- АГ, лизосомы, пероксисомы

Содержание
- 2. Общая схема взаимодействия ЭПР и аппарата Гольджи, 2014
- 3. Три основных типа окаймленных пузырьков Для формирования мембранных пузырьков малого диаметра необходимы специальные белки, сворачивающие мембрану.
- 4. Три основных типа окаймляющих белков Все белки являются отдаленными «родственниками». Разница состоит в том, что СОР
- 5. Формирование пузырьков При формировании мембранных пузырьков сначала образуется «почка», которая потом отшнуровывается с помощью динамина.
- 6. Докование пузырьков
- 7. Слияние мембран с помощью SNARE-комплекса
- 8. Rab-белки Низкомолекулярные (20-25 кД) мономерные ГТФ-азы. Закрепляются в мембране с помощью липидной группы (полуинтегральные белки). Существуют
- 9. Rab-белки - разнообразие Специфические Rab-белки присутствуют в каждом компартменте цитоплазмы. При переносе пузырьков происходит смена Rab-белков
- 10. Транспорт пузырьков 5 этапов переноса – почкование; транспорт пузырька в другой отсек; причаливание; докование; слияние с
- 11. Обмен белков между ЭПР и АГ Обратный путь – для белков-переносчиков, включая СОР I и СОР
- 12. Перенос материала между ЭПР и АГ Перенос белков между ЭПР и АГ осуществляется с помощью замкнутых
- 13. ERGIG – промежуточный комплекс между ЭПР и АГ Места выхода из ЭПР и кластеры ERGIC (промежуточный
- 14. Переход белков от ЭПР к АГ ERES – участки выхода пузырьков из ЭПР; ERGIC – промежуточный
- 15. Эндоцитоз и экзоцитоз Экзоцитоз – выброс секреторных пузырьков из клетки и выделение их содержимого во внешнюю
- 16. Экзоцитоз Секреция (каноническая) – выделение веществ из клетки с помощью мембранных пузырьков. Секреция – конститутивная (постоянная)
- 17. Эндоцитоз – основные типы
- 18. Эндоцитоз Макропиноцитоз – захват больших объемов (~1 мкм3) за счет слияния выдвигающихся мембранных выростов (раффлов). Фагоцитоз
- 19. Микроэндоцитоз и эндосомы
- 20. Клатрин в окаймленных пузырьках
- 21. Формирование эндоцитозного пузырька Формирование эндоцитозного пузырька – сложный процесс разделения мембранных белков, в котором участвуют клатриновый/кавеолиновый
- 22. Клатрин-зависимый эндоцитоз Окаймленные ямки все время находятся на плазматической мембране. Они содержат мало холестерина, и потому
- 23. Цикл эндосом 1. Окаймленные пузырьки частично освобождаются от клатрина и сливаются в ранние эндосомы диаметром 200-500
- 24. Цикл эндосом в живой клетке рН эндосом меняется в зависимости от фазы их цикла
- 25. Лизосомы Открытие – де Дюв, 1960-е г.г. Фракция пузырьков, обогащенная кислой фосфатазой. Размер – 100-1500 нм.
- 26. Лизосомы, ЭМ
- 27. Формирование лизосомы Предшественники лизосом – пузырьки, отщепляющиеся от транс-Гольджи, которые обогащены маннозо-6-фосфатом.
- 28. Формирование вторичных лизосом Начальные этапы формирования вторичных лизосом изучены слабо. Известно несколько путей: слияние поздних эндосом,
- 29. Внутриклеточное «пищеварение»
- 30. Пероксисомы в животной клетке
- 31. Пероксисома в растительной клетке
- 33. Скачать презентацию
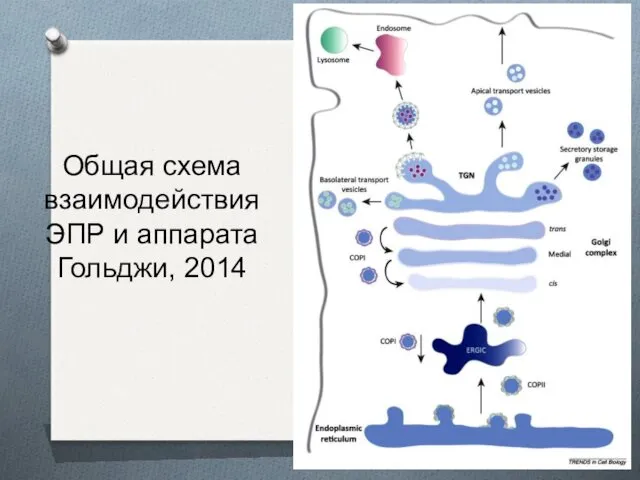
Общая схема взаимодействия ЭПР и аппарата Гольджи, 2014
Общая схема взаимодействия ЭПР и аппарата Гольджи, 2014
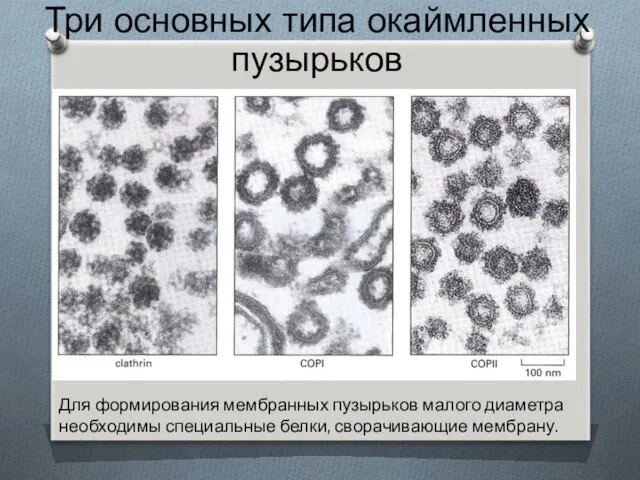
Три основных типа окаймленных пузырьков
Для формирования мембранных пузырьков малого диаметра необходимы
Три основных типа окаймленных пузырьков
Для формирования мембранных пузырьков малого диаметра необходимы

Три основных типа окаймляющих белков
Все белки являются отдаленными «родственниками». Разница состоит
Три основных типа окаймляющих белков
Все белки являются отдаленными «родственниками». Разница состоит
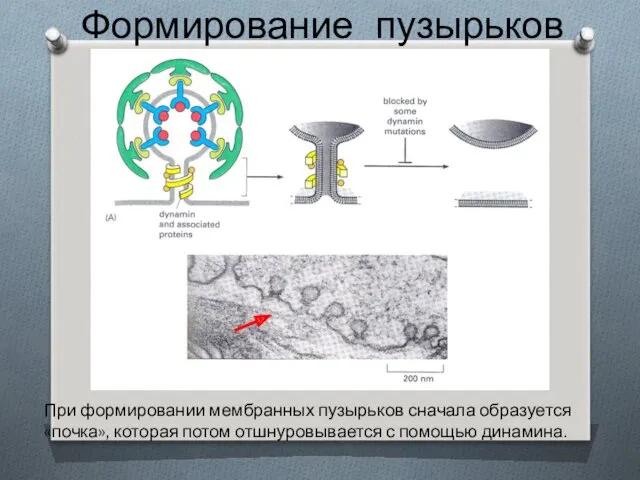
Формирование пузырьков
При формировании мембранных пузырьков сначала образуется «почка», которая потом отшнуровывается
Формирование пузырьков
При формировании мембранных пузырьков сначала образуется «почка», которая потом отшнуровывается
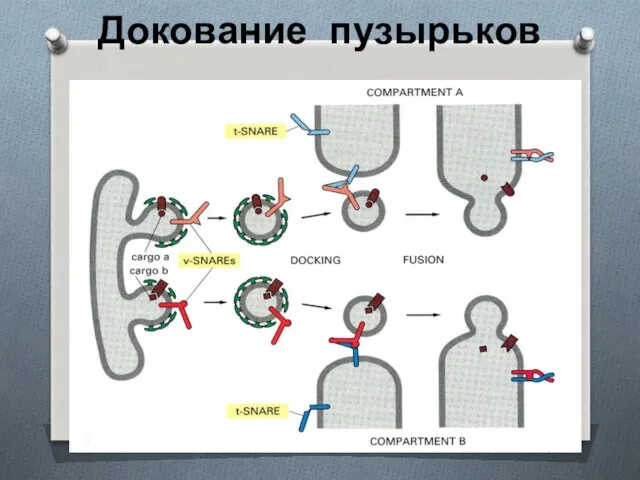
Докование пузырьков
Докование пузырьков
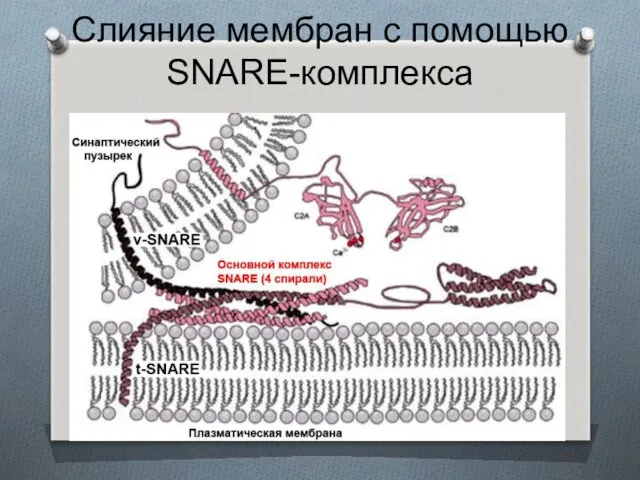
Слияние мембран с помощью SNARE-комплекса
Слияние мембран с помощью SNARE-комплекса
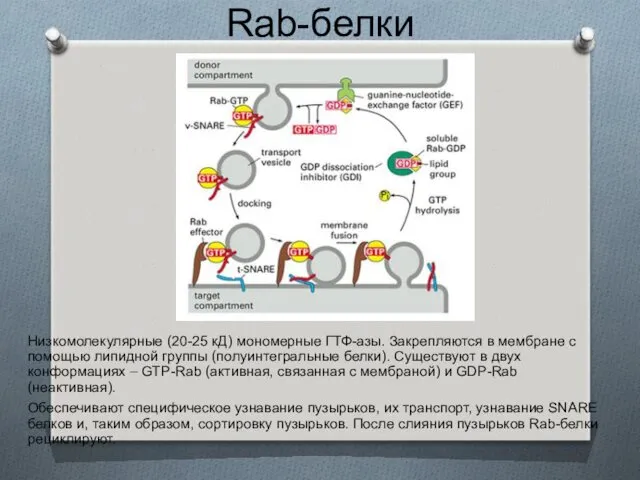
Rab-белки
Низкомолекулярные (20-25 кД) мономерные ГТФ-азы. Закрепляются в мембране с помощью липидной
Rab-белки
Низкомолекулярные (20-25 кД) мономерные ГТФ-азы. Закрепляются в мембране с помощью липидной
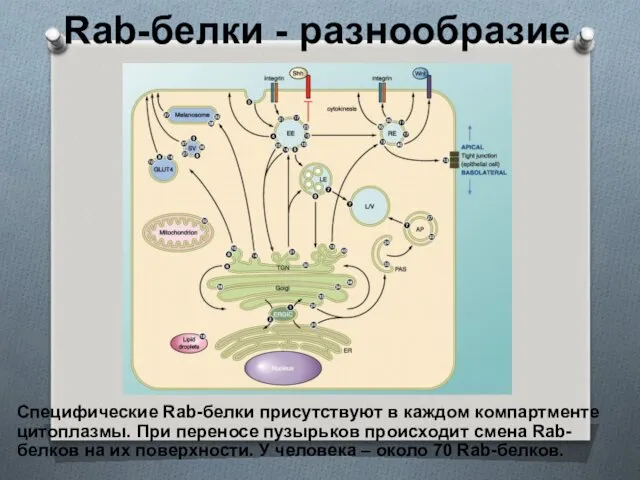
Rab-белки - разнообразие
Специфические Rab-белки присутствуют в каждом компартменте цитоплазмы. При переносе
Rab-белки - разнообразие
Специфические Rab-белки присутствуют в каждом компартменте цитоплазмы. При переносе

Транспорт пузырьков
5 этапов переноса – почкование; транспорт пузырька в другой отсек;
Транспорт пузырьков
5 этапов переноса – почкование; транспорт пузырька в другой отсек;
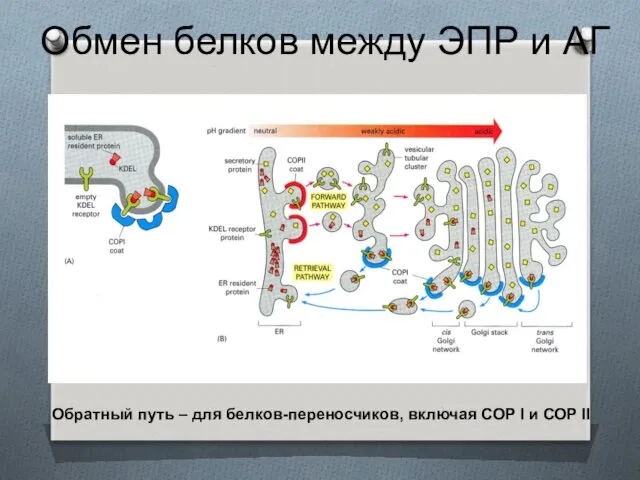
Обмен белков между ЭПР и АГ
Обратный путь – для белков-переносчиков, включая
Обмен белков между ЭПР и АГ
Обратный путь – для белков-переносчиков, включая

Перенос материала между ЭПР и АГ
Перенос белков между ЭПР и АГ
Перенос материала между ЭПР и АГ
Перенос белков между ЭПР и АГ
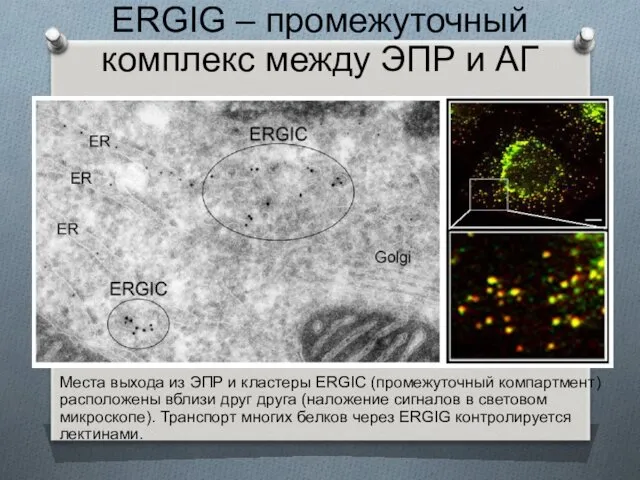
ERGIG – промежуточный комплекс между ЭПР и АГ
Места выхода из ЭПР
ERGIG – промежуточный комплекс между ЭПР и АГ
Места выхода из ЭПР
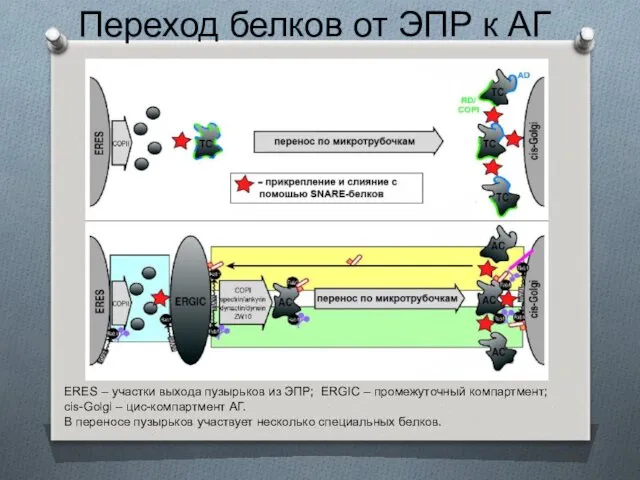
Переход белков от ЭПР к АГ
ERES – участки выхода пузырьков из
Переход белков от ЭПР к АГ
ERES – участки выхода пузырьков из

Эндоцитоз и экзоцитоз
Экзоцитоз – выброс секреторных пузырьков из клетки и выделение
Эндоцитоз и экзоцитоз
Экзоцитоз – выброс секреторных пузырьков из клетки и выделение

Экзоцитоз
Секреция (каноническая) – выделение веществ из клетки с помощью мембранных пузырьков.
Экзоцитоз
Секреция (каноническая) – выделение веществ из клетки с помощью мембранных пузырьков.
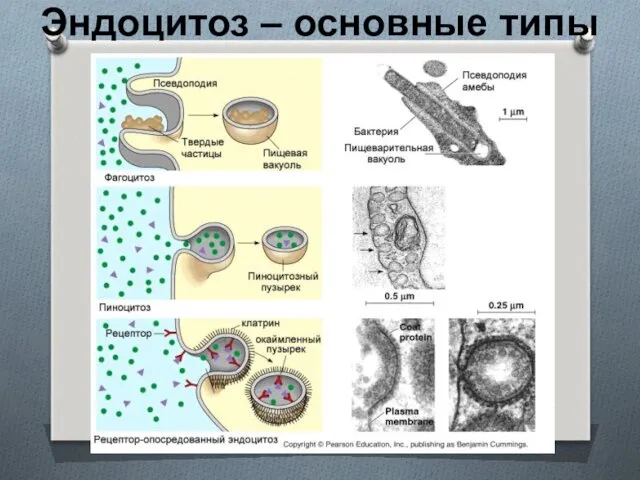
Эндоцитоз – основные типы
Эндоцитоз – основные типы

Эндоцитоз
Макропиноцитоз – захват больших объемов (~1 мкм3) за счет слияния выдвигающихся
Эндоцитоз
Макропиноцитоз – захват больших объемов (~1 мкм3) за счет слияния выдвигающихся
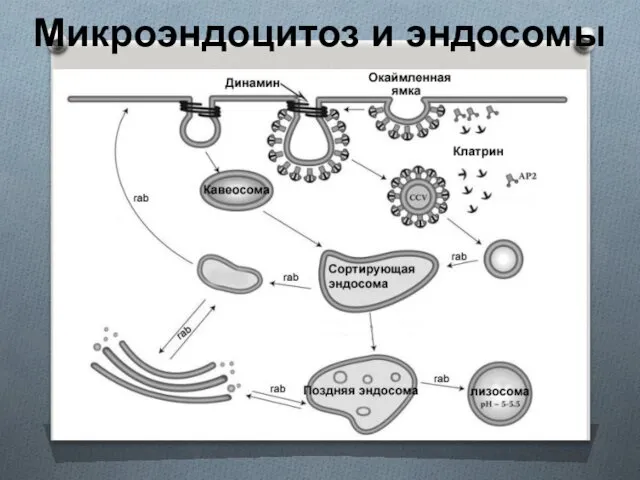
Микроэндоцитоз и эндосомы
Микроэндоцитоз и эндосомы

Клатрин в окаймленных пузырьках
Клатрин в окаймленных пузырьках
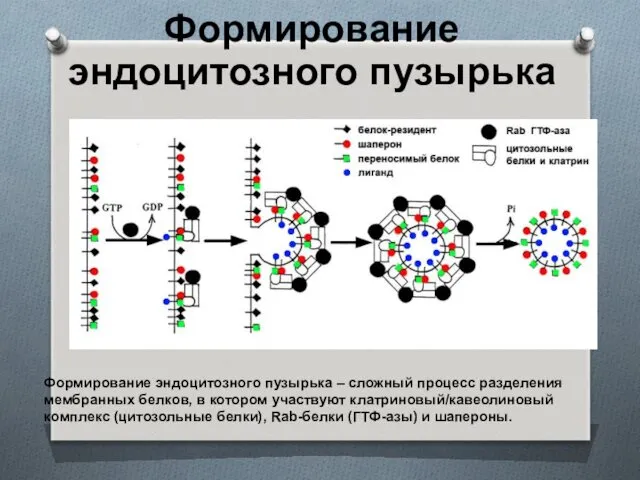
Формирование эндоцитозного пузырька
Формирование эндоцитозного пузырька – сложный процесс разделения мембранных
Формирование эндоцитозного пузырька
Формирование эндоцитозного пузырька – сложный процесс разделения мембранных

Клатрин-зависимый эндоцитоз
Окаймленные ямки все время находятся на плазматической мембране. Они содержат
Клатрин-зависимый эндоцитоз
Окаймленные ямки все время находятся на плазматической мембране. Они содержат

Цикл эндосом
1. Окаймленные пузырьки частично освобождаются от клатрина и сливаются в
Цикл эндосом
1. Окаймленные пузырьки частично освобождаются от клатрина и сливаются в
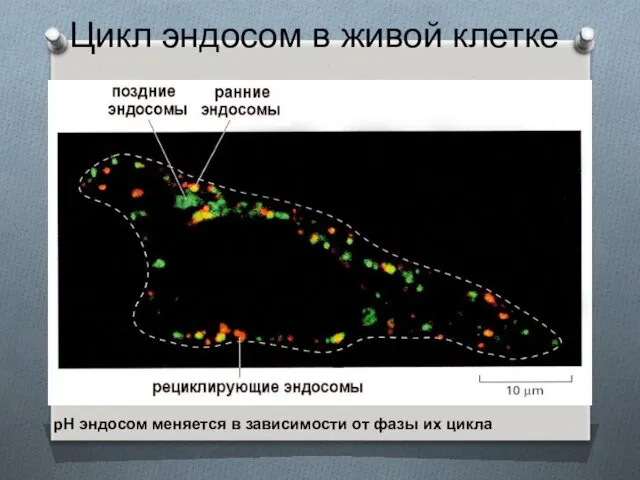
Цикл эндосом в живой клетке
рН эндосом меняется в зависимости от фазы
Цикл эндосом в живой клетке
рН эндосом меняется в зависимости от фазы

Лизосомы
Открытие – де Дюв, 1960-е г.г. Фракция пузырьков, обогащенная кислой
Лизосомы
Открытие – де Дюв, 1960-е г.г. Фракция пузырьков, обогащенная кислой
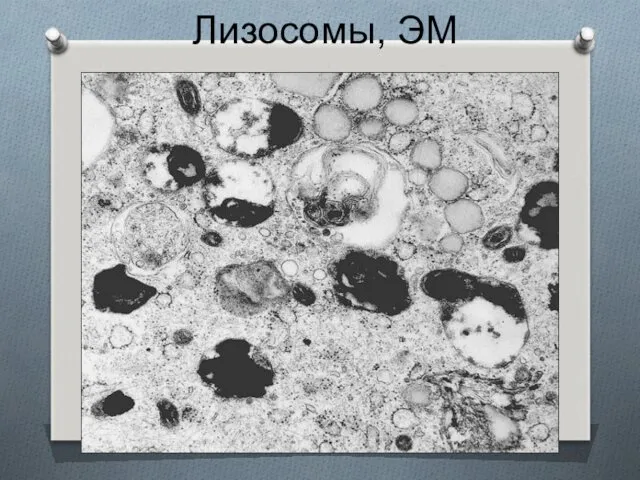
Лизосомы, ЭМ
Лизосомы, ЭМ
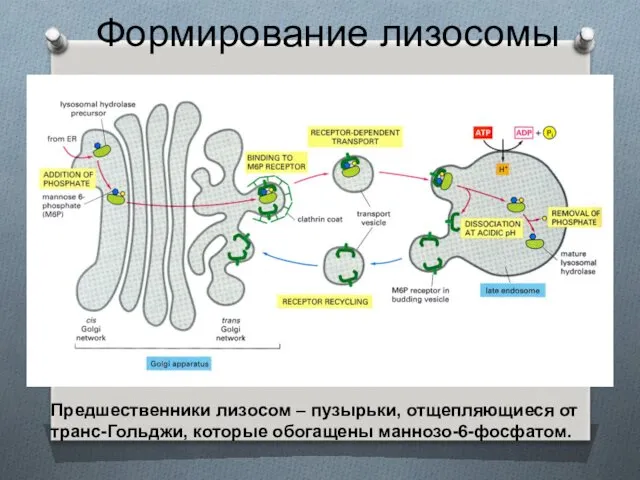
Формирование лизосомы
Предшественники лизосом – пузырьки, отщепляющиеся от транс-Гольджи, которые обогащены маннозо-6-фосфатом.
Формирование лизосомы
Предшественники лизосом – пузырьки, отщепляющиеся от транс-Гольджи, которые обогащены маннозо-6-фосфатом.

Формирование вторичных лизосом
Начальные этапы формирования вторичных лизосом изучены слабо. Известно
Формирование вторичных лизосом
Начальные этапы формирования вторичных лизосом изучены слабо. Известно
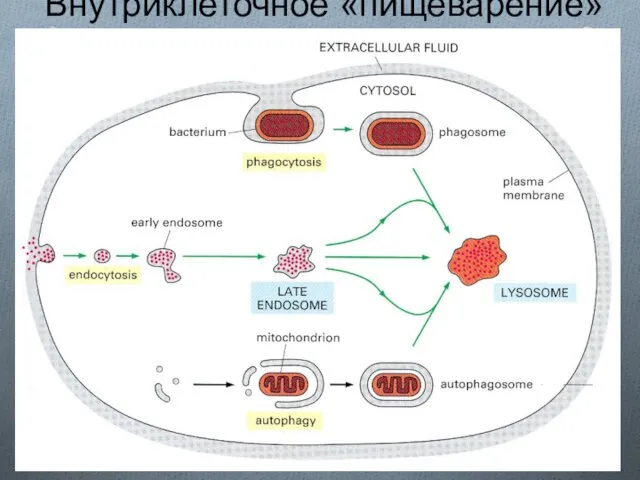
Внутриклеточное «пищеварение»
Внутриклеточное «пищеварение»
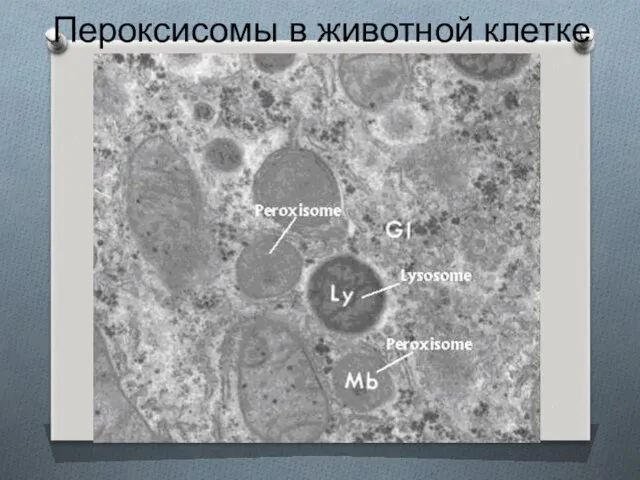
Пероксисомы в животной клетке
Пероксисомы в животной клетке
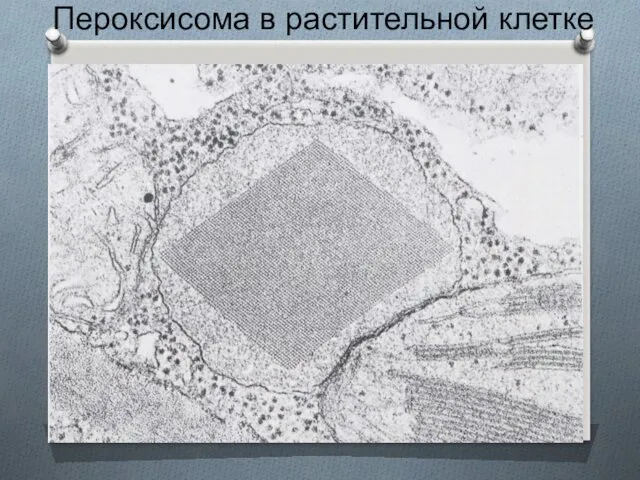
Пероксисома в растительной клетке
Пероксисома в растительной клетке
 Инфекционные заболевания и их классификация
Инфекционные заболевания и их классификация Экологическая безопасность как основа ЗОЖ
Экологическая безопасность как основа ЗОЖ Религия, мировые религии
Религия, мировые религии Контроль качества шероховатости поверхности при различных видах обработки
Контроль качества шероховатости поверхности при различных видах обработки Nice to meet you!
Nice to meet you! Богатства, отданные людям
Богатства, отданные людям Значение транспортного законодательства для страхования грузов и транспортных средств
Значение транспортного законодательства для страхования грузов и транспортных средств Исторические личности IX-XVII веков
Исторические личности IX-XVII веков Экологическая акция Ёлочка, живи!
Экологическая акция Ёлочка, живи! КТД Новогодняя открытка своими руками Новогодний хоровод
КТД Новогодняя открытка своими руками Новогодний хоровод Орфограмма в корне. Готовимся к ГИА
Орфограмма в корне. Готовимся к ГИА Вирус иммунодефицита (ВИЧ
Вирус иммунодефицита (ВИЧ Степенная функция
Степенная функция Альпийские виды лыжного спорта. Скоростной спуск
Альпийские виды лыжного спорта. Скоростной спуск Организация перевозок и управления на транспорте (по видам)
Организация перевозок и управления на транспорте (по видам) Требования безопасности при эксплуатации сосудов, работающих под давлением
Требования безопасности при эксплуатации сосудов, работающих под давлением Типы общественных зданий
Типы общественных зданий терминальные состояния
терминальные состояния Построение (формы) музыки
Построение (формы) музыки Шұжық өндірісі
Шұжық өндірісі Каждой пичужке - моя кормушка
Каждой пичужке - моя кормушка Наш посёлок - Мизур Диск
Наш посёлок - Мизур Диск По одёжке встречают.
По одёжке встречают. Проект Лента Мёбиуса
Проект Лента Мёбиуса Информационный проект Чесма. История морского сражения
Информационный проект Чесма. История морского сражения Конкурс Хочу учиться так! Семинар мечты Финмонополия
Конкурс Хочу учиться так! Семинар мечты Финмонополия Развитие представлений о возникновении жизни на Земле
Развитие представлений о возникновении жизни на Земле Стили речи
Стили речи