- Реакция отторжения трансплантата

Содержание
- 2. Иммунная природа отторжения чужеродных (прежде всего аллогенных) трансплантатов была доказана в 40-е годы прошлого года P.
- 4. Генетика гистосовместимости Успехи в осуществлении анализа генетических основ тканевой несов- местимости и трансплантационного иммунитета были связаны
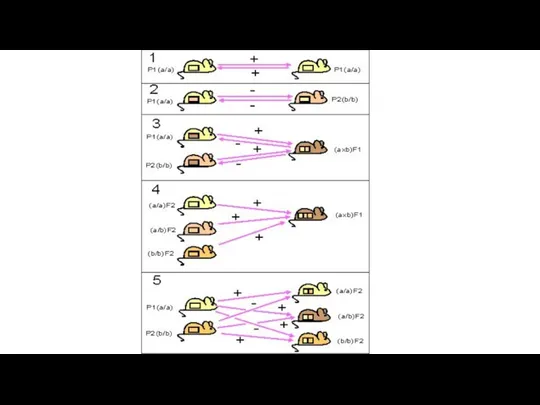
- 5. Выведение достаточного количества инбредных линий мышей позволило сформулировать результаты трансплантации между этими линиями в виде законов
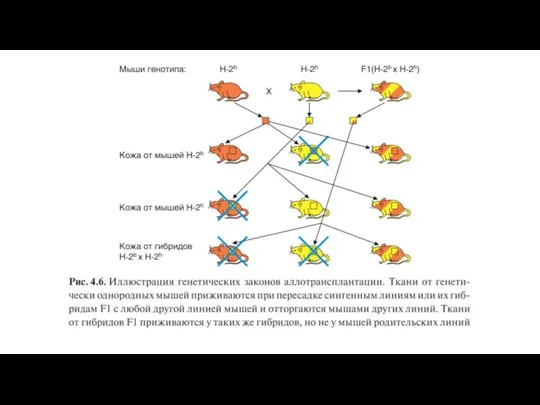
- 7. Известно много (у мышей — 30–40) генетических локусов, обусловливающих тканевую совместимость Однако с различиями только по
- 8. Трансплантационный иммунитет Рассмотрим феноменологию отторжения аллогенных тканей на примере трансплантатов кожи. После подсадки кожного лоскута происходит
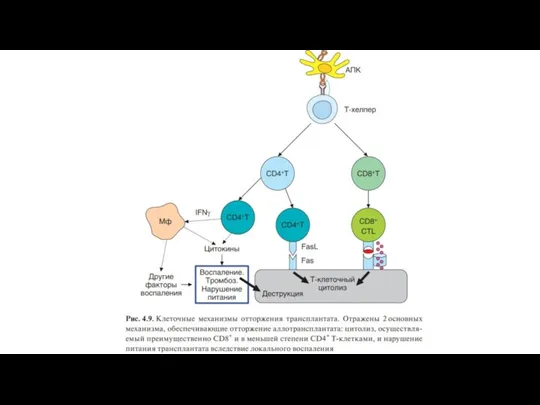
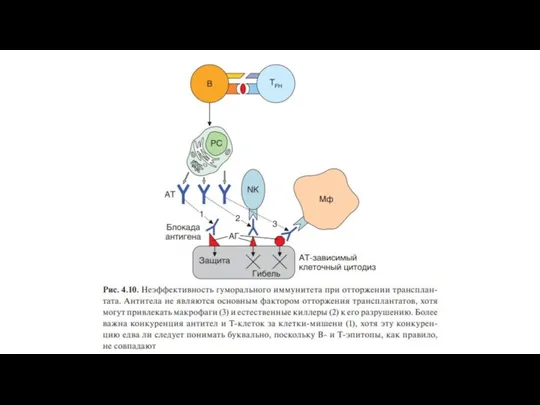
- 9. Трансплантационная реакция сочетает некоторые черты цитотоксической и воспалительной форм клеточного иммунного ответа. Она реализуется с участием
- 11. Формирующиеся эффекторные Т-клетки (Th1-клетки и цитототоксические Т-лимфоциты) поступают в циркуляцию и в результате экспрессии на их
- 13. Повторная подсадка тканей, несущих те же антигены, которые присутствовали в первом трансплантате, вызывает ускоренную реакцию отторжения,
- 14. Еще П. Медавар показал, что гуморальные антитела не играют существенной роли в отторжении аллотраснплантата. При повторной
- 16. Трансплантация костного мозга. Реакция «трансплантат против хозяина» Основные отличия иммунных механизмов отторжения костного мозга от типичных
- 17. В настоящее время общепринятым подходом для воспроизведения РТПХ в экспериментах на мышах служит введение лимфоидных клеток
- 18. Болезнь развивается в двух вариантах — остром и хроническом. Острая болезнь «трансплантат против хозяина» развивается в
- 19. Типичную для экспериментальной РТПХ клиническую картину, состоящую в увеличении селезенки, лимфатических узлов и печени наблюдают достаточно
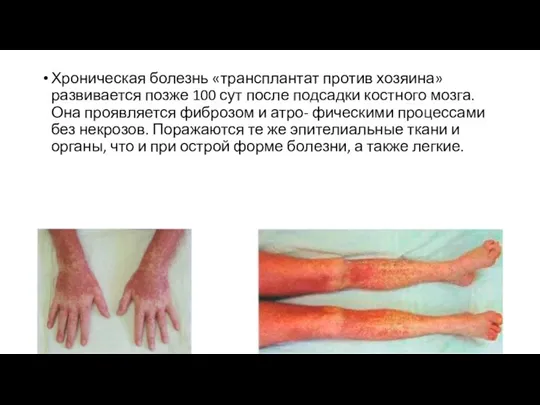
- 20. Хроническая болезнь «трансплантат против хозяина» развивается позже 100 сут после подсадки костного мозга. Она проявляется фиброзом
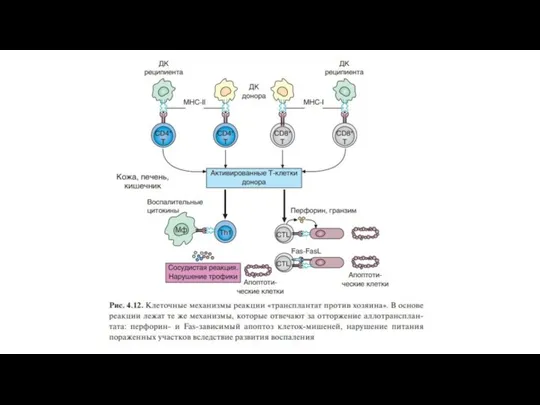
- 21. При острой экспериментальной РТПХ основной мишенью донорских лимфоцитов служат молекулы MHC-II и в первую очередь поражаются
- 24. Скачать презентацию

Иммунная природа отторжения чужеродных (прежде всего аллогенных) трансплантатов была доказана в
Иммунная природа отторжения чужеродных (прежде всего аллогенных) трансплантатов была доказана в

Генетика гистосовместимости
Успехи в осуществлении анализа генетических основ тканевой несов- местимости и
Генетика гистосовместимости
Успехи в осуществлении анализа генетических основ тканевой несов- местимости и

Выведение достаточного количества инбредных линий мышей позволило сформулировать результаты трансплантации между этими линиями
Выведение достаточного количества инбредных линий мышей позволило сформулировать результаты трансплантации между этими линиями

Известно много (у мышей — 30–40) генетических локусов, обусловливающих тканевую совместимость
Однако
Известно много (у мышей — 30–40) генетических локусов, обусловливающих тканевую совместимость
Однако

Трансплантационный иммунитет
Рассмотрим феноменологию отторжения аллогенных тканей на примере трансплантатов кожи. После
Трансплантационный иммунитет
Рассмотрим феноменологию отторжения аллогенных тканей на примере трансплантатов кожи. После

Трансплантационная реакция сочетает некоторые черты цитотоксической и воспалительной форм клеточного иммунного
Трансплантационная реакция сочетает некоторые черты цитотоксической и воспалительной форм клеточного иммунного
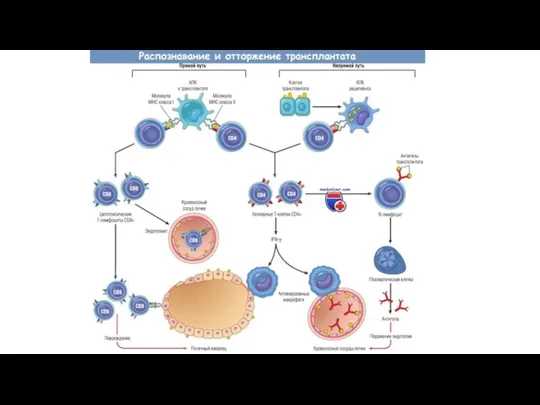

Формирующиеся эффекторные Т-клетки (Th1-клетки и цитототоксические Т-лимфоциты) поступают в циркуляцию и
Формирующиеся эффекторные Т-клетки (Th1-клетки и цитототоксические Т-лимфоциты) поступают в циркуляцию и

Повторная подсадка тканей, несущих те же антигены, которые присутствовали в первом
Повторная подсадка тканей, несущих те же антигены, которые присутствовали в первом

Еще П. Медавар показал, что гуморальные антитела не играют существенной роли
Еще П. Медавар показал, что гуморальные антитела не играют существенной роли

Трансплантация костного мозга. Реакция «трансплантат против хозяина»
Основные отличия иммунных механизмов отторжения
Трансплантация костного мозга. Реакция «трансплантат против хозяина»
Основные отличия иммунных механизмов отторжения

В настоящее время общепринятым подходом для воспроизведения РТПХ в экспериментах на
В настоящее время общепринятым подходом для воспроизведения РТПХ в экспериментах на

Болезнь развивается в двух вариантах — остром и хроническом.
Острая болезнь «трансплантат
Болезнь развивается в двух вариантах — остром и хроническом.
Острая болезнь «трансплантат

Типичную для экспериментальной РТПХ клиническую картину, состоящую в увеличении селезенки, лимфатических
Типичную для экспериментальной РТПХ клиническую картину, состоящую в увеличении селезенки, лимфатических
Хроническая болезнь «трансплантат против хозяина» развивается позже 100 сут после подсадки
Хроническая болезнь «трансплантат против хозяина» развивается позже 100 сут после подсадки

При острой экспериментальной РТПХ основной мишенью донорских лимфоцитов служат молекулы MHC-II
При острой экспериментальной РТПХ основной мишенью донорских лимфоцитов служат молекулы MHC-II
 Производственная практика 2018-2019
Производственная практика 2018-2019 Синдром дисплазии соединительной ткани – синдром Элерса-Данло
Синдром дисплазии соединительной ткани – синдром Элерса-Данло Хирургические методы лечения заболеваний пародонта. Закрытый и открытый кюретаж, гингивотомия и гингивоэктомия
Хирургические методы лечения заболеваний пародонта. Закрытый и открытый кюретаж, гингивотомия и гингивоэктомия Ультразвуковая терапия
Ультразвуковая терапия Средства, влияющие на функции органов пищеварения. Лекция № 14
Средства, влияющие на функции органов пищеварения. Лекция № 14 Студенттің өзіндік жұмысы. Денсаулық сақтау саласында
Студенттің өзіндік жұмысы. Денсаулық сақтау саласында Мезотерапия в эстетической медицине
Мезотерапия в эстетической медицине Нарушения мезентериального кровообращения
Нарушения мезентериального кровообращения Тағамдық уланулар мен токсикоинфекция қоздырғыштары
Тағамдық уланулар мен токсикоинфекция қоздырғыштары Туберкулез мочевыделительной системы
Туберкулез мочевыделительной системы МРТ в гинекологии
МРТ в гинекологии Базовая реанимация
Базовая реанимация Эпидемиологиялық бөртпе сүзегі кезіндегі шаралардың стандарттары мен алгоритмдері
Эпидемиологиялық бөртпе сүзегі кезіндегі шаралардың стандарттары мен алгоритмдері Общая фармакология. Фармакокинетика
Общая фармакология. Фармакокинетика Medical-preventive faculty
Medical-preventive faculty Клиническая анатомия и диагностика заболеваний околоносовых пазух по данным МСКТ
Клиническая анатомия и диагностика заболеваний околоносовых пазух по данным МСКТ Психотерапияның негізгі әдістері
Психотерапияның негізгі әдістері КФ лекарственных средств, применяемых при неотложных состояниях в амбулаторной стоматологической практике
КФ лекарственных средств, применяемых при неотложных состояниях в амбулаторной стоматологической практике Заболевания печени во время беременности
Заболевания печени во время беременности Принципы инфузионной терапии
Принципы инфузионной терапии Лимфогранулематоз (болезнь Ходжкина, злокачественная гранулёма)
Лимфогранулематоз (болезнь Ходжкина, злокачественная гранулёма) Аденомы гипофиза
Аденомы гипофиза Нейроинфекция
Нейроинфекция Ургентные состояния в наркологии
Ургентные состояния в наркологии Бас миының созылмалы ишемиясы
Бас миының созылмалы ишемиясы Виды заболеваний. Фенилпировиноградная олигофрения
Виды заболеваний. Фенилпировиноградная олигофрения Общение в сестринском деле
Общение в сестринском деле Техника забора анализов, правила хранения и транспортировки
Техника забора анализов, правила хранения и транспортировки