- Анализ состава и структуры ДНК

Содержание
- 2. Анализ состава и структуры ДНК Гены локализованы на хромосомах и контролируют проявление признаков в фенотипе. гены
- 3. До 1944 г. о химическом составе и структуре хромосом было известно крайне мало. генетическим материалом могут
- 4. 1944 – 1953 г. во многих лабораториях были получены результаты, позволяющие ответить на вопрос о роли
- 5. Первые исследования генетического материала В конце девятнадцатого века стали активно развиваться исследования химической структуры биомолекул. до
- 6. Первым в 1868 г. исследовал ДНК шведский химик Фридрих Мишер. Из суспензии клеточных ядер он выделил
- 7. в 40-х годах ХХ века Эрвин Чаргафф убедительно показал, что ДНК у большинства организмов содержит неодинаковые
- 8. Доказательство ведущей роли ДНК у бактерий и бактериофагов 1944 г. Освальд Эвери, Колин Мак-Леод и Маклин
- 9. Опыты по трансформации 1927 г. сотрудник Британского Министерства здравоохранения Фредерик Гриффит проводил опыты с различными штаммами
- 10. различия в вирулентности пневмококков обусловлены наличием полисахаридной капсулы: вирулентные клетки имели капсулу, невирулентные – нет.
- 11. Вирулентные и невирулентные штаммы различались по морфологии клеток: инкапсулированные формировали гладкие блестящие колонии (S) бактерии без
- 12. Каждый штамм Diplococcus, вместе с другими подобными штаммами, относится к определенному серотипу. Специфичность серотипа определяется химическим
- 13. пневмонию вызывают только живые вирулентные клетки. Если мышам ввести убитые нагреванием до 65⁰С вирулентные бактерии, то
- 14. эти бактерии идентичны клеткам IIIS, убитым перед инъекцией нагреванием. Контрольные мыши, которым вводили живые невирулентные клетки
- 15. Гриффит предположил, что убитые клетки каким-то образом превращают невирулентные бактерии в вирулентный тип IIIS. Он назвал
- 16. В начале 30-х годов Генри Доусон с сотрудниками показали, что трансформация возможна также in vitro, в
- 17. 1944 г., были опубликованы результаты, полученные Эвери, Мак-Леодом и Мак-Карти. Им удалось выделить трансформирующий фактор и
- 18. Сначала исследователи выращивали большие объемы (50 – 70 л) жидких культур вирулентных бактерий типа IIIS. Затем
- 19. Эвери с коллегами решили, что трансформирующий фактор взаимодействует с клетками типа IIR и координирует, таким образом,
- 20. трансформация происходит у Hemophilus influenzae, Bacillus subtilis, Sigella paradysenteriae, Escherichia coli. трансформация морфологии колоний устойчивость к
- 21. Эксперимент Херши-Чейз Другие доказательства роли ДНК как переносчика генетической информации были получены при исследовании бактериофага Т2,
- 22. В 1952 г. Альфред Херши и Марта Чейз показали, что в репродукции фага особую роль занимает
- 23. Из предыдущих исследований было известно, что: 1. Фаги Т2 примерно на 50% состоят из белков и
- 24. Если предположить, что некоторые из молекулярных компонентов фага (ДНК или белки) проникают внутрь бактерии, то какие
- 25. Херши и Чейз использовали радиоизотопный метод. Они метили ДНК радиоактивным фосфором (32Р), а белки – радиоактивной
- 26. Из опыта Херши и Чейз сделали вывод, что белки фага остаются снаружи бактериальной клетки-хозяина, а для
- 27. Эксперименты Херши и Чейз, а также Эвери с сотрудниками убедительно показали, что носителями наследственности служат молекулы
- 28. Опыты по трансфекции 1957 г. – опубликовано несколько работ, которые показали, что после ферментативной обработки клеток
- 29. 1960 г. – проведены сходные эксперименты с использованием очищенной фаговой ДНК. Заражение клеток-хозяев вирусной нуклеиновой кислотой
- 30. Прямые и непрямые доказательства значения ДНК у эукариот В 50-х годах ХХ века эксперименты, демонстрирующие роль
- 31. Непрямое доказательство: распределение ДНК ДНК локализована в клетке там, где определяются определенные генетические функции, а белки
- 32. Очевидна тесная корреляция между количеством хромосом и содержанием ДНК в клетках. Однако такой корреляции по содержанию
- 33. Непрямое доказательство: мутагенез Ультрафиолетовый свет (УФ) – один из агентов, индуцирующих мутации генетического материала. Можно облучить
- 34. УФ обладает максимальным мутагенным действием при длине волны 260 нанометров (нм). Наиболее сильно поглощают свет в
- 35. Прямое доказательство: анализ рекомбинантных ДНК доказательства этой гипотезы были получены с помощью метода рекомбинантных ДНК. Выделенные
- 36. в оплодотворенные мышиные яйцеклетки с помощью микроинъекций можно ввести ген человеческого бета-глобина. продукты этого гена обнаружили
- 37. Позже в оплодотворенные мышиные яйцеклетки ввели ген rat, кодирующий гормон роста. Около трети мышей, выросших из
- 38. В эукариотических клетках происходит экспрессия генетической информации записанной в ДНК.
- 39. Структурный анализ ДНК 1953 г. Джеймс Уотсон и Фрэнсис Крик предположили, что молекула ДНК двуспиральная. Nature.
- 40. В распоряжении Уотсона и Крика было две группы данных: (1) результаты анализа состава оснований в молекулах
- 41. Химия нуклеиновых кислот Когда Уотсон и Крик работали над своей моделью ДНК, уже был известен химический
- 42. Звеньями, или строительными блоками в молекуле всех нуклеиновых кислот служат нуклеотиды. Они состоят из трех компонентов:
- 43. В состав молекул рибонуклеиновой кислоты (РНК) входит сахар рибоза, а в состав дезоксирибонуклеиновой кислоты (ДНК) –
- 44. Молекулы нуклеозидов состоят из пуриновых или пиримидиновых оснований, а также рибозы или дезоксирибозы. Молекулы нуклеотидов представлены
- 45. В зависимости от количества присоединенных фосфатных групп различают: нуклеозидмонофосфат (НМФ), нуклеозиддифосфат (НДФ) нуклеозидтрифосфат (НТФ).
- 46. Нуклеотиды соединяются в полимерные цепочки с помощью фосфодиэфирных связей. Короткие цепочки из одного-двух десятков нуклеотидов называются
- 47. Химический состав оснований В 1949 – 1953 гг. Эдвин Чаргафф с сотрудниками обнаружили с помощью хроматографии,
- 48. Если молекулы облучают рентгеновскими лучами, то часть Х-лучей, сталкиваясь с атомами, рассеивается, и на фотопленку появляются
- 49. В 1950 – 1953 гг. Розалинда Франклин, работавшая в лаборатории Мориса Уилкинса, исследовала более чистые образцы
- 50. Модель Уотсона-Крика Уотсон и Крик в 1953 г. предположили, что ДНК имеет форму двойной спирали со
- 51. Согласно правилам Чаргаффа, молярное содержание А равно молярному содержанию Т, а молярное содержание G – содержанию
- 52. Специфичность спаривания между этими основаниями обусловлена комплементарностью – химическим сродством, обусловленным водородными связями между основаниями. Водородные
- 53. Более точный анализ структуры ДНК, проведенный впоследствии, выявил небольшие отклонения от классической модели. Так, на один
- 54. Уже в 1953 г. авторы заметили, что из предлагаемого ими принципа комплементарности следует возможный механизм копирования
- 55. Альтернативные формы ДНК При выделении ДНК в разных условиях можно получить различные конформации молекул. Во время
- 56. С помощью тонких методов исследовали структуру А-ДНК преобладает в концентрированных растворах с высокой ионной силой или
- 57. 1979 г. Эндрю Ванг и Александр Рич открыли Z-конформацию Молекула Z-ДНК представлена в виде левозакрученной спирали
- 58. присутствие Z-ДНК in vivo пока не доказано
- 59. Структура РНК Иногда РНК образует двуцепочечные структуры: молекула складывается по длине и между комплеиентарными основаниями двух
- 60. Известно три основных класса РНК, функционирующих в клетке: рибосомная РНК (рРНК), информационная, или матричная РНК (иРНК,
- 61. Самые крупные рРНК обычно составляют около 80% всей клеточной РНК. Они являются важными структурными компонентами рибосом,
- 62. Молекулы мРНК переносят генетическую информацию на рибосомы, где происходит трансляция, они также различны по длине в
- 63. Небольшие молекулы тРНК во время трансляции переносят к рибосомам аминокислоты – материал для синтеза белков.
- 64. малые ядерные РНК (snRNA, или мяРНК) – участвуют в процессинге иРНК. РНК-теломераза – вовлечена в репликацию
- 65. РНК в качестве генетического материала некоторых вирусов 1956 г. – вирус табачной мозаики (ВТМ) относится к
- 66. вирус иммунодефицита человека (ВИЧ), вызывающий СПИД, некоторые РНК-содержащие онгогенные (опухолеродные) вирусы.
- 67. Картирование генов у бактерий и бактериофагов
- 68. Бактерии и бактериофаги очень удобны для генетических исследований. Короткий репродуктивный цикл: за короткое время можно получить
- 69. Мутации у бактерий и рост популяции бактериальных клеток Растущие в культуре бактериальные клетки могут размножаться в
- 70. 1943 г. Сальвадор Луриа и Макс Дельбрюк представили первые доказательства возможности спонтанных мутаций у бактерий по
- 71. Бактерий выращивают в жидкой среде или в агаре на чашках Петри. Минимальная среда, необходимая для роста
- 72. Для количественного анализа бактериальной культуры часть клеток переносят в свежую жидкую среду. Сначала клетки растут медленно
- 73. Для подсчета клеток определенный объем бактериальной культуры высевают из жидкой среды в чашки Петри, на плотную
- 74. С разведением культуры плотность клеток падает и соответственно уменьшается число клеток, дающих начало отдельным колониям. Число
- 75. Генетическая рекомбинация у бактерий: конъюгация 1946 г. Джошуа Ледерберг и Эдвард Татум обнаружили конъюгацию – процесс
- 76. Ледерберг и Татум работали с различными ауксотрофными штаммами E. coli K12. штамм А: метионин и биотин
- 77. В эксперименте прототрофные клетки возникали с частотой 1/107 (10-7).
- 78. Бактерии F+ И F--типа Многочисленные эксперименты, проведенные вслед за опытами Ледерберга и Татума, показали, что между
- 79. для передачи генов необходим тесный контакт двух клеток. Бернард Дэвис доказал это с помощью стеклянной трубки,
- 80. Джошуа и Эстер Ледерберги, Вильям Найес и Лукас Кавалли-Сфорца после конъюгации и генетической рекомбинации F-фактор несут
- 81. Итак схему опытов Ледерберга и Татума можно представить следующим образом: Штамм А Штамм В F+ x
- 82. Анализ структуры выделенного из бактерий F-фага показал, что он состоит из кольцевой двухцепочечной ДНК и занимает
- 83. Считается, что перенос F-фактора во время конъюгации бактерий включает образование цитоплазматического мостика – конъюгационной трубки между
- 84. Бактериальные штаммы Hfr и хромосомное картирование 1950 г. Кавалли-Сфорца воздействовал на F+-клетки азотистым соединением, полученным из
- 85. Если бактерий-доноров из штамма Hfr скрещивать с бактериями-реципиентами, то в результате генетической рекомбинации последние не превращаются
- 86. Оказалось, что такая неслучайная рекомбинация затрагивает в различных штаммах Hfr различные гены. Хайес объяснил эти различия
- 87. В середине 50-х годов ХХ века Элли Вольман и Франсуа Жакоб объяснили различия между F+- и
- 88. метод прерывания скрещивания. В первые 8 минут после смешивания культур генетической рекомбинации между поступившими от донора
- 89. расстояние между линейно расположенными генами на хромосоме E.coli можно измерить в минутах и определить экспериментально.
- 90. Сходные результаты были получены Вольман и Жакобом и на других штаммах Hfr, однако обнаружились важные различия.
- 91. Основное различие касается точки начала переноса (О) и направления переноса генов из клетки в клетку. Схема
- 92. Вольман и Жакоб предположили, что хромосома E.coli представляет собой замкнутое кольцо, а точка начала переноса варьирует
- 93. После репликации гены донорской ДНК могут рекомбинировать с гомологичными генами клетки-реципиента.
- 94. Метод прерывания скрещивания позволил картировать всю хромосому E.coli. хромосомная карта штамма К12 длиной 100 минут включает
- 95. F’-элемент и мерозиготы 1959 г. Эдвард Адельберг в экспериментах с клетками Hfr E.coli обнаружил, что интегрированный
- 96. Присутствие бактериальных генов в составе эписомы (F-фактора) приводит к тому, что бактерии типа F’ ведут себя
- 97. F-факторы и плазмиды Когда экстрахромосомный F-фактор находится в цитоплазме бактериальной клетки, он состоит из двуспиральной кольцевой
- 98. Классификация плазмид: F-фактор – содержит гены, определяющие фертильность и ответственные за формирование половых пилей, важных для
- 99. Большинство R-плазмид содержит два компонента: фактор переноса устойчивости – RTF (resistance transfer factor) и r-детерминанты или
- 100. Плазмиды типа Col, в частности, ColE1, кодируют от одного до нескольких белков – колицинов, которые очень
- 101. Бактериальная трансформация В результате трансформации происходит генетическая рекомбинация между бактериальными клетками.
- 102. включает несколько этапов (1) попадание фрагментов ДНК в клетку-реципиента (2) рекомбинация между этой ДНК и гомологичными
- 103. После поступления фрагмента ДНК в бактериальную клетку одна из нитей двуспиральной молекулы разрушается нуклеазами, а другая
- 104. После интеграции чужеродной ДНК в бактериальную хромосому она несет одну исходную цепь ДНК и одну чужую
- 105. Трансформация и сцепленные гены Для эффективности трансформации фрагмент ДНК должен содержать от 10 000 до 20
- 106. трансформация характерна для многих бактерий, включая Diplococcus pneumoniae, Hemophilus influenzae, Bacillus subtilis, Shigella paradysenteriae и E.coli.
- 107. Генетические исследования бактериофагов Бактериофаги, или фаги – это бактериальные вирусы, которые могут участвовать в передаче генетической
- 108. Фаг Т4: структура и жизненный цикл Бактериофаг Т4 – представитель группы близких бактериальных вирусов – Т-четных
- 109. Жизненный цикл фага T4 начинается с момента абсорбции фага на бактериальной клетке. Жизненный цикл бактериофага. С
- 110. Сборку фаговых частиц подробно изучали Вильям Вуд, Роберт Эдгар и другие исследователи, которые выделили три последовательных
- 111. После того как закончится сборка около 200 вирусных частиц, бактериальная клетка разрушается под действием лизоцима –
- 112. Метод бляшек В одном мл инфицированной культуры, например, содержится более 1010 вирусных частиц. Количественный анализ фагов
- 113. проводят серийное разведение инфицированной бактериальной культуры. по 0,1 мл каждого из разведений наносят в жидкий агар
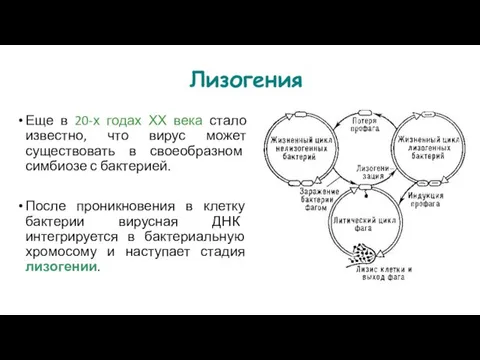
- 114. Лизогения Еще в 20-х годах ХХ века стало известно, что вирус может существовать в своеобразном симбиозе
- 115. Вирусная ДНК интегрирует в хромосому в виде профага. Вирусы, которые могут лизировать бактерии, а также встраиваться
- 116. Трансдукция: перенос бактериальной ДНК вирусом 1952 г. Нортон Зиндер и Джошуа Ледерберг исследовали рекомбинацию у бактерии
- 117. Эксперимент Зиндера-Ледерберга Ледерберг и Зиндер смешали между собой два ауксотрофных штамма Salmonella LA-22 и LA-2 и
- 118. После совместной инкубации в трубке Дэвиса штаммы поместили на разные чашки с минимальной средой и обнаружили,
- 119. С учетом этих наблюдений, а также возможности инфекции сальмонеллы умеренными фагами, исследователи предположили, что рекомбинация между
- 120. Природа трансдукции трансдукция у E.coli связана с инфекцией фагами P1 и λ, а Bacillus subtilis и
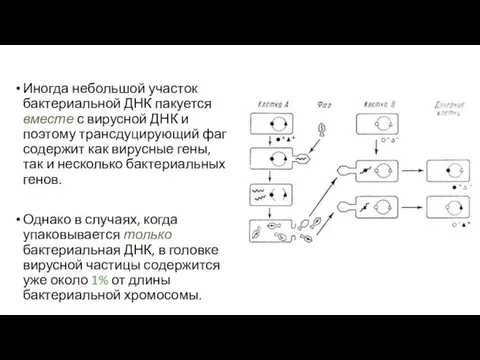
- 121. Иногда небольшой участок бактериальной ДНК пакуется вместе с вирусной ДНК и поэтому трансдуцирующий фаг содержит как
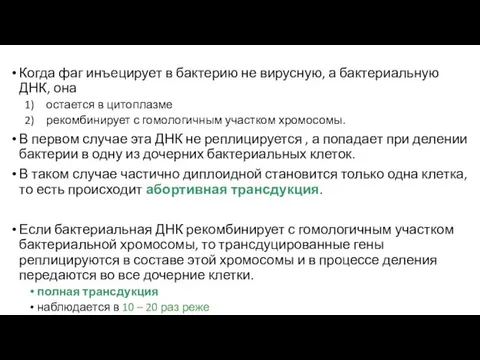
- 122. Когда фаг инъецирует в бактерию не вирусную, а бактериальную ДНК, она остается в цитоплазме рекомбинирует с
- 123. Трансдукционное картирование Фрагменты бактериальной ДНК, вовлеченные в трансдукцию, достаточно велики и содержат целый ряд генов. В
- 124. Межгенная рекомбинация и картирование у бактериофагов К 1947 г. сразу несколько исследовательских групп обнаружили у бактериофагов
- 125. Мутации у бактериофагов 1946 г. Альфред Херши наблюдал необычные по морфологии бляшки на агаре с клетками
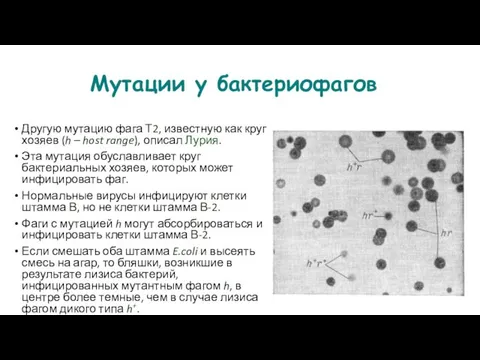
- 126. Мутации у бактериофагов Другую мутацию фага Т2, известную как круг хозяев (h – host range), описал
- 127. Межгенное картирование Генетическую рекомбинацию у бактериофагов открыли во время экспериментов по одновременной смешанной инфекции бактерий двумя
- 129. Скачать презентацию

Анализ состава и структуры ДНК
Гены локализованы на хромосомах и контролируют проявление
Анализ состава и структуры ДНК
Гены локализованы на хромосомах и контролируют проявление

До 1944 г. о химическом составе и структуре хромосом было известно
До 1944 г. о химическом составе и структуре хромосом было известно

1944 – 1953 г. во многих лабораториях были получены результаты, позволяющие
1944 – 1953 г. во многих лабораториях были получены результаты, позволяющие

Первые исследования генетического материала
В конце девятнадцатого века стали активно развиваться исследования
Первые исследования генетического материала
В конце девятнадцатого века стали активно развиваться исследования

Первым в 1868 г. исследовал ДНК шведский химик Фридрих Мишер.
Из суспензии
Первым в 1868 г. исследовал ДНК шведский химик Фридрих Мишер.
Из суспензии

в 40-х годах ХХ века Эрвин Чаргафф убедительно показал, что ДНК

Доказательство ведущей роли ДНК у бактерий и бактериофагов
1944 г. Освальд Эвери,
Доказательство ведущей роли ДНК у бактерий и бактериофагов
1944 г. Освальд Эвери,
Опыты по трансформации
1927 г. сотрудник Британского Министерства здравоохранения Фредерик Гриффит проводил
Опыты по трансформации
1927 г. сотрудник Британского Министерства здравоохранения Фредерик Гриффит проводил

различия в вирулентности пневмококков обусловлены наличием полисахаридной капсулы:
вирулентные клетки имели
различия в вирулентности пневмококков обусловлены наличием полисахаридной капсулы:
вирулентные клетки имели
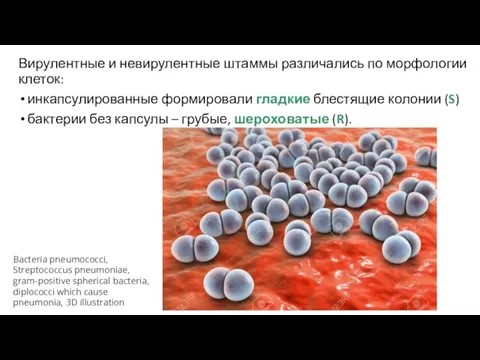
Вирулентные и невирулентные штаммы различались по морфологии клеток:
инкапсулированные формировали гладкие
Вирулентные и невирулентные штаммы различались по морфологии клеток:
инкапсулированные формировали гладкие

Каждый штамм Diplococcus, вместе с другими подобными штаммами, относится к определенному
Каждый штамм Diplococcus, вместе с другими подобными штаммами, относится к определенному
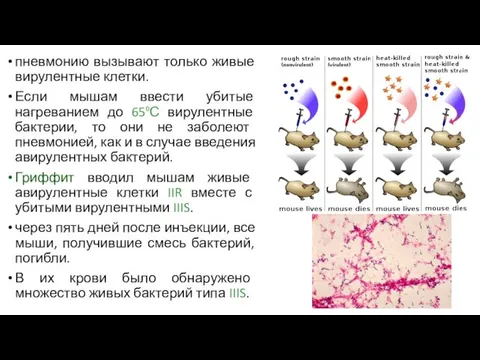
пневмонию вызывают только живые вирулентные клетки.
Если мышам ввести убитые нагреванием до
пневмонию вызывают только живые вирулентные клетки.
Если мышам ввести убитые нагреванием до
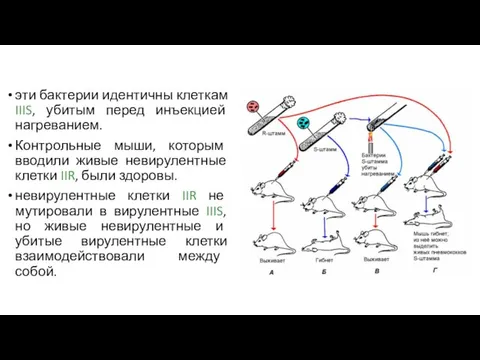
эти бактерии идентичны клеткам IIIS, убитым перед инъекцией нагреванием.
Контрольные мыши, которым
эти бактерии идентичны клеткам IIIS, убитым перед инъекцией нагреванием.
Контрольные мыши, которым

Гриффит предположил, что убитые клетки каким-то образом превращают невирулентные бактерии в
Гриффит предположил, что убитые клетки каким-то образом превращают невирулентные бактерии в

В начале 30-х годов Генри Доусон с сотрудниками показали, что трансформация
В начале 30-х годов Генри Доусон с сотрудниками показали, что трансформация

1944 г., были опубликованы результаты, полученные Эвери, Мак-Леодом и Мак-Карти.
Им удалось
1944 г., были опубликованы результаты, полученные Эвери, Мак-Леодом и Мак-Карти.
Им удалось

Сначала исследователи выращивали большие объемы (50 – 70 л) жидких культур
Сначала исследователи выращивали большие объемы (50 – 70 л) жидких культур

Эвери с коллегами решили, что трансформирующий фактор взаимодействует с клетками типа
Эвери с коллегами решили, что трансформирующий фактор взаимодействует с клетками типа
трансформация происходит у Hemophilus influenzae, Bacillus subtilis, Sigella paradysenteriae, Escherichia coli.
трансформация
трансформация происходит у Hemophilus influenzae, Bacillus subtilis, Sigella paradysenteriae, Escherichia coli.
трансформация

Эксперимент Херши-Чейз
Другие доказательства роли ДНК как переносчика генетической информации были получены
Эксперимент Херши-Чейз
Другие доказательства роли ДНК как переносчика генетической информации были получены

В 1952 г. Альфред Херши и Марта Чейз показали, что в
В 1952 г. Альфред Херши и Марта Чейз показали, что в

Из предыдущих исследований было известно, что:
1. Фаги Т2 примерно на 50%
Из предыдущих исследований было известно, что:
1. Фаги Т2 примерно на 50%

Если предположить, что некоторые из молекулярных компонентов фага (ДНК или белки)
Если предположить, что некоторые из молекулярных компонентов фага (ДНК или белки)

Херши и Чейз использовали радиоизотопный метод.
Они метили ДНК радиоактивным фосфором (32Р),
Херши и Чейз использовали радиоизотопный метод.
Они метили ДНК радиоактивным фосфором (32Р),
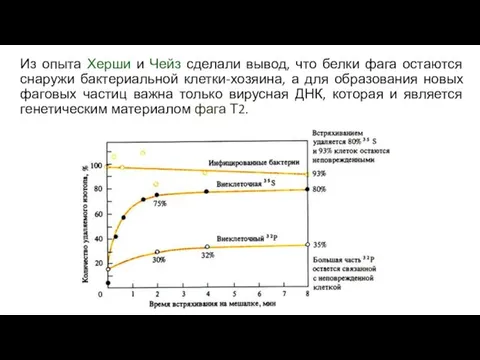
Из опыта Херши и Чейз сделали вывод, что белки фага остаются
Из опыта Херши и Чейз сделали вывод, что белки фага остаются
Эксперименты Херши и Чейз, а также Эвери с сотрудниками убедительно показали,
Эксперименты Херши и Чейз, а также Эвери с сотрудниками убедительно показали,

Опыты по трансфекции
1957 г. – опубликовано несколько работ, которые показали, что
Опыты по трансфекции
1957 г. – опубликовано несколько работ, которые показали, что

1960 г. – проведены сходные эксперименты с использованием очищенной фаговой ДНК.
Заражение
1960 г. – проведены сходные эксперименты с использованием очищенной фаговой ДНК.
Заражение

Прямые и непрямые доказательства значения ДНК у эукариот
В 50-х годах ХХ
Прямые и непрямые доказательства значения ДНК у эукариот
В 50-х годах ХХ

Непрямое доказательство: распределение ДНК
ДНК локализована в клетке там, где определяются определенные
Непрямое доказательство: распределение ДНК
ДНК локализована в клетке там, где определяются определенные

Очевидна тесная корреляция между количеством хромосом и содержанием ДНК в клетках.
Однако
Очевидна тесная корреляция между количеством хромосом и содержанием ДНК в клетках.
Однако

Непрямое доказательство: мутагенез
Ультрафиолетовый свет (УФ) – один из агентов, индуцирующих мутации
Непрямое доказательство: мутагенез
Ультрафиолетовый свет (УФ) – один из агентов, индуцирующих мутации

УФ обладает максимальным мутагенным действием при длине волны 260 нанометров (нм).
Наиболее
УФ обладает максимальным мутагенным действием при длине волны 260 нанометров (нм).
Наиболее

Прямое доказательство:
анализ рекомбинантных ДНК
доказательства этой гипотезы были получены с помощью метода
Прямое доказательство:
анализ рекомбинантных ДНК
доказательства этой гипотезы были получены с помощью метода

в оплодотворенные мышиные яйцеклетки с помощью микроинъекций можно ввести ген человеческого
в оплодотворенные мышиные яйцеклетки с помощью микроинъекций можно ввести ген человеческого

Позже в оплодотворенные мышиные яйцеклетки ввели ген rat, кодирующий гормон роста.
Около
Позже в оплодотворенные мышиные яйцеклетки ввели ген rat, кодирующий гормон роста.
Около

В эукариотических клетках происходит экспрессия генетической информации записанной в ДНК.
В эукариотических клетках происходит экспрессия генетической информации записанной в ДНК.

Структурный анализ ДНК
1953 г. Джеймс Уотсон и Фрэнсис Крик предположили, что
Структурный анализ ДНК
1953 г. Джеймс Уотсон и Фрэнсис Крик предположили, что

В распоряжении Уотсона и Крика было две группы данных:
(1) результаты
В распоряжении Уотсона и Крика было две группы данных:
(1) результаты

Химия нуклеиновых кислот
Когда Уотсон и Крик работали над своей моделью ДНК,
Химия нуклеиновых кислот
Когда Уотсон и Крик работали над своей моделью ДНК,
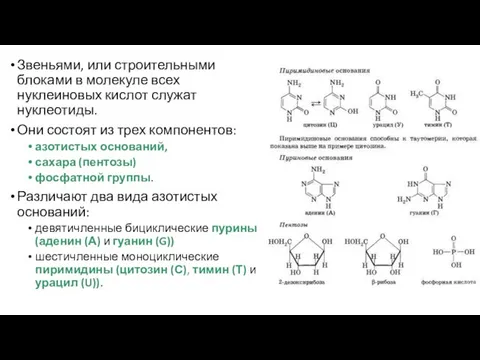
Звеньями, или строительными блоками в молекуле всех нуклеиновых кислот служат нуклеотиды.
Они
Звеньями, или строительными блоками в молекуле всех нуклеиновых кислот служат нуклеотиды.
Они
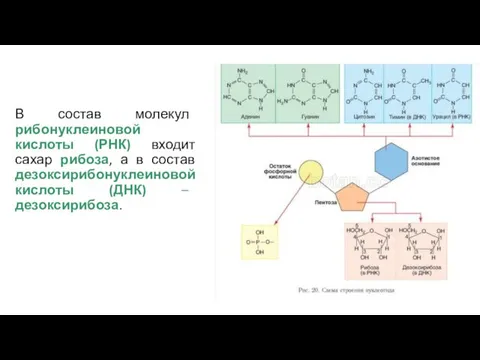
В состав молекул рибонуклеиновой кислоты (РНК) входит сахар рибоза, а в
В состав молекул рибонуклеиновой кислоты (РНК) входит сахар рибоза, а в
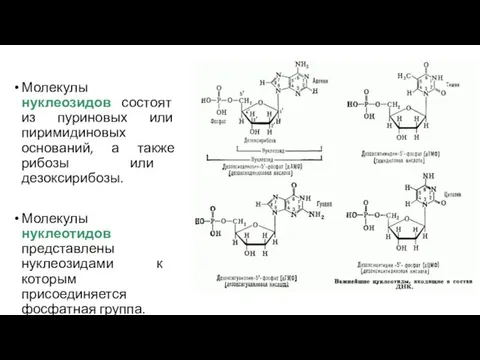
Молекулы нуклеозидов состоят из пуриновых или пиримидиновых оснований, а также рибозы
Молекулы нуклеозидов состоят из пуриновых или пиримидиновых оснований, а также рибозы
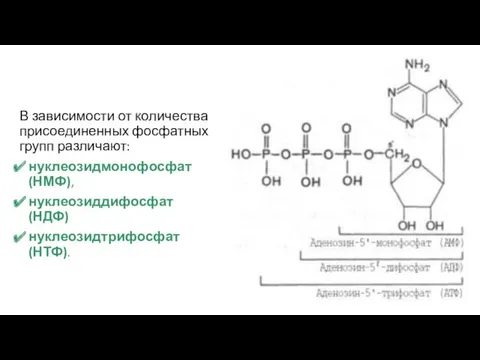
В зависимости от количества присоединенных фосфатных групп различают:
нуклеозидмонофосфат (НМФ),
нуклеозиддифосфат (НДФ)
В зависимости от количества присоединенных фосфатных групп различают:
нуклеозидмонофосфат (НМФ),
нуклеозиддифосфат (НДФ)
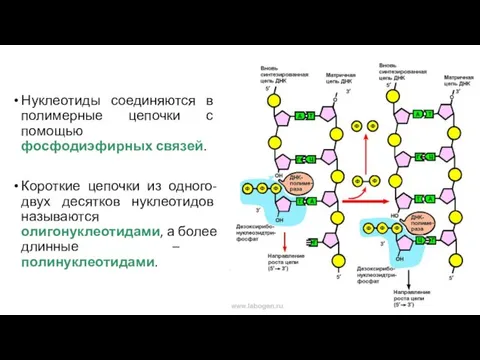
Нуклеотиды соединяются в полимерные цепочки с помощью фосфодиэфирных связей.
Короткие цепочки из
Нуклеотиды соединяются в полимерные цепочки с помощью фосфодиэфирных связей.
Короткие цепочки из

Химический состав оснований
В 1949 – 1953 гг. Эдвин Чаргафф с сотрудниками
Химический состав оснований
В 1949 – 1953 гг. Эдвин Чаргафф с сотрудниками

Если молекулы облучают рентгеновскими лучами, то часть Х-лучей, сталкиваясь с атомами,
Если молекулы облучают рентгеновскими лучами, то часть Х-лучей, сталкиваясь с атомами,

В 1950 – 1953 гг. Розалинда Франклин, работавшая в лаборатории Мориса
В 1950 – 1953 гг. Розалинда Франклин, работавшая в лаборатории Мориса

Модель Уотсона-Крика
Уотсон и Крик в 1953 г. предположили, что ДНК имеет
Модель Уотсона-Крика
Уотсон и Крик в 1953 г. предположили, что ДНК имеет

Согласно правилам Чаргаффа, молярное содержание А равно молярному содержанию Т, а
Согласно правилам Чаргаффа, молярное содержание А равно молярному содержанию Т, а
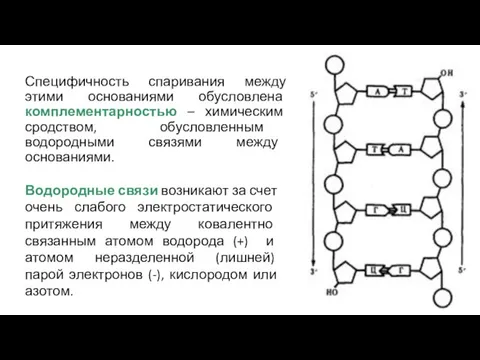
Специфичность спаривания между этими основаниями обусловлена комплементарностью – химическим сродством, обусловленным
Специфичность спаривания между этими основаниями обусловлена комплементарностью – химическим сродством, обусловленным

Более точный анализ структуры ДНК, проведенный впоследствии, выявил небольшие отклонения от
Более точный анализ структуры ДНК, проведенный впоследствии, выявил небольшие отклонения от

Уже в 1953 г. авторы заметили, что из предлагаемого ими принципа
Уже в 1953 г. авторы заметили, что из предлагаемого ими принципа

Альтернативные формы ДНК
При выделении ДНК в разных условиях можно получить различные
Альтернативные формы ДНК
При выделении ДНК в разных условиях можно получить различные

С помощью тонких методов исследовали структуру А-ДНК
преобладает в концентрированных растворах с
С помощью тонких методов исследовали структуру А-ДНК
преобладает в концентрированных растворах с

1979 г. Эндрю Ванг и Александр Рич открыли Z-конформацию
Молекула Z-ДНК представлена
1979 г. Эндрю Ванг и Александр Рич открыли Z-конформацию
Молекула Z-ДНК представлена

присутствие Z-ДНК in vivo пока не доказано
присутствие Z-ДНК in vivo пока не доказано

Структура РНК
Иногда РНК образует двуцепочечные структуры:
молекула складывается по длине и
Структура РНК
Иногда РНК образует двуцепочечные структуры:
молекула складывается по длине и

Известно три основных класса РНК, функционирующих в клетке:
рибосомная РНК (рРНК),
Известно три основных класса РНК, функционирующих в клетке:
рибосомная РНК (рРНК),
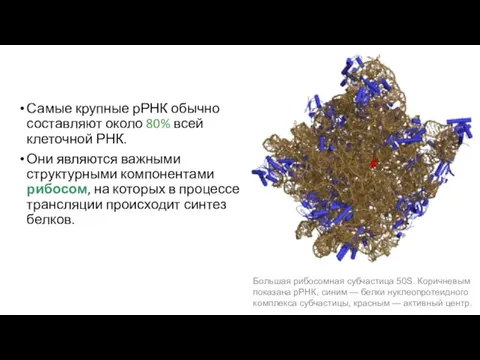
Самые крупные рРНК обычно составляют около 80% всей клеточной РНК.
Они являются
Самые крупные рРНК обычно составляют около 80% всей клеточной РНК.
Они являются
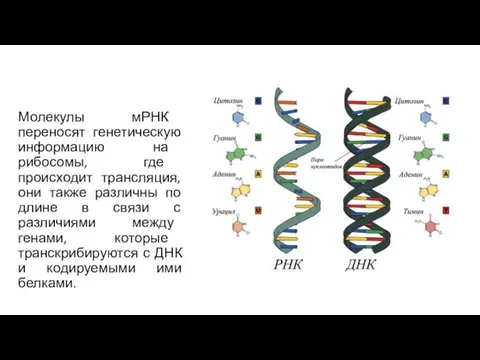
Молекулы мРНК переносят генетическую информацию на рибосомы, где происходит трансляция, они
Молекулы мРНК переносят генетическую информацию на рибосомы, где происходит трансляция, они
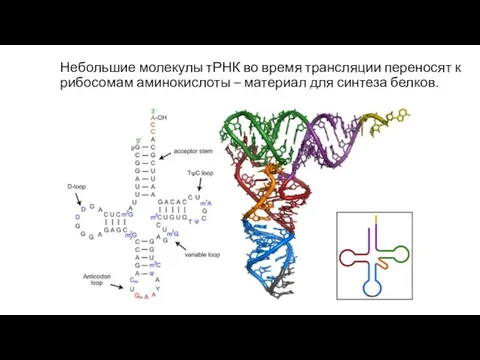
Небольшие молекулы тРНК во время трансляции переносят к рибосомам аминокислоты –
Небольшие молекулы тРНК во время трансляции переносят к рибосомам аминокислоты –

малые ядерные РНК (snRNA, или мяРНК) – участвуют в процессинге иРНК.
РНК-теломераза
малые ядерные РНК (snRNA, или мяРНК) – участвуют в процессинге иРНК.
РНК-теломераза

РНК в качестве генетического материала некоторых вирусов
1956 г. – вирус табачной
РНК в качестве генетического материала некоторых вирусов
1956 г. – вирус табачной
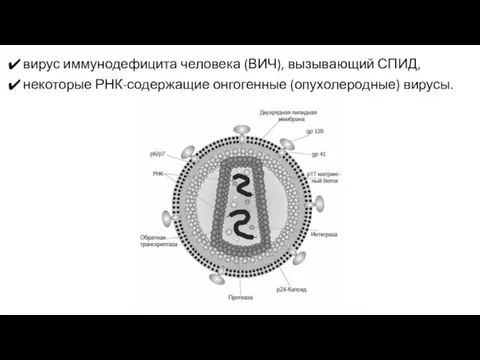
вирус иммунодефицита человека (ВИЧ), вызывающий СПИД,
некоторые РНК-содержащие онгогенные (опухолеродные) вирусы.
вирус иммунодефицита человека (ВИЧ), вызывающий СПИД,
некоторые РНК-содержащие онгогенные (опухолеродные) вирусы.

Картирование генов у бактерий и бактериофагов
Картирование генов у бактерий и бактериофагов

Бактерии и бактериофаги очень удобны для генетических исследований.
Короткий репродуктивный цикл: за
Бактерии и бактериофаги очень удобны для генетических исследований.
Короткий репродуктивный цикл: за

Мутации у бактерий и рост популяции бактериальных клеток
Растущие в культуре бактериальные
Мутации у бактерий и рост популяции бактериальных клеток
Растущие в культуре бактериальные

1943 г. Сальвадор Луриа и Макс Дельбрюк представили первые доказательства возможности
1943 г. Сальвадор Луриа и Макс Дельбрюк представили первые доказательства возможности

Бактерий выращивают в жидкой среде или в агаре на чашках Петри.
Минимальная
Бактерий выращивают в жидкой среде или в агаре на чашках Петри.
Минимальная

Для количественного анализа бактериальной культуры часть клеток переносят в свежую жидкую
Для количественного анализа бактериальной культуры часть клеток переносят в свежую жидкую

Для подсчета клеток определенный объем бактериальной культуры высевают из жидкой среды
Для подсчета клеток определенный объем бактериальной культуры высевают из жидкой среды

С разведением культуры плотность клеток падает и соответственно уменьшается число клеток,
С разведением культуры плотность клеток падает и соответственно уменьшается число клеток,

Генетическая рекомбинация у бактерий: конъюгация
1946 г. Джошуа Ледерберг и Эдвард Татум
Генетическая рекомбинация у бактерий: конъюгация
1946 г. Джошуа Ледерберг и Эдвард Татум

Ледерберг и Татум работали с различными ауксотрофными штаммами E. coli K12.
штамм
Ледерберг и Татум работали с различными ауксотрофными штаммами E. coli K12.
штамм

В эксперименте прототрофные клетки возникали с частотой 1/107 (10-7).
В эксперименте прототрофные клетки возникали с частотой 1/107 (10-7).

Бактерии F+ И F--типа
Многочисленные эксперименты, проведенные вслед за опытами Ледерберга и
Бактерии F+ И F--типа
Многочисленные эксперименты, проведенные вслед за опытами Ледерберга и

для передачи генов необходим тесный контакт двух клеток.
Бернард Дэвис доказал это
для передачи генов необходим тесный контакт двух клеток.
Бернард Дэвис доказал это

Джошуа и Эстер Ледерберги, Вильям Найес и Лукас Кавалли-Сфорца
после конъюгации
Джошуа и Эстер Ледерберги, Вильям Найес и Лукас Кавалли-Сфорца
после конъюгации
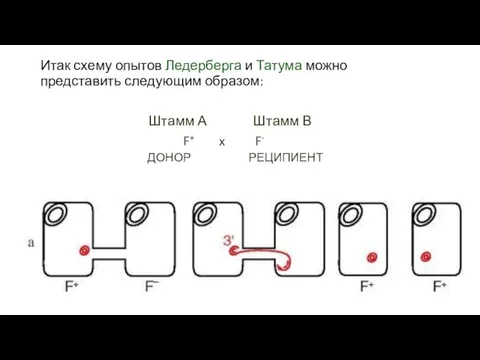
Итак схему опытов Ледерберга и Татума можно представить следующим образом:
Штамм
Итак схему опытов Ледерберга и Татума можно представить следующим образом:
Штамм
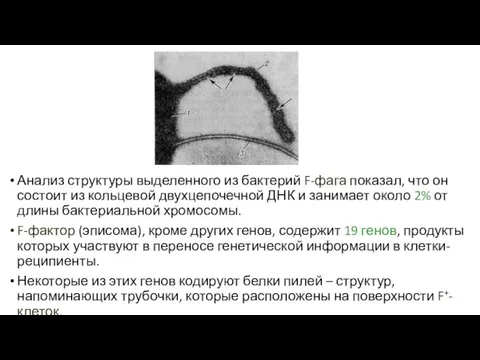
Анализ структуры выделенного из бактерий F-фага показал, что он состоит из
Анализ структуры выделенного из бактерий F-фага показал, что он состоит из
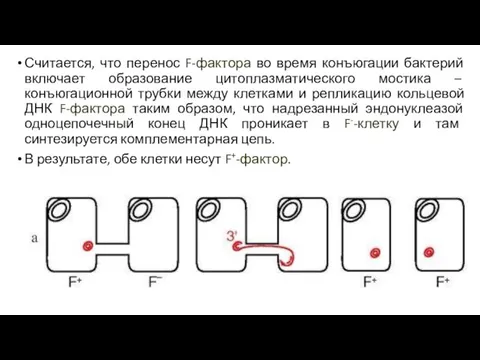
Считается, что перенос F-фактора во время конъюгации бактерий включает образование цитоплазматического
Считается, что перенос F-фактора во время конъюгации бактерий включает образование цитоплазматического

Бактериальные штаммы Hfr и хромосомное картирование
1950 г. Кавалли-Сфорца воздействовал на F+-клетки
Бактериальные штаммы Hfr и хромосомное картирование
1950 г. Кавалли-Сфорца воздействовал на F+-клетки
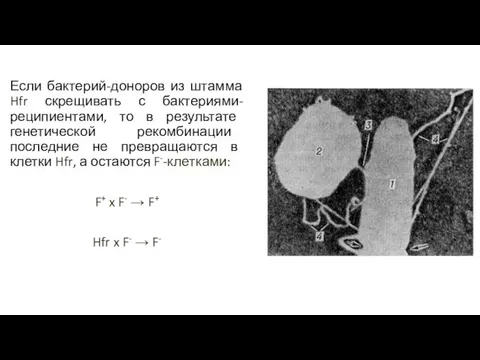
Если бактерий-доноров из штамма Hfr скрещивать с бактериями-реципиентами, то в результате
Если бактерий-доноров из штамма Hfr скрещивать с бактериями-реципиентами, то в результате

Оказалось, что такая неслучайная рекомбинация затрагивает в различных штаммах Hfr различные
Оказалось, что такая неслучайная рекомбинация затрагивает в различных штаммах Hfr различные

В середине 50-х годов ХХ века Элли Вольман и Франсуа Жакоб
В середине 50-х годов ХХ века Элли Вольман и Франсуа Жакоб

метод прерывания скрещивания.
В первые 8 минут после смешивания культур генетической рекомбинации
метод прерывания скрещивания.
В первые 8 минут после смешивания культур генетической рекомбинации

расстояние между линейно расположенными генами на хромосоме E.coli можно измерить в
расстояние между линейно расположенными генами на хромосоме E.coli можно измерить в

Сходные результаты были получены Вольман и Жакобом и на других штаммах
Сходные результаты были получены Вольман и Жакобом и на других штаммах

Основное различие касается точки начала переноса (О) и направления переноса генов
Основное различие касается точки начала переноса (О) и направления переноса генов
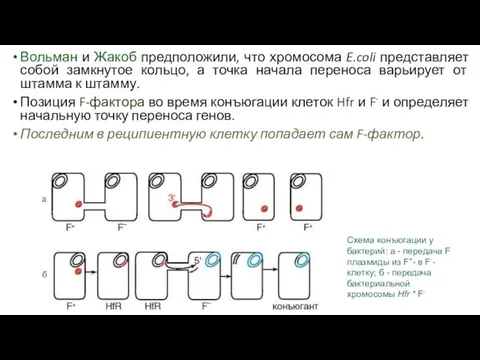
Вольман и Жакоб предположили, что хромосома E.coli представляет собой замкнутое кольцо,
Вольман и Жакоб предположили, что хромосома E.coli представляет собой замкнутое кольцо,
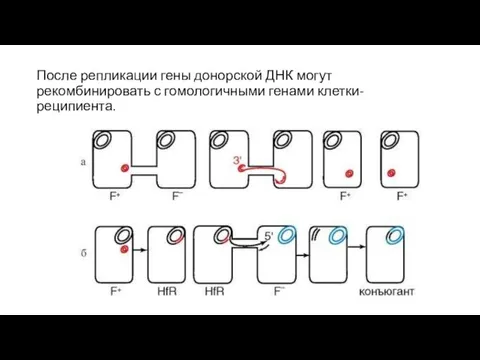
После репликации гены донорской ДНК могут рекомбинировать с гомологичными генами клетки-реципиента.
После репликации гены донорской ДНК могут рекомбинировать с гомологичными генами клетки-реципиента.

Метод прерывания скрещивания позволил картировать всю хромосому E.coli.
хромосомная карта штамма К12
Метод прерывания скрещивания позволил картировать всю хромосому E.coli.
хромосомная карта штамма К12

F’-элемент и мерозиготы
1959 г. Эдвард Адельберг в экспериментах с клетками Hfr
F’-элемент и мерозиготы
1959 г. Эдвард Адельберг в экспериментах с клетками Hfr

Присутствие бактериальных генов в составе эписомы (F-фактора) приводит к тому, что
Присутствие бактериальных генов в составе эписомы (F-фактора) приводит к тому, что

F-факторы и плазмиды
Когда экстрахромосомный F-фактор находится в цитоплазме бактериальной клетки, он
F-факторы и плазмиды
Когда экстрахромосомный F-фактор находится в цитоплазме бактериальной клетки, он

Классификация плазмид:
F-фактор – содержит гены, определяющие фертильность и ответственные за формирование
Классификация плазмид:
F-фактор – содержит гены, определяющие фертильность и ответственные за формирование

Большинство R-плазмид содержит два компонента: фактор переноса устойчивости – RTF (resistance
Большинство R-плазмид содержит два компонента: фактор переноса устойчивости – RTF (resistance

Плазмиды типа Col, в частности, ColE1, кодируют от одного до нескольких
Плазмиды типа Col, в частности, ColE1, кодируют от одного до нескольких

Бактериальная трансформация
В результате трансформации происходит генетическая рекомбинация между бактериальными клетками.
Бактериальная трансформация
В результате трансформации происходит генетическая рекомбинация между бактериальными клетками.
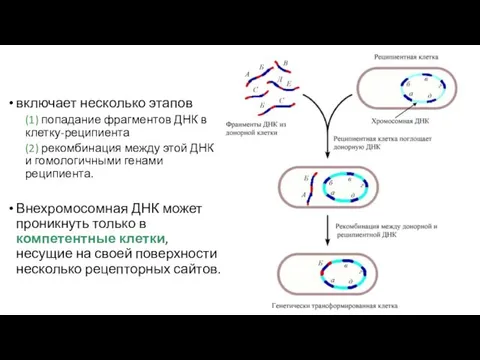
включает несколько этапов
(1) попадание фрагментов ДНК в клетку-реципиента
(2) рекомбинация между
включает несколько этапов
(1) попадание фрагментов ДНК в клетку-реципиента
(2) рекомбинация между
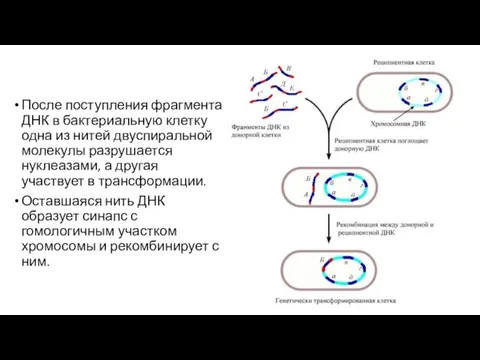
После поступления фрагмента ДНК в бактериальную клетку одна из нитей двуспиральной
После поступления фрагмента ДНК в бактериальную клетку одна из нитей двуспиральной

После интеграции чужеродной ДНК в бактериальную хромосому она несет одну исходную
После интеграции чужеродной ДНК в бактериальную хромосому она несет одну исходную

Трансформация и сцепленные гены
Для эффективности трансформации фрагмент ДНК должен содержать от
Трансформация и сцепленные гены
Для эффективности трансформации фрагмент ДНК должен содержать от

трансформация характерна для многих бактерий, включая Diplococcus pneumoniae, Hemophilus influenzae, Bacillus
трансформация характерна для многих бактерий, включая Diplococcus pneumoniae, Hemophilus influenzae, Bacillus

Генетические исследования бактериофагов
Бактериофаги, или фаги – это бактериальные вирусы, которые могут
Генетические исследования бактериофагов
Бактериофаги, или фаги – это бактериальные вирусы, которые могут
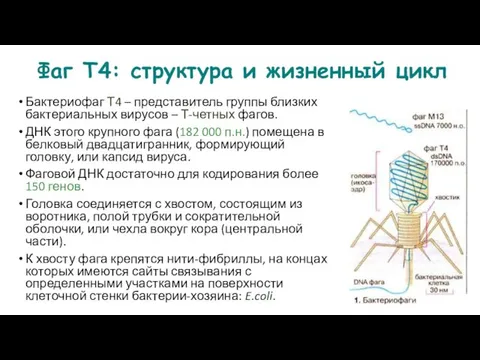
Фаг Т4: структура и жизненный цикл
Бактериофаг Т4 – представитель группы близких
Фаг Т4: структура и жизненный цикл
Бактериофаг Т4 – представитель группы близких
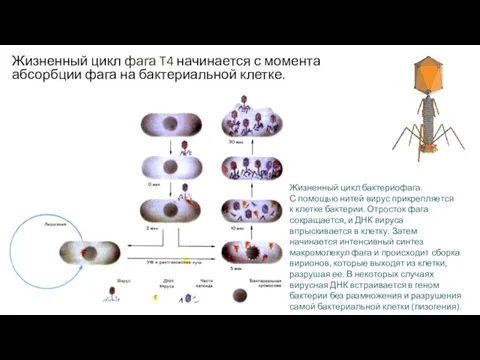
Жизненный цикл фага T4 начинается с момента абсорбции фага на бактериальной
Жизненный цикл фага T4 начинается с момента абсорбции фага на бактериальной

Сборку фаговых частиц подробно изучали Вильям Вуд, Роберт Эдгар и другие
Сборку фаговых частиц подробно изучали Вильям Вуд, Роберт Эдгар и другие
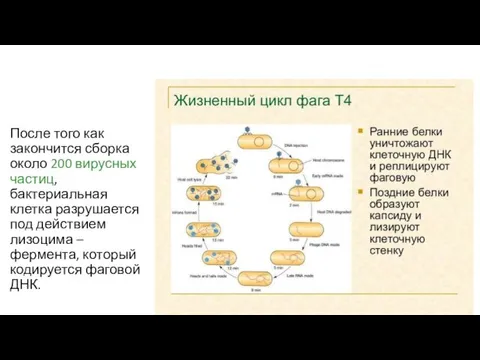
После того как закончится сборка около 200 вирусных частиц, бактериальная клетка
После того как закончится сборка около 200 вирусных частиц, бактериальная клетка

Метод бляшек
В одном мл инфицированной культуры, например, содержится более 1010 вирусных
Метод бляшек
В одном мл инфицированной культуры, например, содержится более 1010 вирусных

проводят серийное разведение инфицированной бактериальной культуры.
по 0,1 мл каждого из разведений
проводят серийное разведение инфицированной бактериальной культуры.
по 0,1 мл каждого из разведений
Лизогения
Еще в 20-х годах ХХ века стало известно, что вирус может
Лизогения
Еще в 20-х годах ХХ века стало известно, что вирус может

Вирусная ДНК интегрирует в хромосому в виде профага.
Вирусы, которые могут лизировать
Вирусная ДНК интегрирует в хромосому в виде профага.
Вирусы, которые могут лизировать

Трансдукция:
перенос бактериальной ДНК вирусом
1952 г. Нортон Зиндер и Джошуа Ледерберг
Трансдукция:
перенос бактериальной ДНК вирусом
1952 г. Нортон Зиндер и Джошуа Ледерберг

Эксперимент Зиндера-Ледерберга
Ледерберг и Зиндер смешали между собой два ауксотрофных штамма Salmonella
Эксперимент Зиндера-Ледерберга
Ледерберг и Зиндер смешали между собой два ауксотрофных штамма Salmonella

После совместной инкубации в трубке Дэвиса штаммы поместили на разные чашки
После совместной инкубации в трубке Дэвиса штаммы поместили на разные чашки

С учетом этих наблюдений, а также возможности инфекции сальмонеллы умеренными фагами,
С учетом этих наблюдений, а также возможности инфекции сальмонеллы умеренными фагами,
Природа трансдукции
трансдукция у E.coli связана с инфекцией фагами P1 и λ,
Природа трансдукции
трансдукция у E.coli связана с инфекцией фагами P1 и λ,
Иногда небольшой участок бактериальной ДНК пакуется вместе с вирусной ДНК и
Иногда небольшой участок бактериальной ДНК пакуется вместе с вирусной ДНК и
Когда фаг инъецирует в бактерию не вирусную, а бактериальную ДНК, она
Когда фаг инъецирует в бактерию не вирусную, а бактериальную ДНК, она

Трансдукционное картирование
Фрагменты бактериальной ДНК, вовлеченные в трансдукцию, достаточно велики и содержат
Трансдукционное картирование
Фрагменты бактериальной ДНК, вовлеченные в трансдукцию, достаточно велики и содержат

Межгенная рекомбинация и картирование у бактериофагов
К 1947 г. сразу несколько исследовательских
Межгенная рекомбинация и картирование у бактериофагов
К 1947 г. сразу несколько исследовательских

Мутации у бактериофагов
1946 г. Альфред Херши наблюдал необычные по морфологии бляшки
Мутации у бактериофагов
1946 г. Альфред Херши наблюдал необычные по морфологии бляшки
Мутации у бактериофагов
Другую мутацию фага Т2, известную как круг хозяев (h
Мутации у бактериофагов
Другую мутацию фага Т2, известную как круг хозяев (h

Межгенное картирование
Генетическую рекомбинацию у бактериофагов открыли во время экспериментов по одновременной
Межгенное картирование
Генетическую рекомбинацию у бактериофагов открыли во время экспериментов по одновременной
 Презентация проекта Луковое счастье.
Презентация проекта Луковое счастье. Презентация Мировой океан.
Презентация Мировой океан. Конспект урока по теме: Простые задачи на нахождение процентов
Конспект урока по теме: Простые задачи на нахождение процентов Углеводороды. Основные гомологические ряды. Типичные представители рядов
Углеводороды. Основные гомологические ряды. Типичные представители рядов Роль школьного музея в духовно-нравственном воспитании обучающихся
Роль школьного музея в духовно-нравственном воспитании обучающихся Литературная викторина Диск
Литературная викторина Диск Нормативные документы по обеспечению транспортной безопасности
Нормативные документы по обеспечению транспортной безопасности История создания МЧС.
История создания МЧС. Изгибаемые элементы. Расчет прочности нормальных сечений. Тема 7
Изгибаемые элементы. Расчет прочности нормальных сечений. Тема 7 CASE-технологии
CASE-технологии Класс Млекопитающие. 4 500 видов. Систематика
Класс Млекопитающие. 4 500 видов. Систематика Презентация к диплому
Презентация к диплому Управление временем, или тайм – менеджмент
Управление временем, или тайм – менеджмент Валюта будущего - Bitcoin
Валюта будущего - Bitcoin Лакокрасочные матариалы
Лакокрасочные матариалы Итоговый отчёт по работе: Экология
Итоговый отчёт по работе: Экология Государство Норвегия
Государство Норвегия Русский дом
Русский дом Углеводороды. Алкены
Углеводороды. Алкены Общая характеристика способов и средств наблюдения
Общая характеристика способов и средств наблюдения Проект 6
Проект 6 Состав штукатурных работ
Состав штукатурных работ Презентация Путешествие в страну Мойдодыра
Презентация Путешествие в страну Мойдодыра Geographical situation, climate, government of Great Britain
Geographical situation, climate, government of Great Britain Cистемная красная волчанка
Cистемная красная волчанка Псориаз. Красный плоский лишай
Псориаз. Красный плоский лишай Паспорт проекта ПСР (шаблон), рекомендации по выбору ПСР проектов
Паспорт проекта ПСР (шаблон), рекомендации по выбору ПСР проектов Серия бесед по Евангелию от Матфея
Серия бесед по Евангелию от Матфея