- Неалкогольна жирова хвороба печінки

Содержание
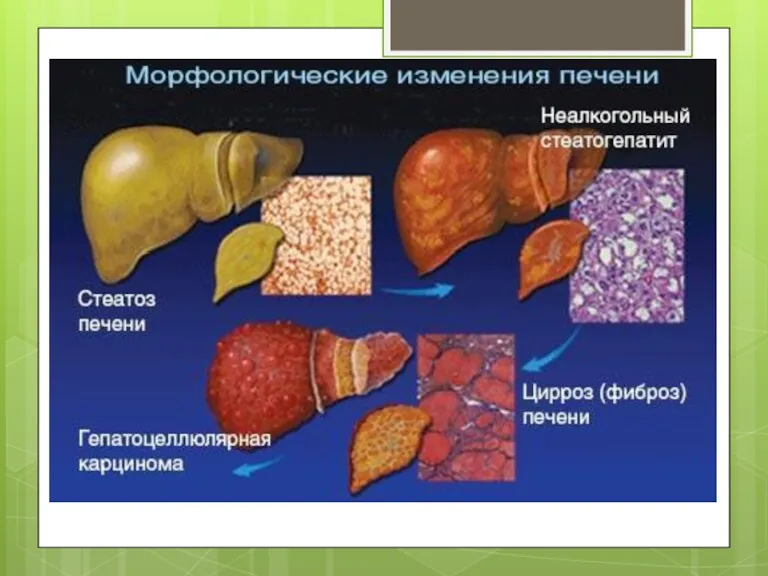
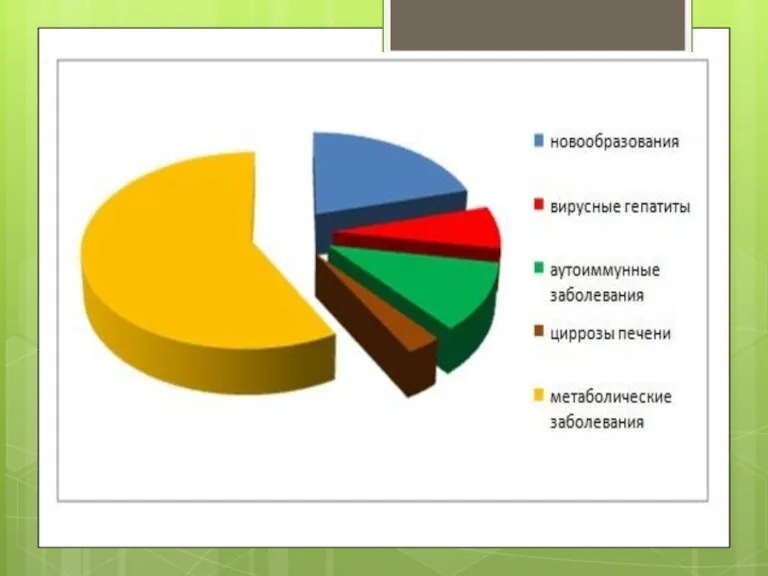
- 2. Неалкогольна жирова хвороба печінки це клініко-патологічний синдромокомплекс, який включає стеатоз печінки (накопичення триацилгліцеролів у печінці більше
- 4. Епідеміологія В Україні Згідно з епідеміологічним дослідженням поширеність НАЖХП становить 10-40 % Приблизно у 20% пацієнтів
- 5. У той час як алкогольна жирова хвороба печінки спостерігається переважно у чоловіків, серед хворих на НАЖХП
- 6. Етіологія Розрізняють два види НАЖХП: первинний і вторинний, які зумовлені ендогенними та екзогенними етіологічними чинниками.
- 7. Первинний ендогенний пов’язаний з метаболічним синдромом. Метаболічний синдром (синдром резистентності до інсуліну) - розлад, пов'язаний з
- 10. Вторинний ендогенний чинник належать синдром мальабсорбції, який ви- никає внаслідок хірургічного лікування при ожи- рінні накладання
- 11. Екзогенні фактори Первинні швидке зменшення маси тіла при лікуванні ожиріння, повне тривале парентеральне живлення(незбалансова за вмістом
- 13. Також провідну роль у розвитку неалкогольної жирової хвороби печінку відіграє дефіцит фолієвої кислоти в гепатоцитах. Відомо
- 14. Патогенез 1.Накопичення тригліцеридів і формування жирового гепатозу.Вивільнення із жирової тканини і синтез в гепатоцитах вільних жирних
- 15. Чинники зовнішнього середовища,ожиріння Инсулинорзистентность Генетичні чинники Цукровий діабет Стеатоз Оксидативний стрес ПУЛ Проникнисть мембран мітохондрій Некроз
- 16. Некроз Воспаление Фиброз Цироз печинки Портальная гипертензия Гепатоцелюлярная карцинома Смерть
- 18. Гомоцистеїн (Hcy) - амінокислота з формулою HSCH2CH2CH(NH2)CO2H. Є гомологом амінокислоти цистеїну, відрізняється від останнього однією метиленовою
- 19. Біосинтез гомоцистеїну Біосинтезується з метіоніну у багатостадійному процесі. Спочатку метіонін алкилює АТФ з утворенням S-аденозил метіоніну(SAM).
- 20. Метаболізм гомоцистеїну Вміст гомоцистеїна забезпечується: а) шляхом реметилювання до метіоніну б) шляхом транссульфування до цистеїну. Реметилювання
- 21. Метаболізм Hcy залежить від чинників - похідних вітамінів. Тому дефіцит будь-якого з вітамінів (В12, фолієвої кислоти
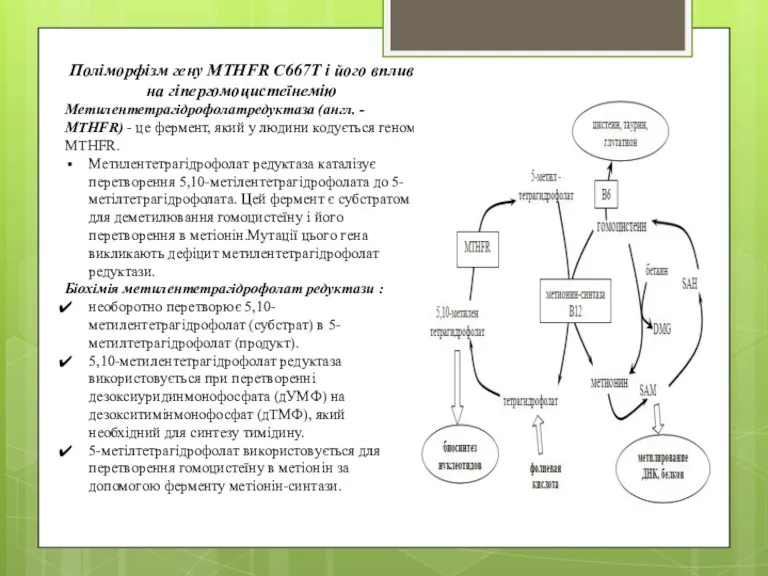
- 22. Поліморфізм гену MTHFR C667T і його вплив на гіпергомоцистеїнемію Метилентетрагідрофолатредуктаза (англ. - MTHFR) - це фермент,
- 23. Структура MTHFR (у ссавців) складається з N-кінцевого каталітичного домену і С-кінцевого регуляторного домену. MTHFR має не
- 24. Люди з 677TT схильні помірним проявам гомоцістінеміі (високий рівень гомоцистеїну в крові), оскільки у них рівень
- 26. Скачать презентацию

Неалкогольна жирова хвороба печінки
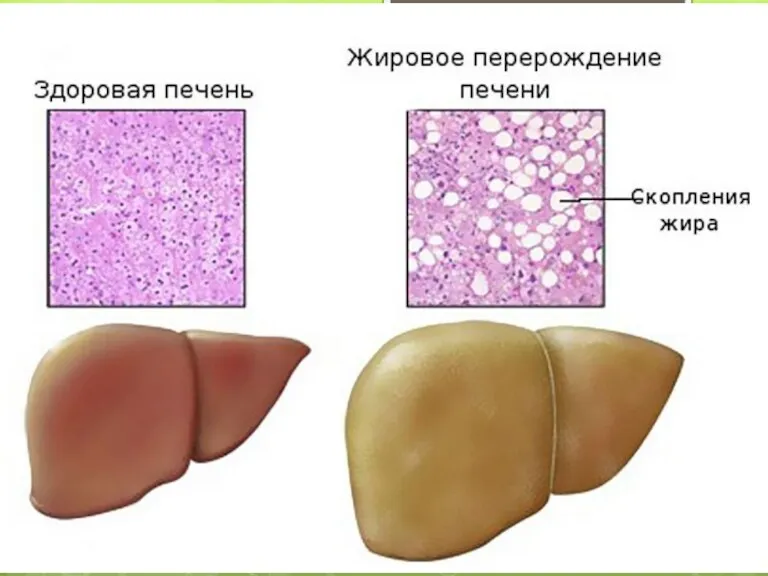
це клініко-патологічний синдромокомплекс, який включає стеатоз печінки (накопичення
Неалкогольна жирова хвороба печінки
це клініко-патологічний синдромокомплекс, який включає стеатоз печінки (накопичення

Епідеміологія
В Україні
Згідно з епідеміологічним дослідженням поширеність НАЖХП становить 10-40 %
Приблизно у
Епідеміологія
В Україні
Згідно з епідеміологічним дослідженням поширеність НАЖХП становить 10-40 %
Приблизно у

У той час як алкогольна жирова хвороба печінки
спостерігається переважно у чоловіків,
У той час як алкогольна жирова хвороба печінки
спостерігається переважно у чоловіків,

Етіологія
Розрізняють два види НАЖХП:
первинний і вторинний, які зумовлені ендогенними та екзогенними
Етіологія
Розрізняють два види НАЖХП:
первинний і вторинний, які зумовлені ендогенними та екзогенними
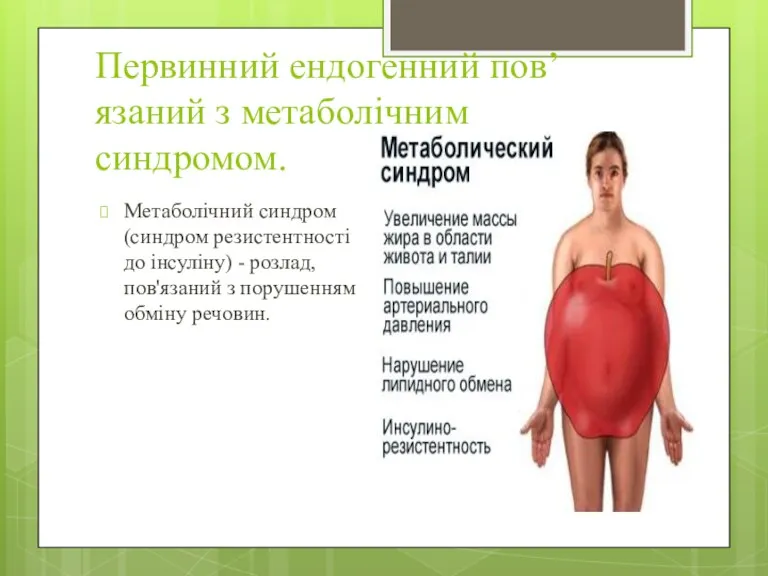
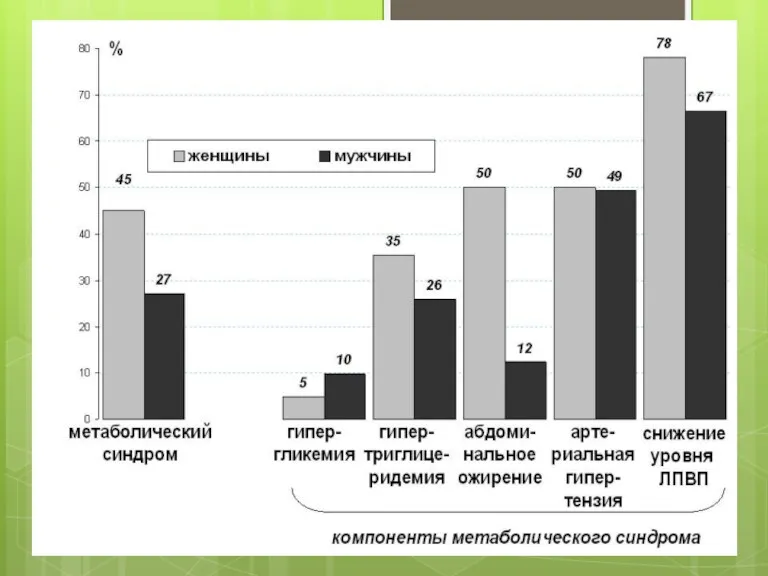
Первинний ендогенний пов’язаний з метаболічним синдромом.
Метаболічний синдром (синдром резистентності до інсуліну)
Первинний ендогенний пов’язаний з метаболічним синдромом.
Метаболічний синдром (синдром резистентності до інсуліну)


Вторинний ендогенний чинник
належать синдром мальабсорбції, який ви-
никає внаслідок хірургічного лікування при
Вторинний ендогенний чинник
належать синдром мальабсорбції, який ви-
никає внаслідок хірургічного лікування при

Екзогенні фактори
Первинні
швидке зменшення маси тіла при лікуванні ожиріння, повне тривале парентеральне
Екзогенні фактори
Первинні
швидке зменшення маси тіла при лікуванні ожиріння, повне тривале парентеральне

Також провідну роль у розвитку неалкогольної жирової хвороби печінку відіграє дефіцит
Також провідну роль у розвитку неалкогольної жирової хвороби печінку відіграє дефіцит

Патогенез
1.Накопичення тригліцеридів і формування жирового гепатозу.Вивільнення із жирової тканини і синтез
Патогенез
1.Накопичення тригліцеридів і формування жирового гепатозу.Вивільнення із жирової тканини і синтез
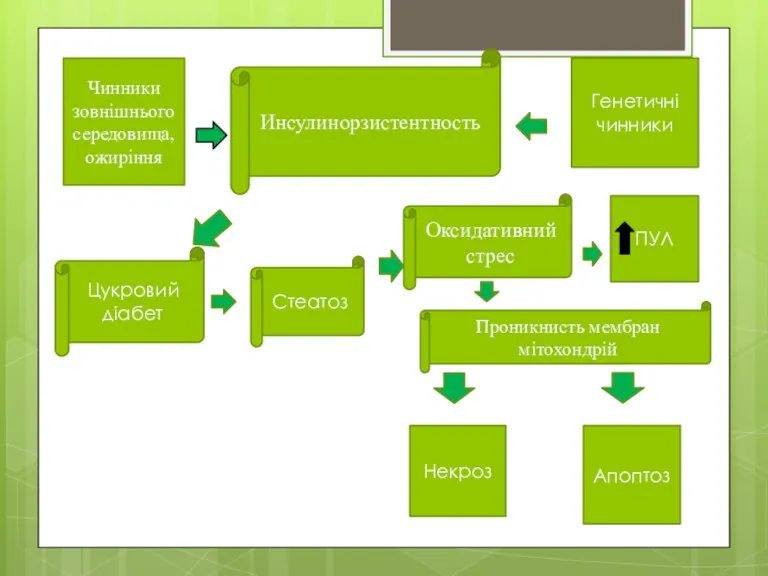
Чинники зовнішнього середовища,ожиріння
Инсулинорзистентность
Генетичні чинники
Цукровий діабет
Стеатоз
Оксидативний стрес
ПУЛ
Проникнисть мембран мітохондрій
Некроз
Апоптоз
Чинники зовнішнього середовища,ожиріння
Инсулинорзистентность
Генетичні чинники
Цукровий діабет
Стеатоз
Оксидативний стрес
ПУЛ
Проникнисть мембран мітохондрій
Некроз
Апоптоз
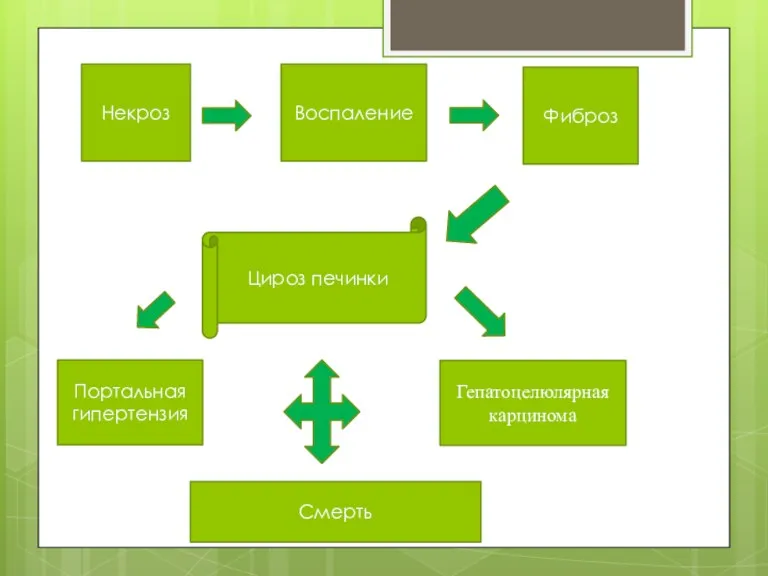
Некроз
Воспаление
Фиброз
Цироз печинки
Портальная гипертензия
Гепатоцелюлярная карцинома
Смерть
Некроз
Воспаление
Фиброз
Цироз печинки
Портальная гипертензия
Гепатоцелюлярная карцинома
Смерть

Гомоцистеїн (Hcy) - амінокислота з формулою HSCH2CH2CH(NH2)CO2H. Є гомологом амінокислоти цистеїну, відрізняється від останнього
Гомоцистеїн (Hcy) - амінокислота з формулою HSCH2CH2CH(NH2)CO2H. Є гомологом амінокислоти цистеїну, відрізняється від останнього

Біосинтез гомоцистеїну
Біосинтезується з метіоніну у багатостадійному процесі.
Спочатку метіонін алкилює АТФ з утворенням S-аденозил метіоніну(SAM).
Далі
Біосинтез гомоцистеїну
Біосинтезується з метіоніну у багатостадійному процесі.
Спочатку метіонін алкилює АТФ з утворенням S-аденозил метіоніну(SAM).
Далі
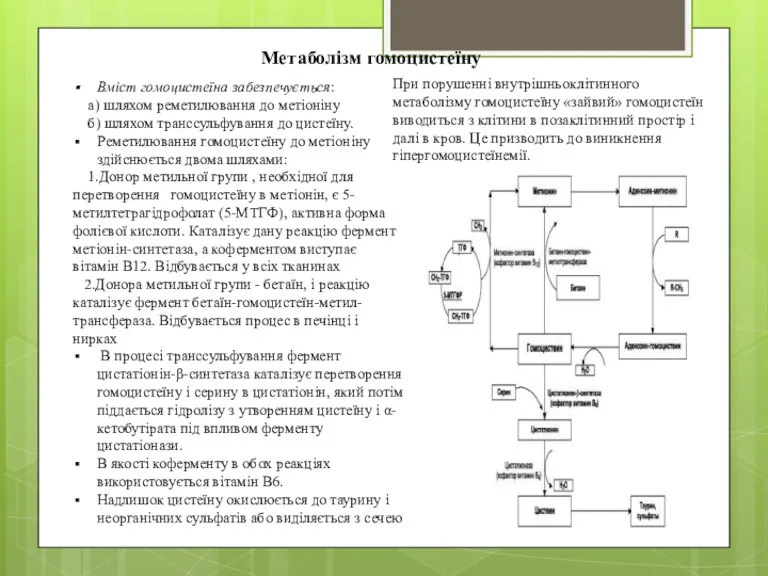
Метаболізм гомоцистеїну
Вміст гомоцистеїна забезпечується:
а) шляхом реметилювання до метіоніну
б)
Метаболізм гомоцистеїну
Вміст гомоцистеїна забезпечується:
а) шляхом реметилювання до метіоніну
б)

Метаболізм Hcy залежить від чинників - похідних вітамінів. Тому дефіцит будь-якого
Метаболізм Hcy залежить від чинників - похідних вітамінів. Тому дефіцит будь-якого
Поліморфізм гену MTHFR C667T і його вплив на гіпергомоцистеїнемію
Метилентетрагідрофолатредуктаза (англ. -
Поліморфізм гену MTHFR C667T і його вплив на гіпергомоцистеїнемію
Метилентетрагідрофолатредуктаза (англ. -

Структура MTHFR (у ссавців) складається з N-кінцевого каталітичного домену і С-кінцевого
Структура MTHFR (у ссавців) складається з N-кінцевого каталітичного домену і С-кінцевого

Люди з 677TT схильні помірним проявам гомоцістінеміі (високий рівень гомоцистеїну в
Люди з 677TT схильні помірним проявам гомоцістінеміі (високий рівень гомоцистеїну в
 Оценка функционального состояния
Оценка функционального состояния Asphyxia
Asphyxia Гастроэзофагальды рефлюкс ауруы
Гастроэзофагальды рефлюкс ауруы Общественное здравоохранение
Общественное здравоохранение Жұқпалы аурумен ауырған малдың ұшасы мен ағзаларын малдәрiгерлiк-санитариялық сараптау
Жұқпалы аурумен ауырған малдың ұшасы мен ағзаларын малдәрiгерлiк-санитариялық сараптау Туберкулез, алкоголизм и наркомания
Туберкулез, алкоголизм и наркомания Коклюш у детей раннего возраста
Коклюш у детей раннего возраста Неспецифический язвенный колит и болезнь Крона
Неспецифический язвенный колит и болезнь Крона Внутрибольничные инфекции. Масштаб, проблемы и структура. Инфекционный процесс
Внутрибольничные инфекции. Масштаб, проблемы и структура. Инфекционный процесс Клинические рекомендации по диагностике, лечению и профилактике тяжелой внебольничной пневмонии у взрослых
Клинические рекомендации по диагностике, лечению и профилактике тяжелой внебольничной пневмонии у взрослых Внебольничная пневмония у взрослых: диагностика, лечение, профилактика
Внебольничная пневмония у взрослых: диагностика, лечение, профилактика Нарушения менструального цикла
Нарушения менструального цикла Комалар кезіндегі диагностикалау және жедел көмек көрсету алгоритмі
Комалар кезіндегі диагностикалау және жедел көмек көрсету алгоритмі Острая дыхательная недостаточность
Острая дыхательная недостаточность Травма шейного отдела позвоночника
Травма шейного отдела позвоночника Дифференциальный диагноз при аритмиях. Клиническая и ЭКГ-диагностика. Принципы лечения и неотложная помощь
Дифференциальный диагноз при аритмиях. Клиническая и ЭКГ-диагностика. Принципы лечения и неотложная помощь Слайд-лекция №12. Нейролептики. Транквилизаторы. Седативные. Соли лития
Слайд-лекция №12. Нейролептики. Транквилизаторы. Седативные. Соли лития Влияние наушников на слух школьников
Влияние наушников на слух школьников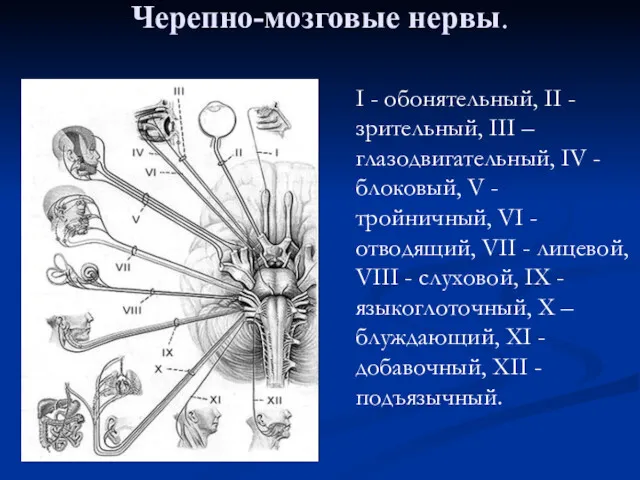 Черепно-мозговые нервы и их поражение
Черепно-мозговые нервы и их поражение Державна санітарноепідеміологічна експертиза товарів для дітей та підлітків
Державна санітарноепідеміологічна експертиза товарів для дітей та підлітків Расстройства восприятия
Расстройства восприятия Авиценна и народная медицина
Авиценна и народная медицина Гастрит. Жедел гастрит
Гастрит. Жедел гастрит Патогенез туберкулеза
Патогенез туберкулеза Кости верхней конечности и их соединения
Кости верхней конечности и их соединения Государственная регистрация лекарственных средств
Государственная регистрация лекарственных средств Плевриты. Инфекционные и неинфекционные плевриты
Плевриты. Инфекционные и неинфекционные плевриты Covid-19. Симптомы и признаки коронавируса у человека
Covid-19. Симптомы и признаки коронавируса у человека