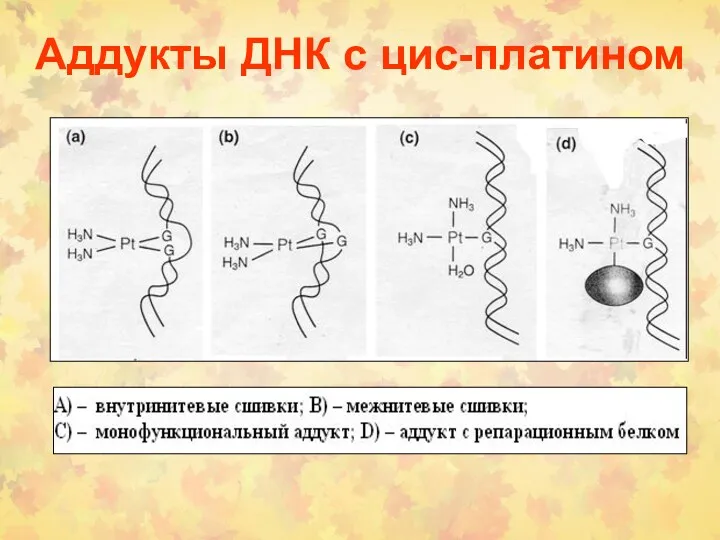
Аддукты ДНК с цис-платином
Аддукты ДНК с цис-платином
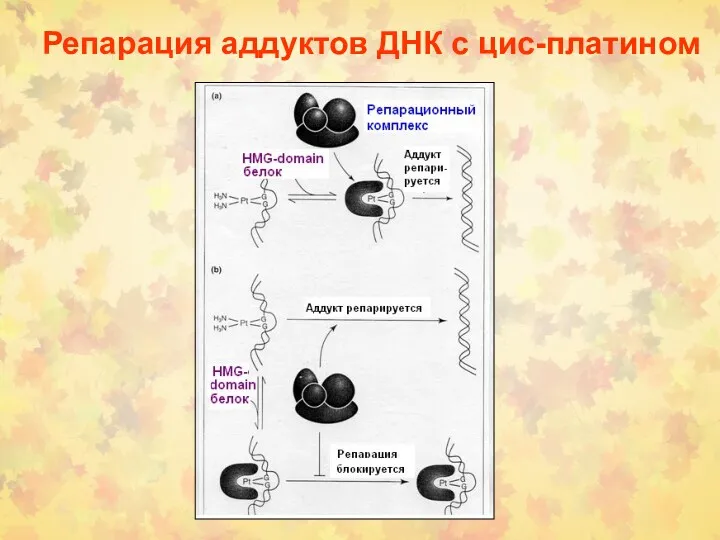
Репарация аддуктов ДНК с цис-платином
Репарация аддуктов ДНК с цис-платином
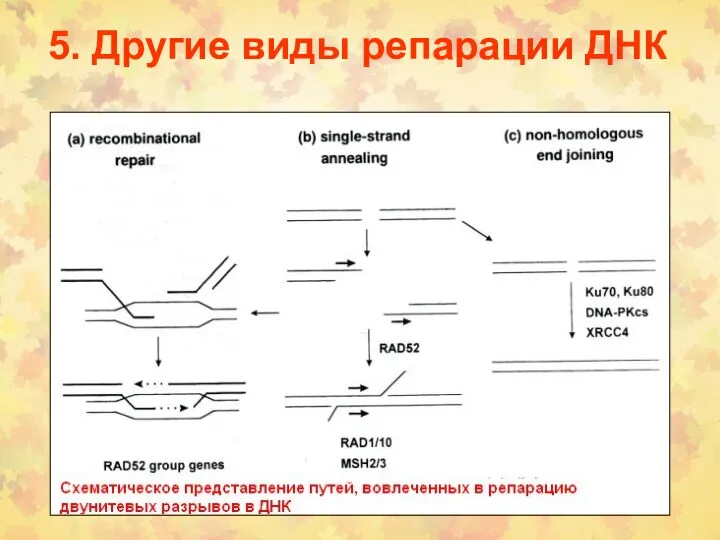
5. Другие виды репарации ДНК
5. Другие виды репарации ДНК
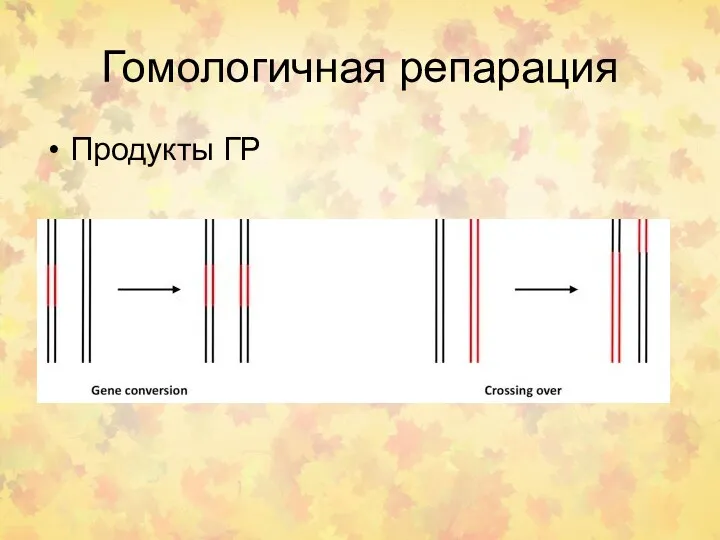
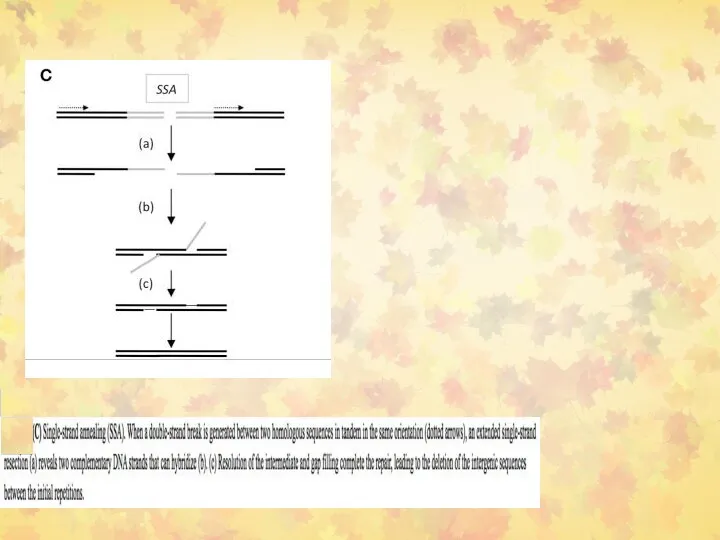
Гомологичная репарация
Продукты ГР
Гомологичная репарация
Продукты ГР
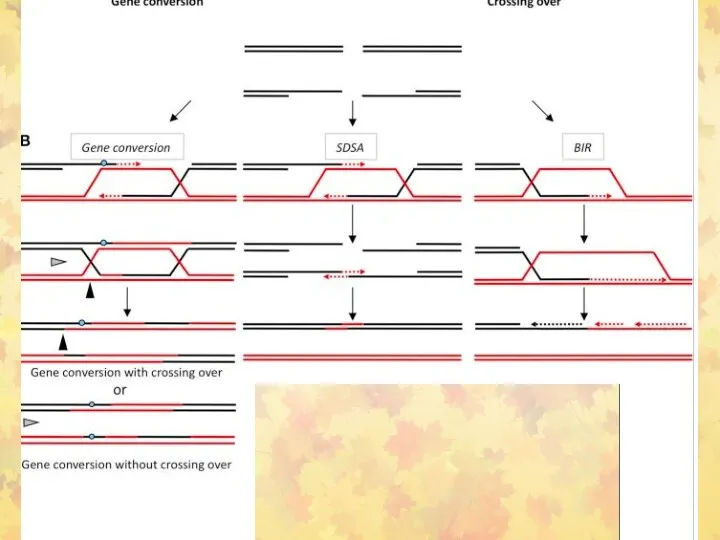

The double-strand break repair models through HR. Left panel: Gene conversion.
The double-strand break repair models through HR. Left panel: Gene conversion.

Белки ATM
ATM (="ataxia telangiectasia mutated") получила название от болезни, пациенты,
Белки ATM
ATM (="ataxia telangiectasia mutated") получила название от болезни, пациенты,

Семейство АТМ белков
Семейство АТМ белков
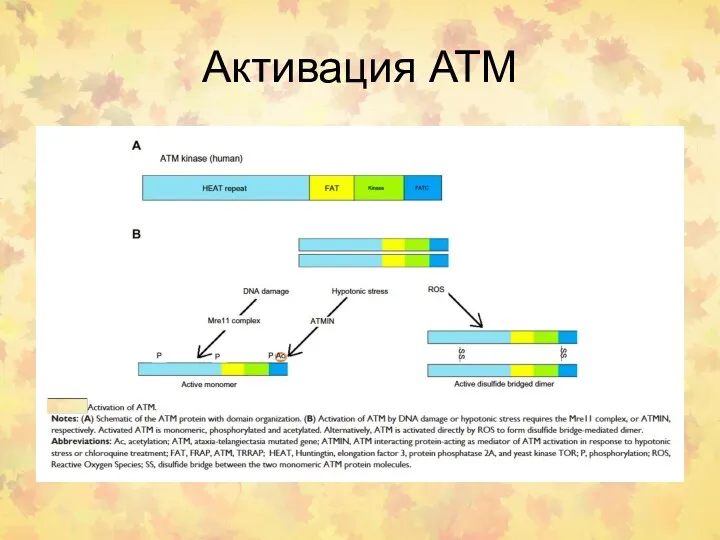
Активация АТМ
Активация АТМ
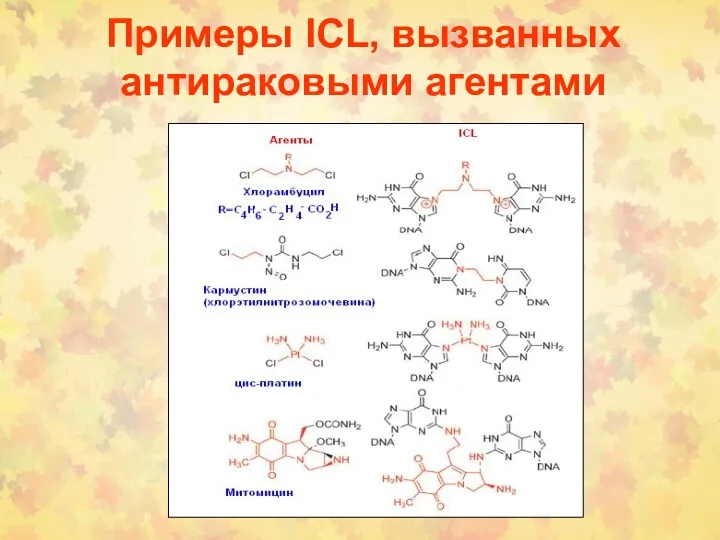
Примеры ICL, вызванных антираковыми агентами
Примеры ICL, вызванных антираковыми агентами
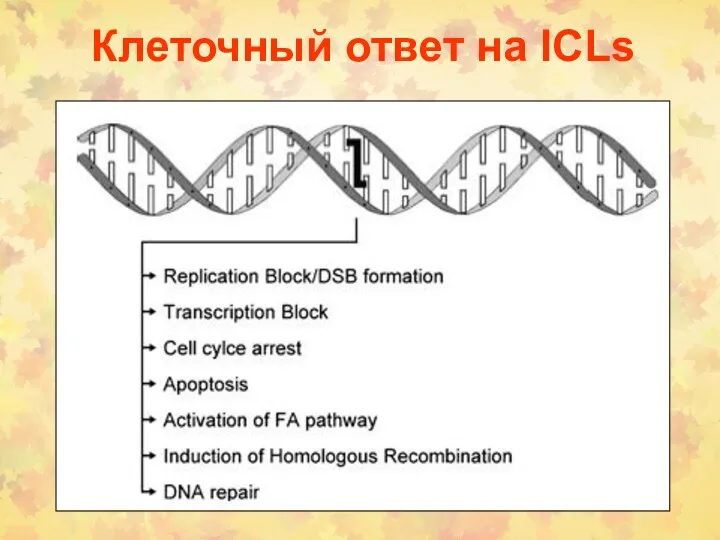
Клеточный ответ на ICLs
Клеточный ответ на ICLs
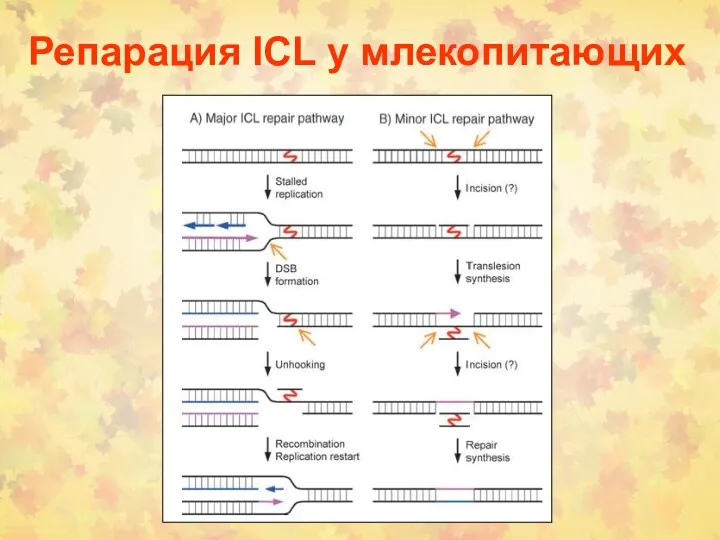
Репарация ICL у млекопитающих
Репарация ICL у млекопитающих

Fanconi Anemia (FA)путь репарации
У пациентов с FA повреждено, по крайней мере,
Fanconi Anemia (FA)путь репарации
У пациентов с FA повреждено, по крайней мере,

Сравнение FA генов у человека, Drosophila, Dictyostelium and C. elegans
Сравнение FA генов у человека, Drosophila, Dictyostelium and C. elegans
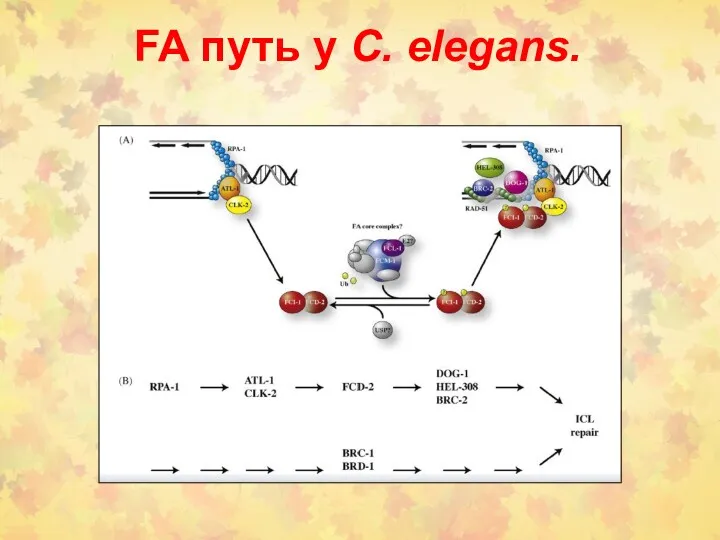
FA путь у C. elegans.
FA путь у C. elegans.
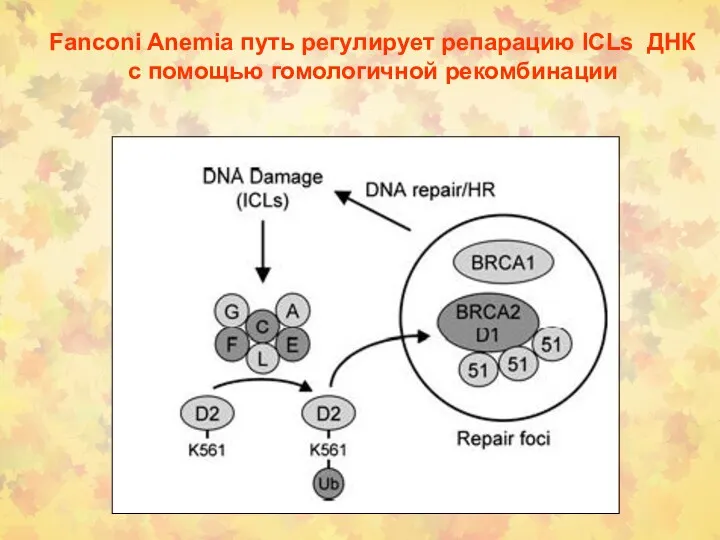
Fanconi Anemia путь регулирует репарацию ICLs ДНК с помощью гомологичной рекомбинации
Fanconi Anemia путь регулирует репарацию ICLs ДНК с помощью гомологичной рекомбинации
BRCA1
BRCA1
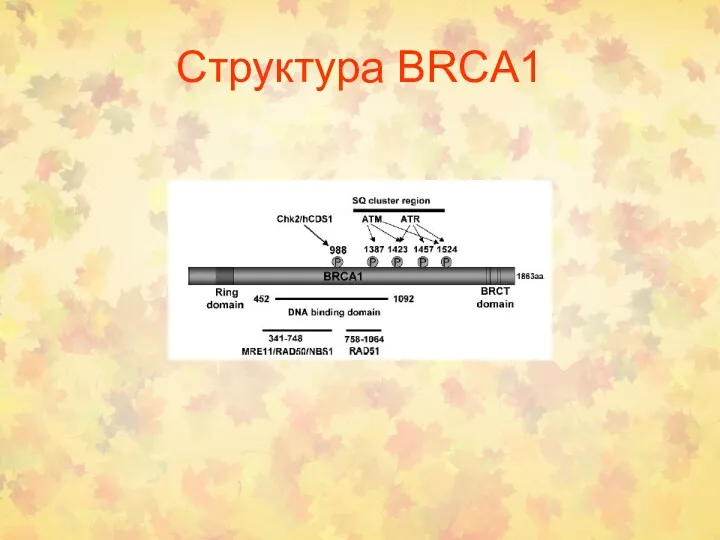
Структура BRCA1
Структура BRCA1
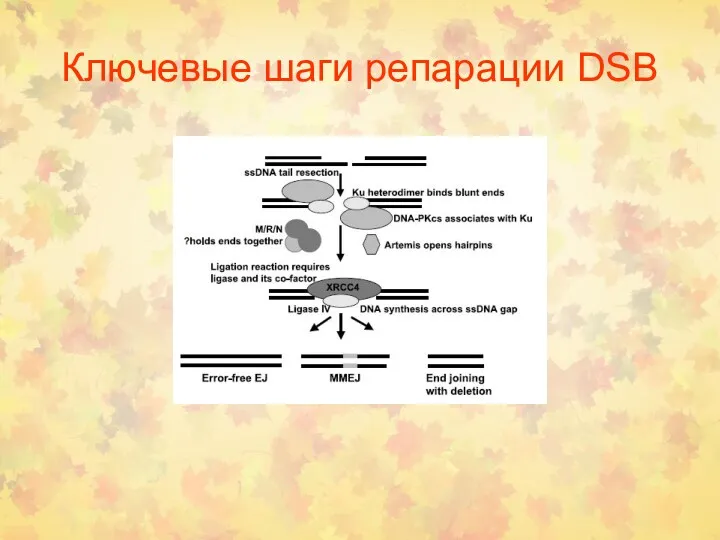
Ключевые шаги репарации DSB
Ключевые шаги репарации DSB
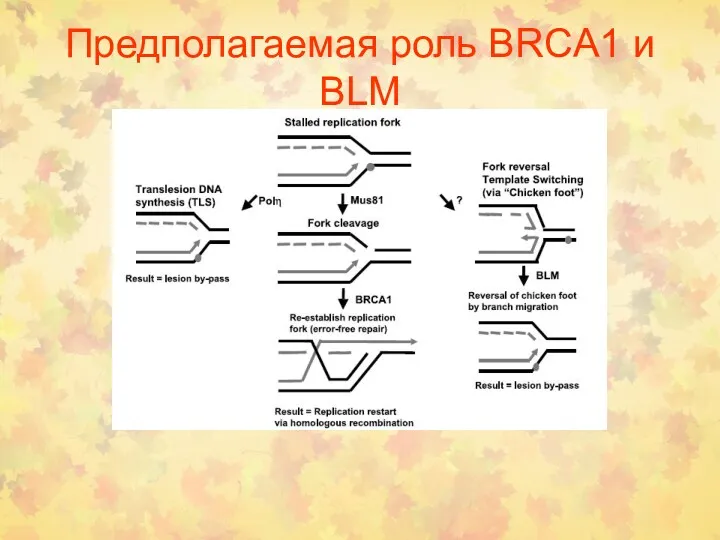
Предполагаемая роль BRCA1 и BLM
Предполагаемая роль BRCA1 и BLM
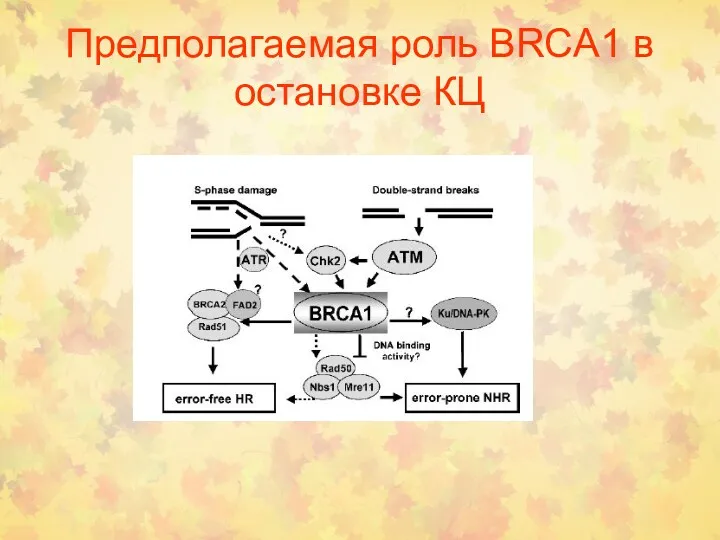
Предполагаемая роль BRCA1 в остановке КЦ
Предполагаемая роль BRCA1 в остановке КЦ
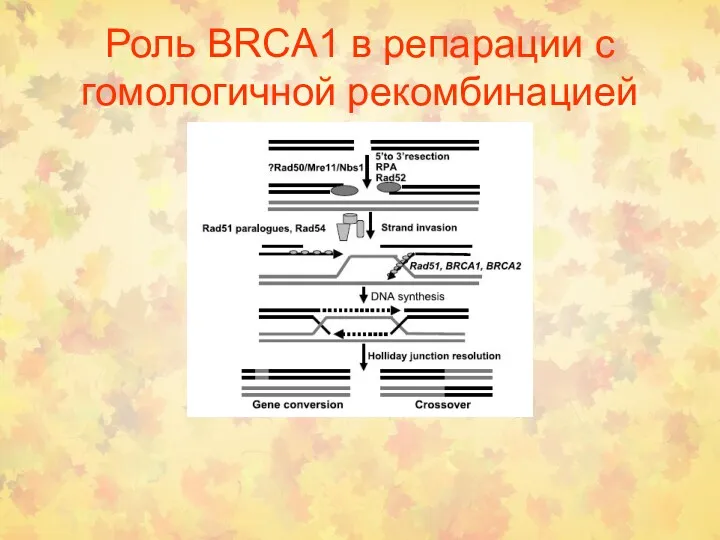
Роль BRCA1 в репарации с гомологичной рекомбинацией
Роль BRCA1 в репарации с гомологичной рекомбинацией

SUMO
(small ubiquitin-related modifier) конъюгация
Несколько SUMO E3 лигаз выявлено: SP-RING (secretory
SUMO
(small ubiquitin-related modifier) конъюгация
Несколько SUMO E3 лигаз выявлено: SP-RING (secretory
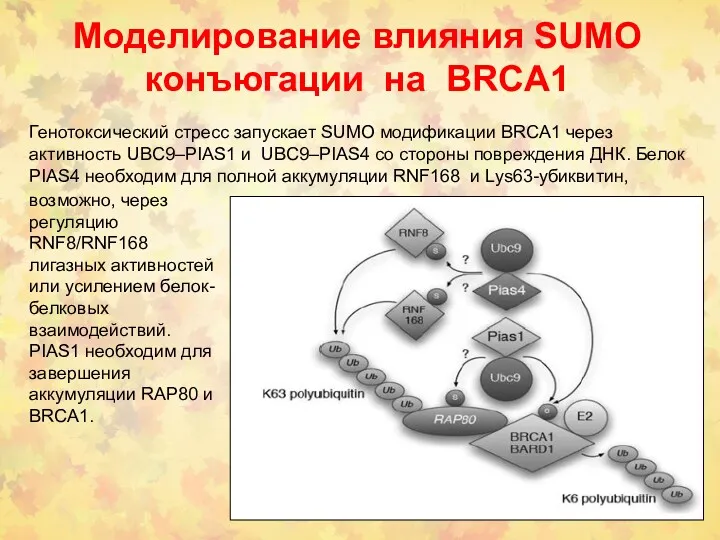
Моделирование влияния SUMO конъюгации на BRCA1
Генотоксический стресс запускает SUMO модификации BRCA1
Моделирование влияния SUMO конъюгации на BRCA1
Генотоксический стресс запускает SUMO модификации BRCA1

Репарация и рак
Репарация и рак

IY. Эпигенетические модификации ДНК
Модификации хроматина,
Метилирование ДНК,
Геномный импринтинг.
IY. Эпигенетические модификации ДНК
Модификации хроматина,
Метилирование ДНК,
Геномный импринтинг.
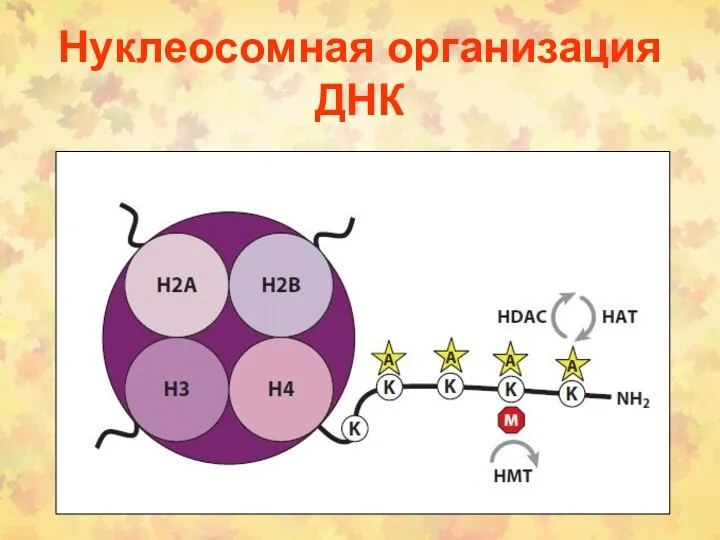
Нуклеосомная организация ДНК
Нуклеосомная организация ДНК
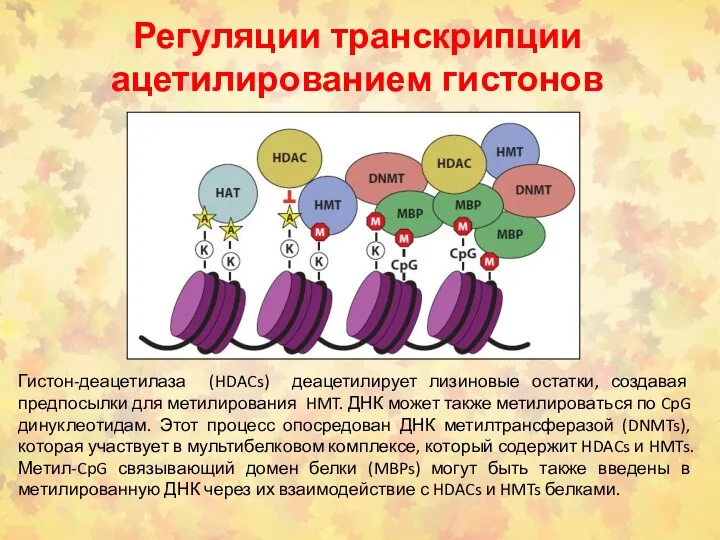
Регуляции транскрипции ацетилированием гистонов
Гистон-деацетилаза (HDACs) деацетилирует лизиновые остатки, создавая предпосылки для
Регуляции транскрипции ацетилированием гистонов
Гистон-деацетилаза (HDACs) деацетилирует лизиновые остатки, создавая предпосылки для
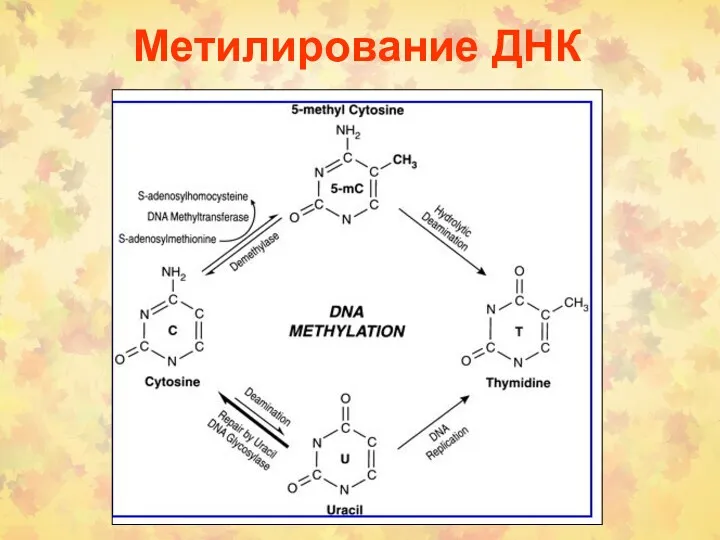
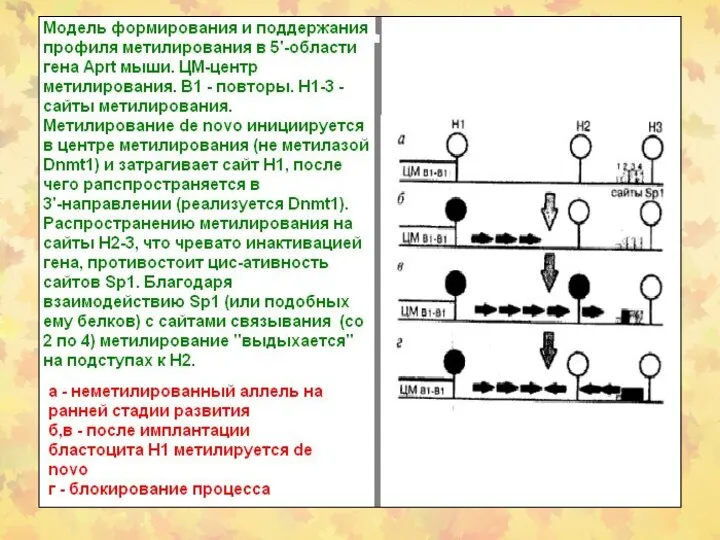
Метилирование ДНК
Метилирование ДНК
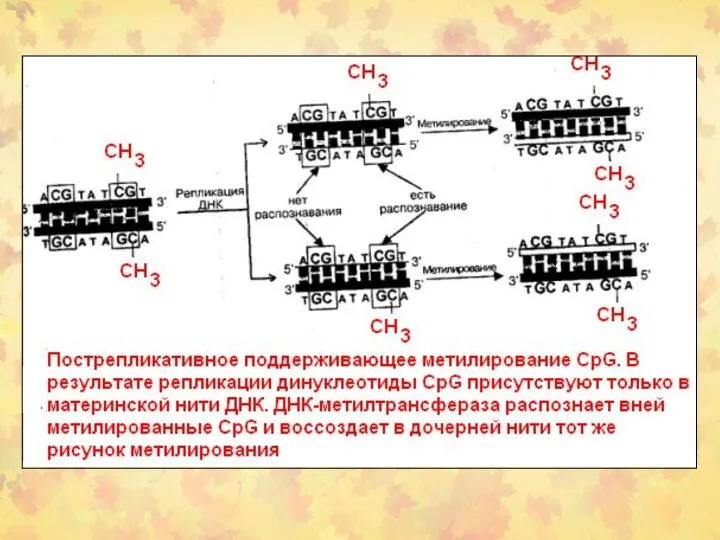
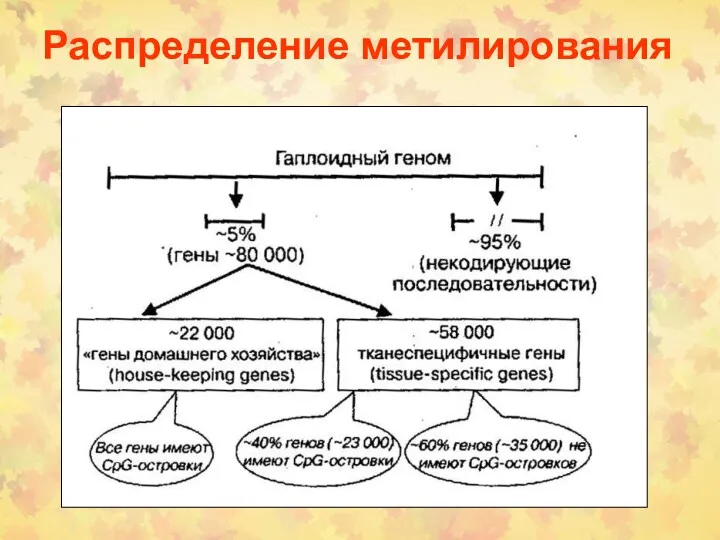
Распределение метилирования
Распределение метилирования

Статус метилирования и функциональные особенности промоторов, содержащих CpG-островки
Статус метилирования и функциональные особенности промоторов, содержащих CpG-островки
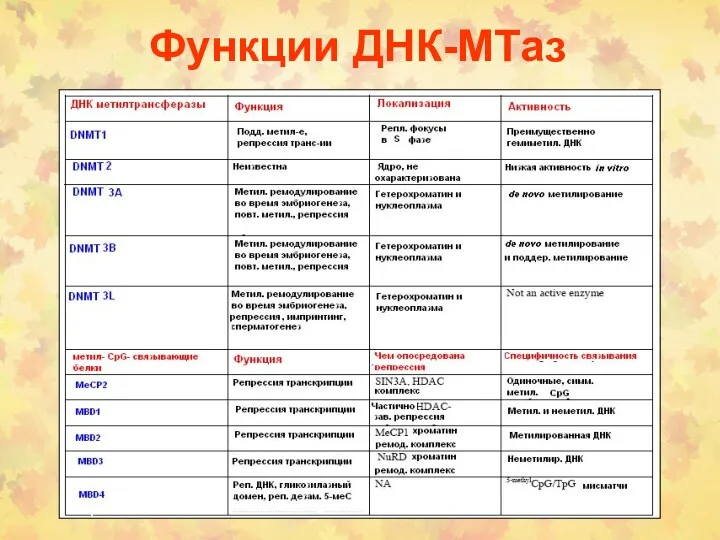
Функции ДНК-МТаз
Функции ДНК-МТаз
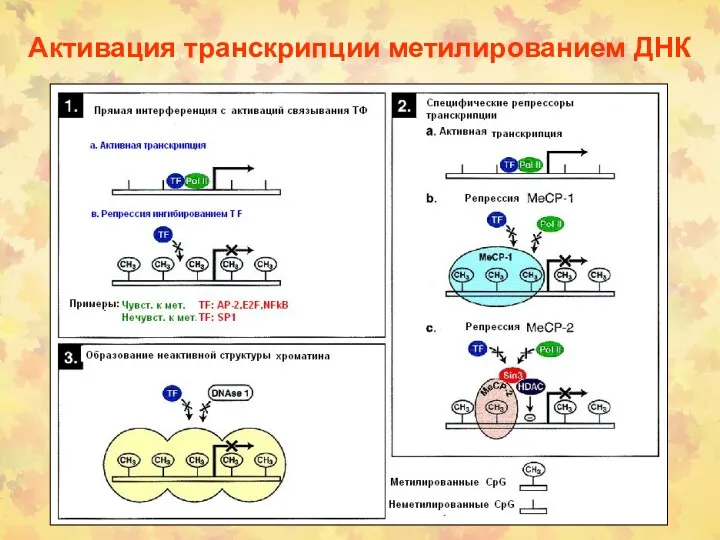
Активация транскрипции метилированием ДНК
Активация транскрипции метилированием ДНК

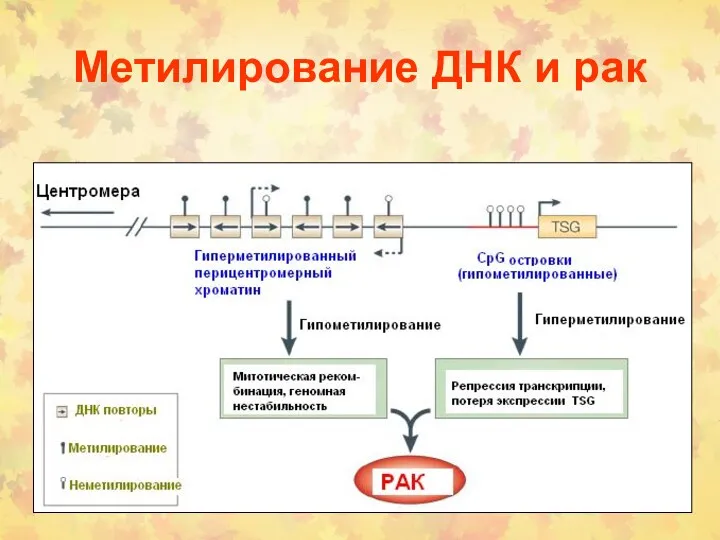
Метилирование ДНК и рак
Метилирование ДНК и рак
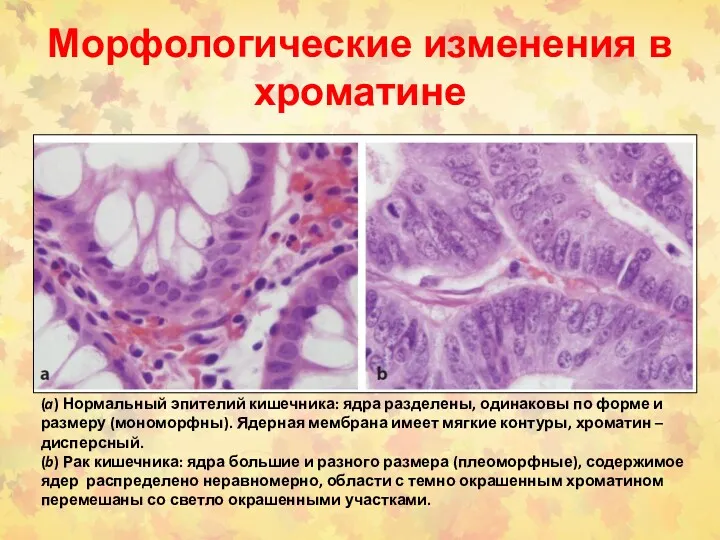
Морфологические изменения в хроматине
(a) Нормальный эпителий кишечника: ядра разделены, одинаковы по
Морфологические изменения в хроматине
(a) Нормальный эпителий кишечника: ядра разделены, одинаковы по
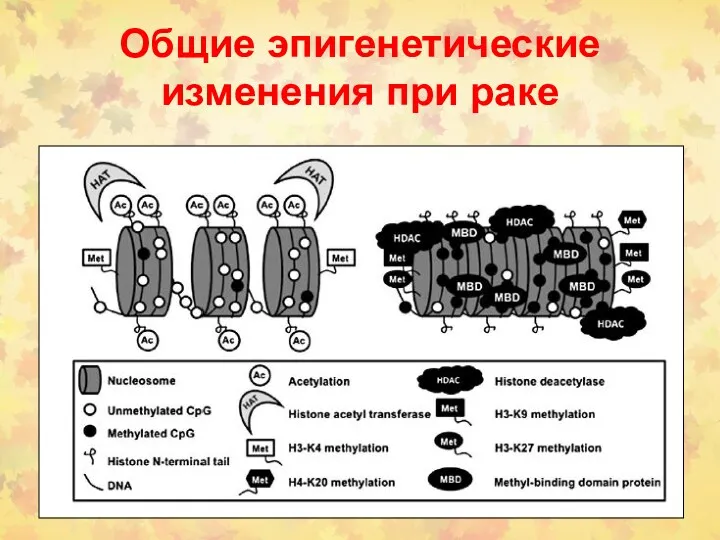
Общие эпигенетические изменения при раке
Общие эпигенетические изменения при раке
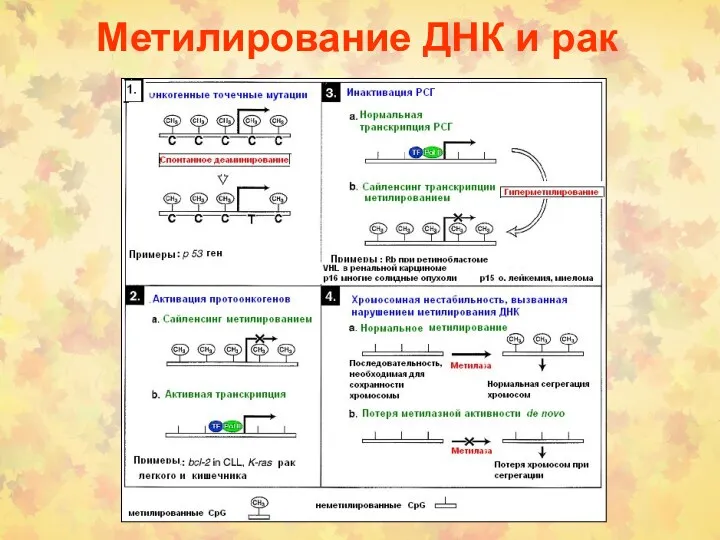
Метилирование ДНК и рак
Метилирование ДНК и рак
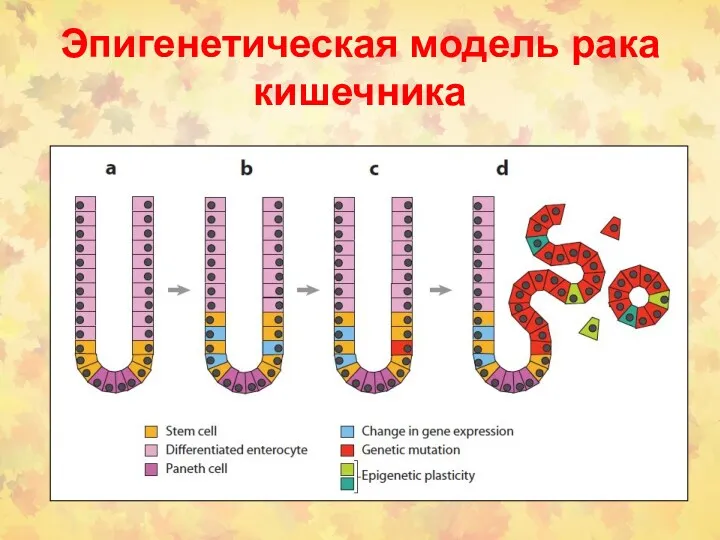
Эпигенетическая модель рака кишечника
Эпигенетическая модель рака кишечника
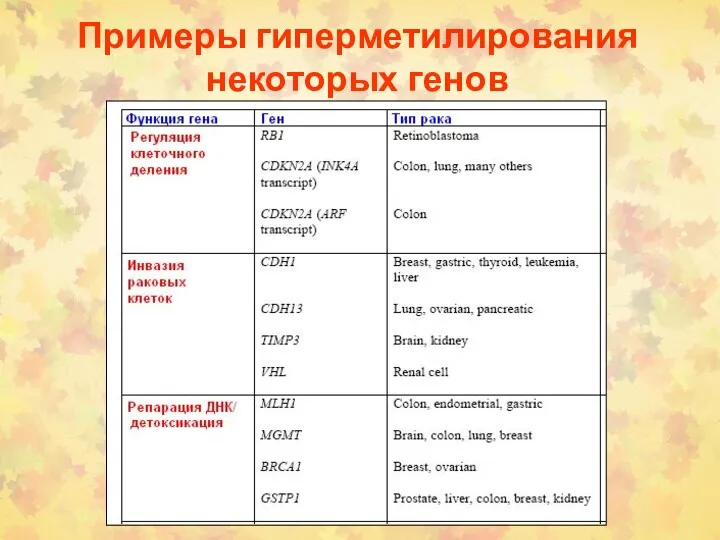
Примеры гиперметилирования некоторых генов
Примеры гиперметилирования некоторых генов

Примеры гиперметилирования некоторых генов
Примеры гиперметилирования некоторых генов
 Особенности межличностного общения с коллегами. Врачи Скорой помощи
Особенности межличностного общения с коллегами. Врачи Скорой помощи ЭКГ–диагностика нарушений сердечного ритма и проводимости
ЭКГ–диагностика нарушений сердечного ритма и проводимости Тұқым қуалайтын ауруларды емдеудің негізгі принциптері. Генотерапия
Тұқым қуалайтын ауруларды емдеудің негізгі принциптері. Генотерапия Артефакты КТ-изображений. (Лекция 2)
Артефакты КТ-изображений. (Лекция 2) Порядок оказания медицинской помощи беременным женщинам с врожденными пороками внутренних органов у плода
Порядок оказания медицинской помощи беременным женщинам с врожденными пороками внутренних органов у плода Болезнь, вызванная новым коронавирусом 2019-nCoV. Клиническая картина. Лечение. Профилактические мероприятия
Болезнь, вызванная новым коронавирусом 2019-nCoV. Клиническая картина. Лечение. Профилактические мероприятия Лабораторно-инструментальные методы исследования сердечно-сосудистой системы
Лабораторно-инструментальные методы исследования сердечно-сосудистой системы Гингивит, породонтит, ісік тәрізді және идиопатиялық породонт ауруларының клиникасы, диагностикасы және потологиялық анатомиясы
Гингивит, породонтит, ісік тәрізді және идиопатиялық породонт ауруларының клиникасы, диагностикасы және потологиялық анатомиясы Амбулатория жағдайында өкпелік гипертензия кезіндегі рационалды дифференциалды диагностика алгоритмі
Амбулатория жағдайында өкпелік гипертензия кезіндегі рационалды дифференциалды диагностика алгоритмі Неспецифическая хирургическая инфекция
Неспецифическая хирургическая инфекция Развитие в онтогенезе. Новорожденность
Развитие в онтогенезе. Новорожденность Предмет, история развития цель и задачи гигиены как науки и области практической деятельности
Предмет, история развития цель и задачи гигиены как науки и области практической деятельности Эндоцервикоз шейки матки
Эндоцервикоз шейки матки Принципы рациональной фармакотерапии артериальной гипертензии у беременных женщин, при гестационной гипертензии
Принципы рациональной фармакотерапии артериальной гипертензии у беременных женщин, при гестационной гипертензии АВС-реанимация. Принципы интенсивной терапии и реанимации новорожденных
АВС-реанимация. Принципы интенсивной терапии и реанимации новорожденных Методы исследования заболеваний органов дыхания (лекция 3)
Методы исследования заболеваний органов дыхания (лекция 3) 27 Физиология сосудистой системы
27 Физиология сосудистой системы Эпидемиологиялық көрсеткіштердін анализі
Эпидемиологиялық көрсеткіштердін анализі Метил спирті. Этил спирті
Метил спирті. Этил спирті Средства, действующие на периферическую нервную систему
Средства, действующие на периферическую нервную систему Профилактика туберкулеза
Профилактика туберкулеза Дизартрия. Определение понятия дизартрия и ее признаки
Дизартрия. Определение понятия дизартрия и ее признаки Общественное здоровье - современная парадигма улучшения жизни через профилактику хронических неинфекционных заболеваний
Общественное здоровье - современная парадигма улучшения жизни через профилактику хронических неинфекционных заболеваний Система гемостаза. Синдром диссеминированного внутрисосудистого свертывания
Система гемостаза. Синдром диссеминированного внутрисосудистого свертывания Роль медицинской сестры в уходе за пациентом после острого нарушения мозгового кровообращения
Роль медицинской сестры в уходе за пациентом после острого нарушения мозгового кровообращения Оказание первой помощи при укусах змей и насекомых
Оказание первой помощи при укусах змей и насекомых Психология детей с задержкой психического развития со слабовыраженными отклонениями в психическом развитии
Психология детей с задержкой психического развития со слабовыраженными отклонениями в психическом развитии Рентгенологическая диагностика патологии лёгких
Рентгенологическая диагностика патологии лёгких