- Антиоксидантная защита мозга

Содержание
- 2. Особенности окислительного метаболизма мозга Высокий уровень потребления кислорода Большое количество липидов с ненасыщеными жирнокислотными радикалами Насыщенность
- 3. БАЛАНС АФК В ЖИВЫХ КЛЕТКАХ ТУШЕНИЕ АФК ГЕНЕРАЦИЯ АФК Дыхательная цепь митохондрий, NADPH-оксидаза нейтрофилов, микросомальное окисление,
- 4. Нейроденеративные процессы Старение АФК AO система ДЕФИЦИТ антиокси-дантов Метаболические нарушения Токсиканты Тяжелые металлы ОКИСЛИТЕЛЬНЫЙ С Т
- 6. Антиоксидантные ферменты и низкомолекулярные антиоксиданты Супероксиддисмутаза (разные формы содержат Cu/Zn и Mn): О2- ∙ + О2-
- 7. РЕГУЛЯЦИЯ
- 8. Убиквити-нилирование и деградация поврежденных молекул белка
- 9. Контроль уровня АФК клетками глии Соотношение глия/нейроны растет в онтогенезе от 0,2 до 1,6 (человек) Соотношение
- 10. Роль каталазы Н202 Н202 Время Контрольная культура Knock-out глиальных клеток Glu-peroxidase -/- + BSO (ингибитор глу-пероксидазы)
- 11. GSH γ-L-glutamyl-L-cysteinylglycine В клетках млекопитающих концентрация от 1 - 10 мМ В мозге ~ 1 -
- 12. Функции GSH в клетках Антиоксидантная - прямое взаимодействие с радикалами в неэнзиматических реакциях (Saer et al.,1990;
- 13. Способы изменения содержания глутатиона в клетках in vitro - GSH CDNB (chloro-2,4- dinitrobenzene) цитозоль+ митохондрии DEM
- 14. Истощение цитозольного и митохондриального пулов GSH при помощи CDNB приводит к увеличению генерации митохондриальных АФК, снижает
- 15. Этанол
- 16. Гомоцистеин (ГЦ) представляет собой серосодержащую аминокислоту история исследований, связанных с определением гомоцистеина, начинается с 1932 г.,
- 17. ГОМОЦИСТЕИН КАК ФАКТОР РИСКА ОКИСЛИТЕЛЬНОГО СТРЕССА МОЗГА И СЕРДЕЧНО-СОСУДИСТОЙ СИСТЕМЫ ГОМОЦИСТЕИН ГОМОЦИСТЕИНОВАЯ КИСЛОТА
- 18. Причины и следствия повышения уровня гомоцистеина в плазме крови Развитие седечно-сосудистых патологий Тромбообразование (риск тромбоэмболии увеличивается
- 20. Молекулярные последствия гипергомоцистеинемии Интенсификация метилирования нуклеиновых кислот, белков и фосфолипидов Повышенный внутриклеточный уровень свободных радикалов ---------------------
- 21. In vitro
- 22. PI (пропидий иодид ) λex=485 нм, λem=610нм (оценка смертности) DCFH-DA (2,7 – дихлордигидрофлуоресцеин) λexc=485 нм, λem=530нм
- 23. Действие ГЦ и ГЦК на глутаматные рецепторы нейронов in vitro Кальций АФК
- 24. ГЦ и ГЦК способны взаимодействовать как с ионотропными, так и с метаботропными глутаматными рецепторами AIDA –
- 25. In vivo
- 26. Experimental protocol ± Treatment with possible protector Pregnant rat Pups Methionine with drinking water 1 g/kg
- 27. Модель пренатальной гипергомоцистеинемии
- 28. Content of HC in the blood of rats under methionine over-loading Control 8-13 mkM Methionine overload
- 29. Определение чувствительности глутаматных рецепторов к лигандам У животных, получавших метионин, наблюдается тенденция к снижению чувствительности глутаматных
- 30. В случае активации нейронов при инкубации с HC или HCA ответ нейронов реализуется через не-NMDA глутаматные
- 31. При обработке данных использовали специальную программу, которая позволяет оценить следующие параметры: 1) время от начала движения
- 32. Анализ результата теста Морриса Оценивали пространственную ориентацию животных: в первый день эксперимента осуществляется претренинг животных во
- 33. Антиоксидантные ферменты и низкомолекулярные антиоксиданты Супероксиддисмутаза (разные формы содержат Cu/Zn и Mn): О2- ∙ + О2-
- 40. Парадоксальное увеличение генерации АФК в нейронах в ответ на увеличение содержания GSH может быть связано с
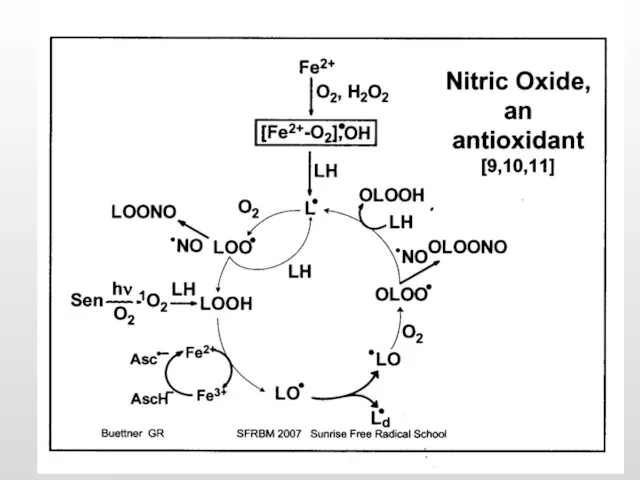
- 41. Развитие окислительного стресса в нейроне. Регистрация АФК методом проточной цитометрии +2Н+ Н2О2 + О2 О2 +
- 42. Влияние 100 µM уабаина на внутрикеточный уровень АФК в грануляных клетках мозжечка ROS signal PI versus
- 43. Изменение содержания АФК в нейронах в условиях активации ионотропных NMDA-рецепторов и метаботропных рецепторов I(3-HPG) и III(ACPD)
- 44. Effect of Glutamate Ligands on Na/K-ATPase Effect of Glutamate Ligands on Na/K-ATPase 3-HPG NMDA ACPD
- 45. D Aktivated PkB, % Нейробластома SH-SY5Y Р 14-3-3 АПОПТОЗ Выход цит С из митохондрий в цитоплазму
- 46. Na-насос в нейронах регулирует активность МАР киназы Контроль Уабаин Уабаин+D-AP5
- 47. При инкубации нейронов с уабаином рост МАР киназы зависит от активности NMDA-рецепторов и ионов кальция
- 48. Активация нейрональной MAPK уабаином требует активного состояния NMDA-рецепторов и реализуется при участии ионов Са и АФК
- 49. Корреляция между ингибированием Na/K-АТФазы и активацией МАРК уабаином
- 50. Участие Na/K-ATФазы в регуляции апоптоза АФК ПkC MAP Kиназа Na+ K+ Ca2+ α 2+3 NMDA IP3K
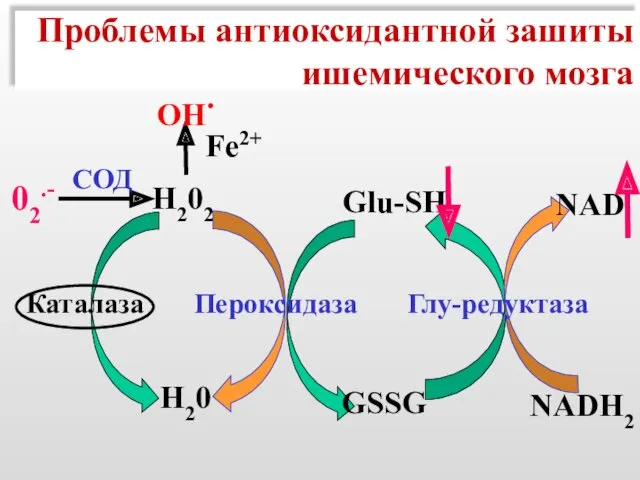
- 51. Проблемы антиоксидантной зашиты ишемического мозга Н202 Fe2+ OH. Н20 Glu-SH NAD NADH2 GSSG Каталаза Пероксидаза Глу-редуктаза
- 52. Проблемы антиоксидантной зашиты ишемического мозга Н202 Fe2+ OH. Н20 Glu-SH NAD NADH2 GSSG Каталаза Пероксидаза Глу-редуктаза
- 54. Скачать презентацию

Особенности окислительного метаболизма мозга
Высокий уровень потребления кислорода
Большое количество липидов с ненасыщеными
Особенности окислительного метаболизма мозга
Высокий уровень потребления кислорода
Большое количество липидов с ненасыщеными
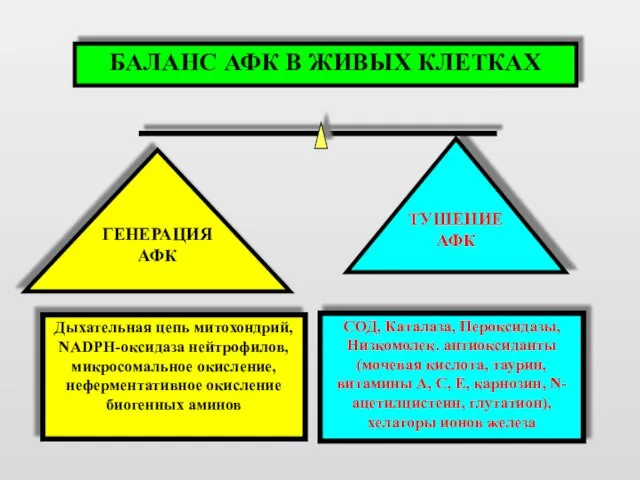
БАЛАНС АФК В ЖИВЫХ КЛЕТКАХ
ТУШЕНИЕ
АФК
ГЕНЕРАЦИЯ
АФК
Дыхательная цепь митохондрий, NADPH-оксидаза нейтрофилов, микросомальное
БАЛАНС АФК В ЖИВЫХ КЛЕТКАХ
ТУШЕНИЕ
АФК
ГЕНЕРАЦИЯ
АФК
Дыхательная цепь митохондрий, NADPH-оксидаза нейтрофилов, микросомальное
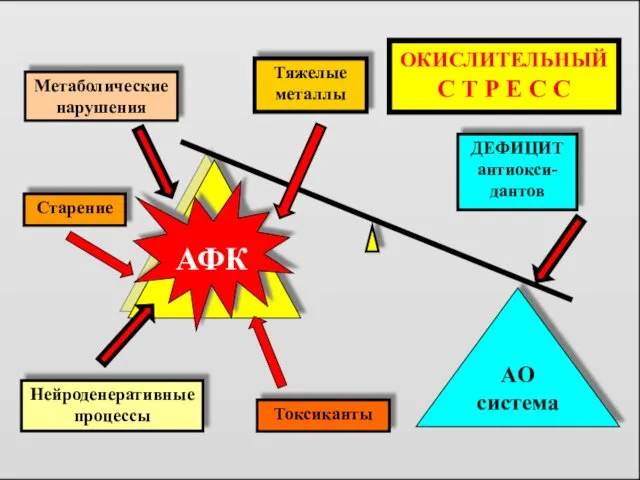
Нейроденеративные процессы
Старение
АФК
AO
система
ДЕФИЦИТ антиокси-дантов
Метаболические нарушения
Токсиканты
Тяжелые металлы
ОКИСЛИТЕЛЬНЫЙ
С Т Р Е С С
Нейроденеративные процессы
Старение
АФК
AO
система
ДЕФИЦИТ антиокси-дантов
Метаболические нарушения
Токсиканты
Тяжелые металлы
ОКИСЛИТЕЛЬНЫЙ
С Т Р Е С С


Антиоксидантные ферменты и низкомолекулярные антиоксиданты
Супероксиддисмутаза (разные формы содержат Cu/Zn и Mn):
О2-
Антиоксидантные ферменты и низкомолекулярные антиоксиданты
Супероксиддисмутаза (разные формы содержат Cu/Zn и Mn):
О2-
РЕГУЛЯЦИЯ
РЕГУЛЯЦИЯ
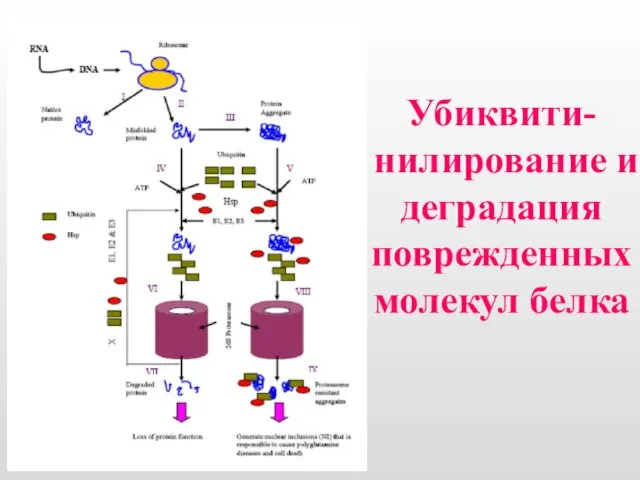
Убиквити-нилирование и деградация поврежденных молекул белка
Убиквити-нилирование и деградация поврежденных молекул белка

Контроль уровня АФК клетками глии
Соотношение глия/нейроны растет в онтогенезе от 0,2
Контроль уровня АФК клетками глии
Соотношение глия/нейроны растет в онтогенезе от 0,2
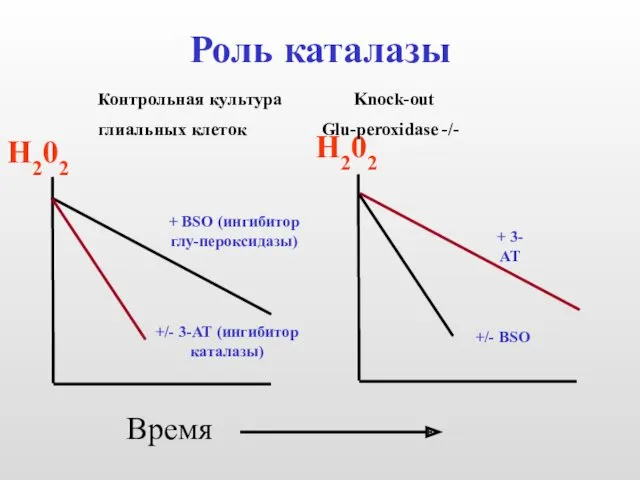
Роль каталазы
Н202
Н202
Время
Контрольная культура Knock-out
глиальных клеток Glu-peroxidase -/-
+ BSO (ингибитор
глу-пероксидазы)
+/- 3-АТ (ингибитор
каталазы)
+
Роль каталазы
Н202
Н202
Время
Контрольная культура Knock-out
глиальных клеток Glu-peroxidase -/-
+ BSO (ингибитор
глу-пероксидазы)
+/- 3-АТ (ингибитор
каталазы)
+

GSH
γ-L-glutamyl-L-cysteinylglycine
В клетках млекопитающих концентрация от 1 - 10 мМ
В мозге ~
GSH
γ-L-glutamyl-L-cysteinylglycine
В клетках млекопитающих концентрация от 1 - 10 мМ
В мозге ~

Функции GSH в клетках
Антиоксидантная - прямое взаимодействие с радикалами в неэнзиматических
Функции GSH в клетках
Антиоксидантная - прямое взаимодействие с радикалами в неэнзиматических

Способы изменения содержания глутатиона в клетках in vitro
- GSH
CDNB (chloro-2,4-
Способы изменения содержания глутатиона в клетках in vitro
- GSH
CDNB (chloro-2,4-
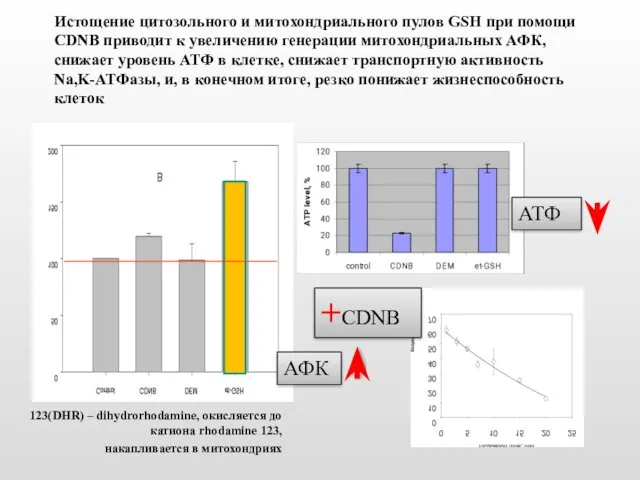
Истощение цитозольного и митохондриального пулов GSH при помощи CDNB приводит к
Истощение цитозольного и митохондриального пулов GSH при помощи CDNB приводит к
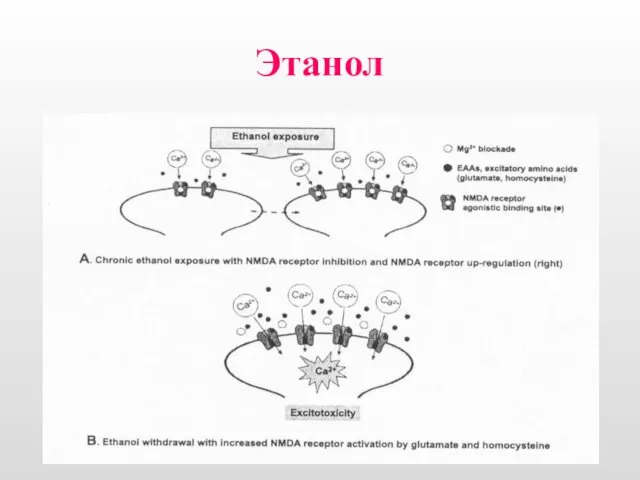
Этанол
Этанол

Гомоцистеин (ГЦ) представляет собой серосодержащую аминокислоту
история исследований, связанных с определением гомоцистеина,
Гомоцистеин (ГЦ) представляет собой серосодержащую аминокислоту
история исследований, связанных с определением гомоцистеина,
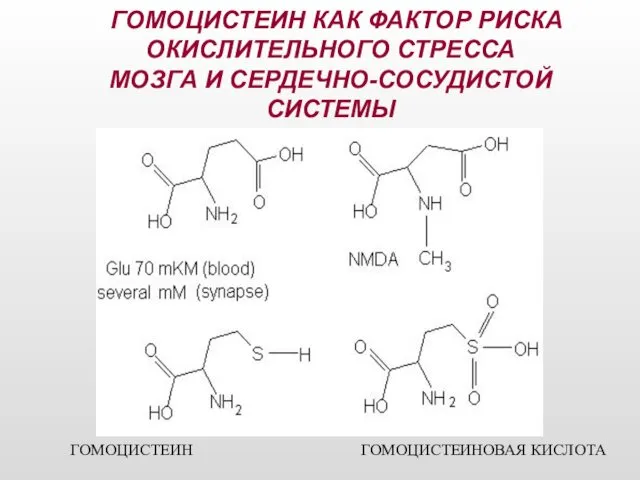
ГОМОЦИСТЕИН КАК ФАКТОР РИСКА ОКИСЛИТЕЛЬНОГО СТРЕССА
МОЗГА И СЕРДЕЧНО-СОСУДИСТОЙ СИСТЕМЫ
ГОМОЦИСТЕИН
ГОМОЦИСТЕИН КАК ФАКТОР РИСКА ОКИСЛИТЕЛЬНОГО СТРЕССА
МОЗГА И СЕРДЕЧНО-СОСУДИСТОЙ СИСТЕМЫ
ГОМОЦИСТЕИН

Причины и следствия повышения уровня гомоцистеина в плазме крови
Развитие седечно-сосудистых патологий
Тромбообразование
Причины и следствия повышения уровня гомоцистеина в плазме крови
Развитие седечно-сосудистых патологий
Тромбообразование


Молекулярные последствия гипергомоцистеинемии
Интенсификация метилирования нуклеиновых кислот, белков и фосфолипидов
Повышенный внутриклеточный уровень
Молекулярные последствия гипергомоцистеинемии
Интенсификация метилирования нуклеиновых кислот, белков и фосфолипидов
Повышенный внутриклеточный уровень

In vitro
In vitro

PI (пропидий иодид ) λex=485 нм, λem=610нм (оценка смертности)
DCFH-DA (2,7
PI (пропидий иодид ) λex=485 нм, λem=610нм (оценка смертности)
DCFH-DA (2,7
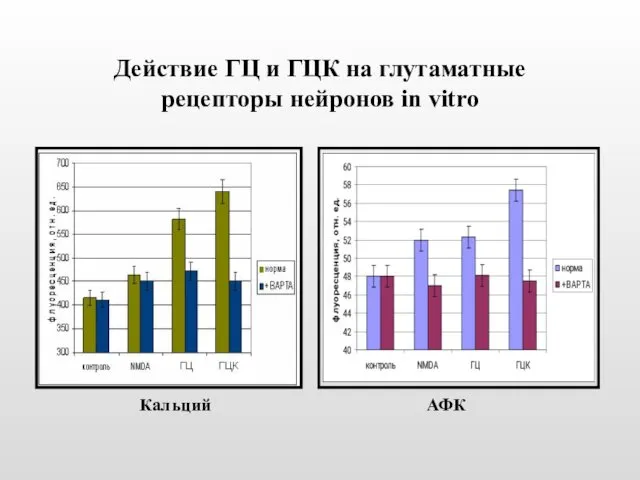
Действие ГЦ и ГЦК на глутаматные рецепторы нейронов in vitro
Кальций
АФК
Действие ГЦ и ГЦК на глутаматные рецепторы нейронов in vitro
Кальций
АФК
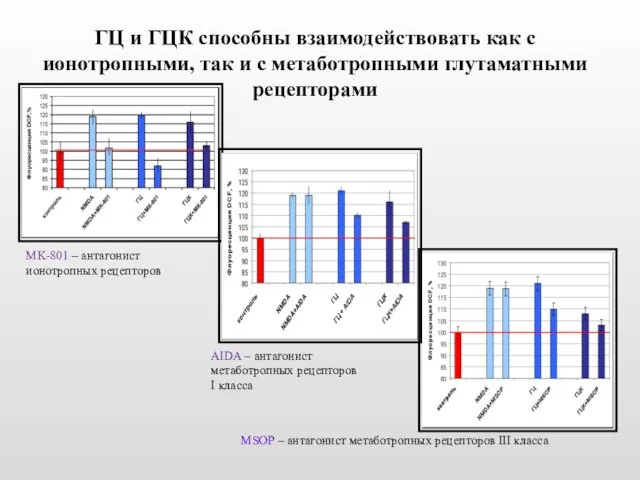
ГЦ и ГЦК способны взаимодействовать как с ионотропными, так и с
ГЦ и ГЦК способны взаимодействовать как с ионотропными, так и с

In vivo
In vivo
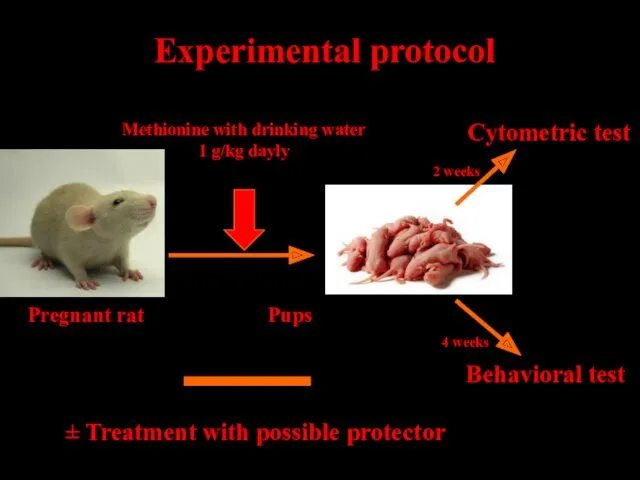
Experimental protocol
± Treatment with possible protector
Pregnant rat Pups
Methionine with drinking
Experimental protocol
± Treatment with possible protector
Pregnant rat Pups
Methionine with drinking
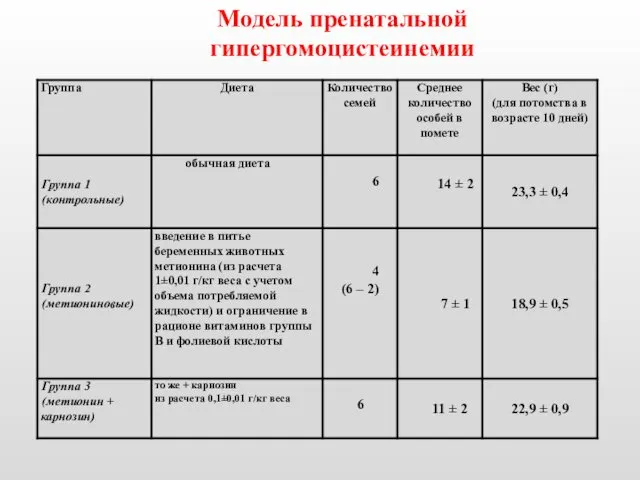
Модель пренатальной гипергомоцистеинемии
Модель пренатальной гипергомоцистеинемии

Content of HC in the blood of rats under methionine
over-loading
Content of HC in the blood of rats under methionine over-loading
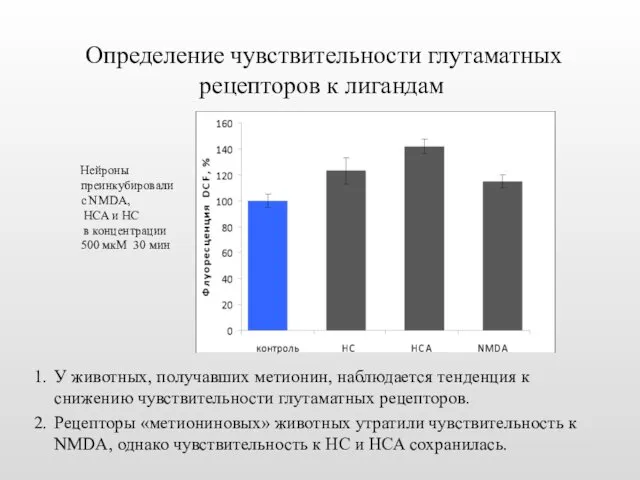
Определение чувствительности глутаматных рецепторов к лигандам
У животных, получавших метионин, наблюдается
Определение чувствительности глутаматных рецепторов к лигандам
У животных, получавших метионин, наблюдается
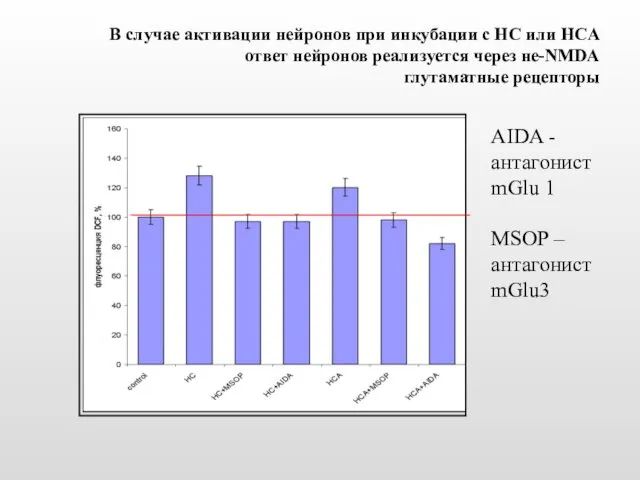
В случае активации нейронов при инкубации с HC или HCA
ответ
В случае активации нейронов при инкубации с HC или HCA ответ

При обработке данных использовали специальную программу, которая позволяет оценить следующие параметры:
1)
При обработке данных использовали специальную программу, которая позволяет оценить следующие параметры:
1)
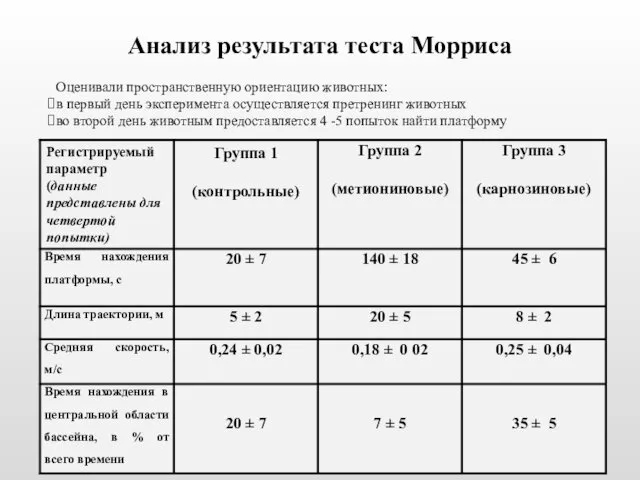
Анализ результата теста Морриса
Оценивали пространственную ориентацию животных:
в первый день эксперимента осуществляется
Анализ результата теста Морриса
Оценивали пространственную ориентацию животных:
в первый день эксперимента осуществляется

Антиоксидантные ферменты и низкомолекулярные антиоксиданты
Супероксиддисмутаза (разные формы содержат Cu/Zn и Mn):
О2-
Антиоксидантные ферменты и низкомолекулярные антиоксиданты
Супероксиддисмутаза (разные формы содержат Cu/Zn и Mn):
О2-



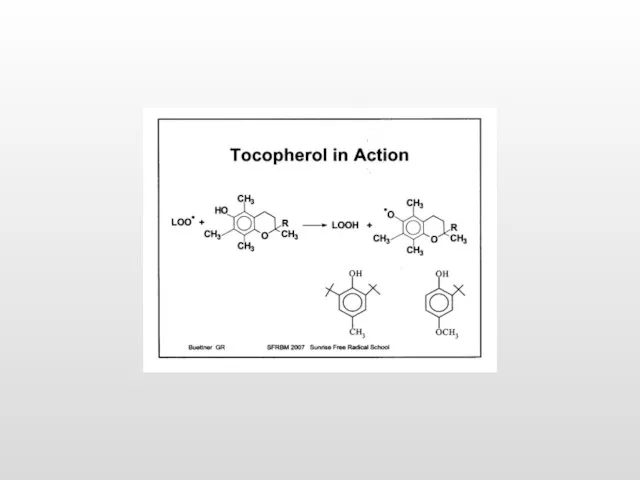


Парадоксальное увеличение генерации АФК в нейронах в ответ на увеличение содержания
Парадоксальное увеличение генерации АФК в нейронах в ответ на увеличение содержания
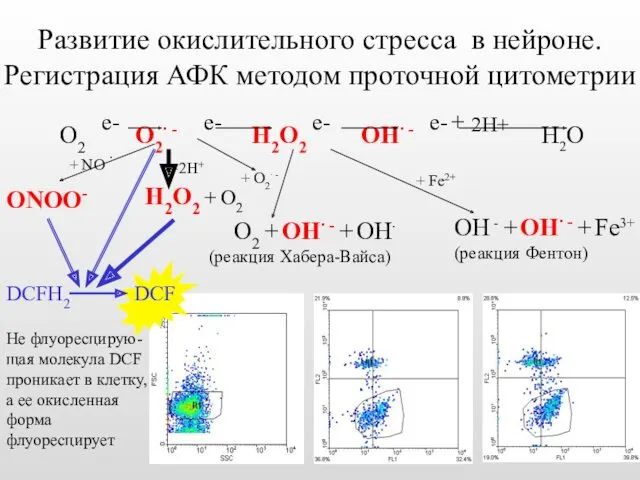
Развитие окислительного стресса в нейроне. Регистрация АФК методом проточной цитометрии
+2Н+
Н2О2 +
Развитие окислительного стресса в нейроне. Регистрация АФК методом проточной цитометрии
+2Н+
Н2О2 +
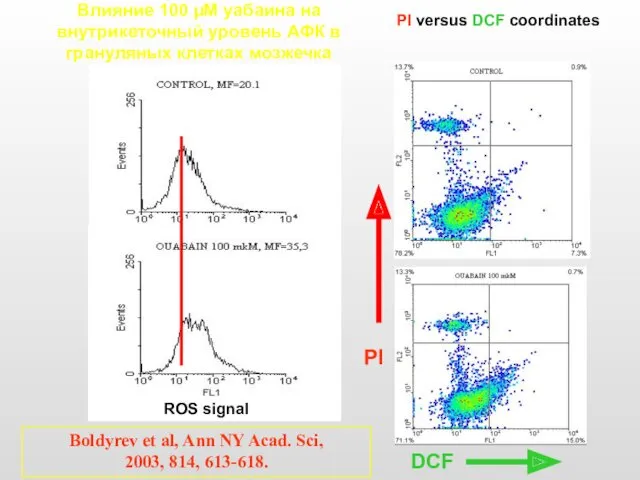
Влияние 100 µM уабаина на внутрикеточный уровень АФК в грануляных клетках
Влияние 100 µM уабаина на внутрикеточный уровень АФК в грануляных клетках
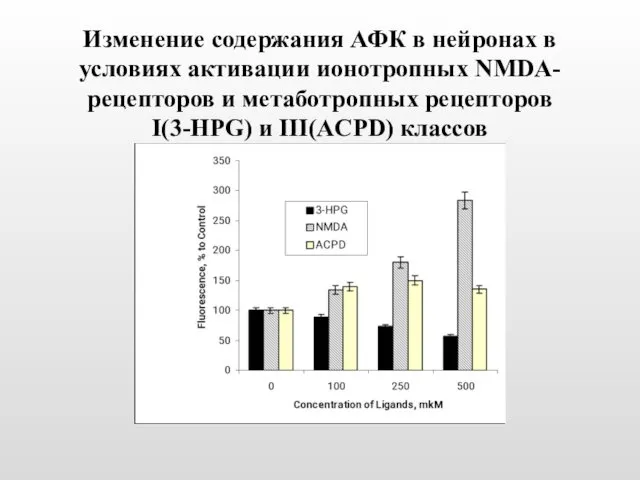
Изменение содержания АФК в нейронах в условиях активации ионотропных NMDA-рецепторов и
Изменение содержания АФК в нейронах в условиях активации ионотропных NMDA-рецепторов и
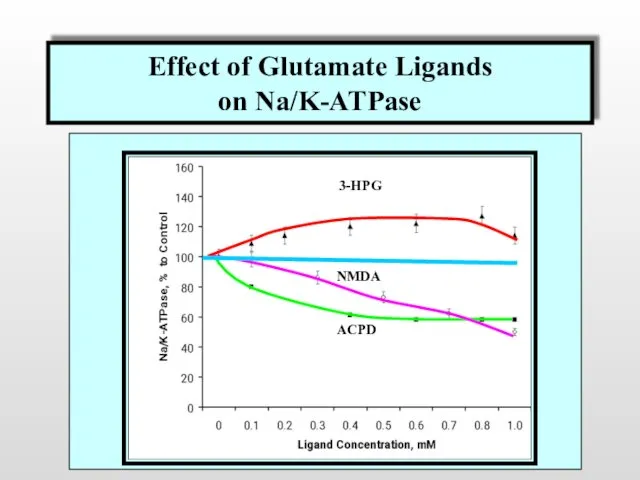
Effect of Glutamate Ligands
on Na/K-ATPase
Effect of Glutamate Ligands
on Na/K-ATPase
3-HPG
NMDA
ACPD
Effect of Glutamate Ligands
on Na/K-ATPase
Effect of Glutamate Ligands
on Na/K-ATPase
3-HPG
NMDA
ACPD
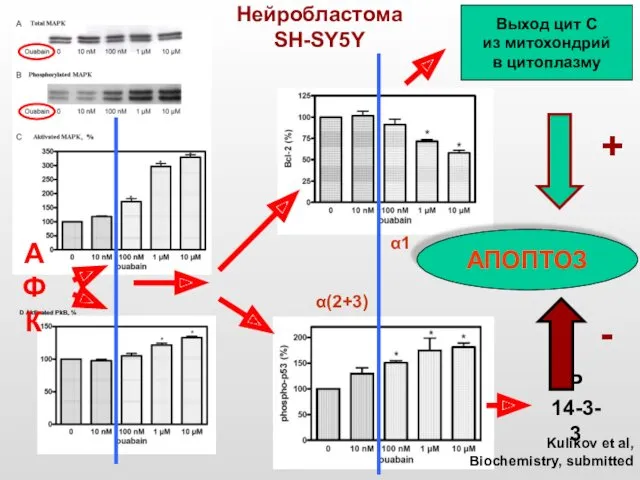
D Aktivated PkB, %
Нейробластома
SH-SY5Y
Р 14-3-3
АПОПТОЗ
Выход цит С
из митохондрий
D Aktivated PkB, %
Нейробластома
SH-SY5Y
Р 14-3-3
АПОПТОЗ
Выход цит С
из митохондрий
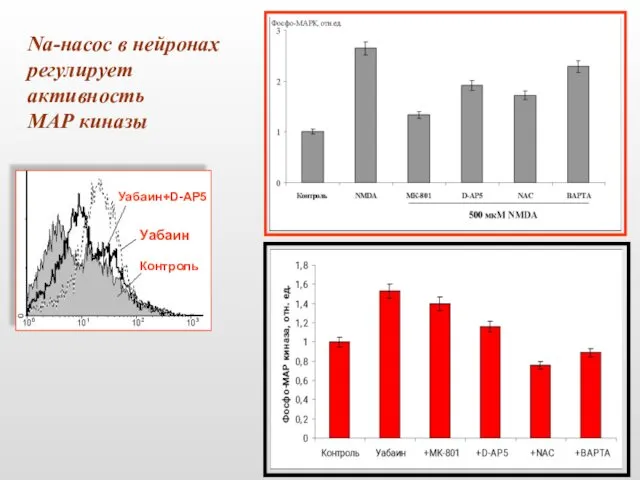
Na-насос в нейронах
регулирует
активность
МАР киназы
Контроль
Уабаин
Уабаин+D-AP5
Na-насос в нейронах
регулирует
активность
МАР киназы
Контроль
Уабаин
Уабаин+D-AP5
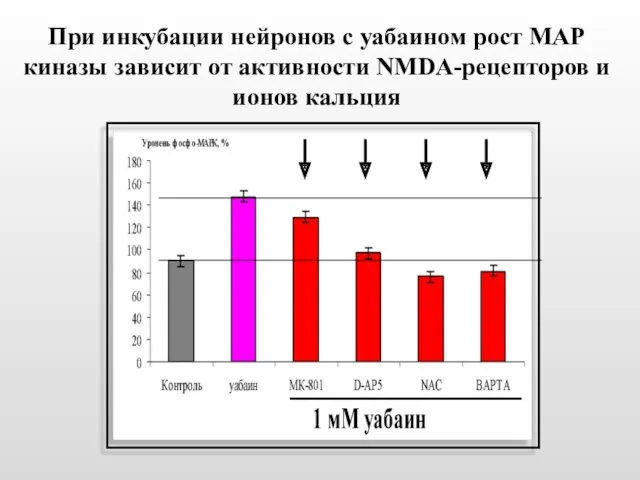
При инкубации нейронов с уабаином рост МАР киназы зависит от активности
При инкубации нейронов с уабаином рост МАР киназы зависит от активности
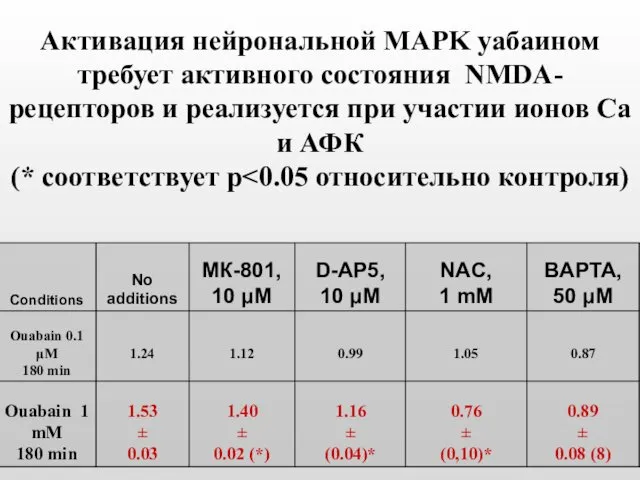
Активация нейрональной MAPK уабаином требует активного состояния NMDA-рецепторов и реализуется при
Активация нейрональной MAPK уабаином требует активного состояния NMDA-рецепторов и реализуется при
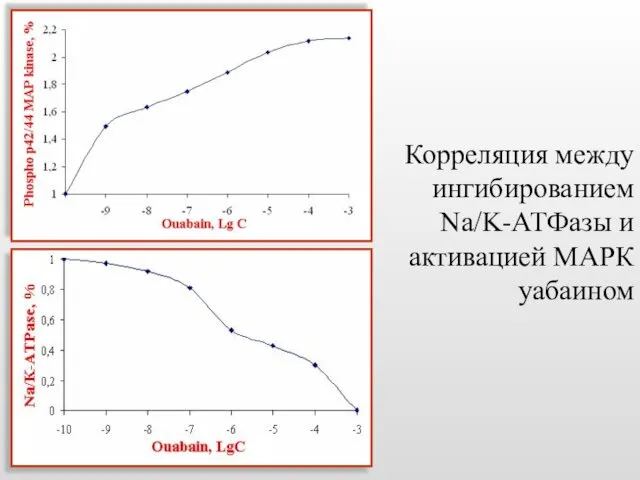
Корреляция между ингибированием Na/K-АТФазы и активацией МАРК уабаином
Корреляция между ингибированием Na/K-АТФазы и активацией МАРК уабаином
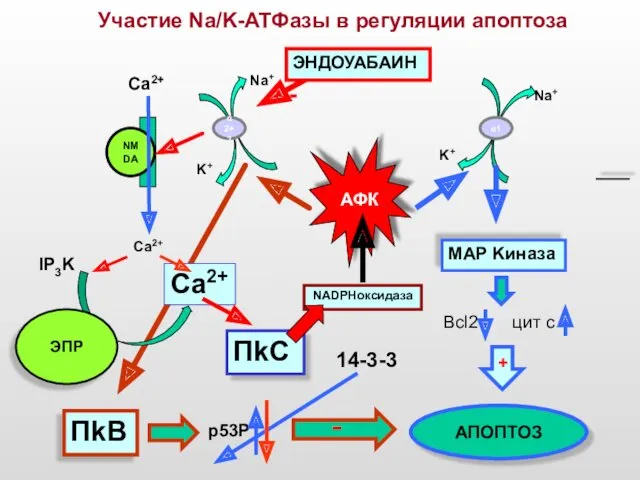
Участие Na/K-ATФазы в регуляции апоптоза
АФК
ПkC
MAP Kиназа
Na+
K+
Ca2+
α 2+3
NMDA
IP3K
ЭПР
Ca2+
NADPHоксидаза
-
ЭНДОУАБАИН
Са2+
АПОПТОЗ
Bcl2
Участие Na/K-ATФазы в регуляции апоптоза
АФК
ПkC
MAP Kиназа
Na+
K+
Ca2+
α 2+3
NMDA
IP3K
ЭПР
Ca2+
NADPHоксидаза
-
ЭНДОУАБАИН
Са2+
АПОПТОЗ
Bcl2
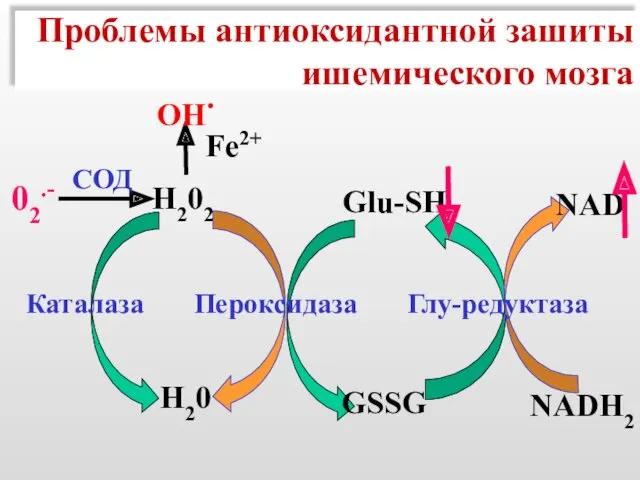
Проблемы антиоксидантной зашиты
ишемического мозга
Н202
Fe2+
OH.
Н20
Glu-SH
NAD
NADH2
GSSG
Каталаза Пероксидаза Глу-редуктаза
02.-
СОД
Проблемы антиоксидантной зашиты
ишемического мозга
Н202
Fe2+
OH.
Н20
Glu-SH
NAD
NADH2
GSSG
Каталаза Пероксидаза Глу-редуктаза
02.-
СОД
Проблемы антиоксидантной зашиты
ишемического мозга
Н202
Fe2+
OH.
Н20
Glu-SH
NAD
NADH2
GSSG
Каталаза Пероксидаза Глу-редуктаза
02.-
СОД
Проблемы антиоксидантной зашиты
ишемического мозга
Н202
Fe2+
OH.
Н20
Glu-SH
NAD
NADH2
GSSG
Каталаза Пероксидаза Глу-редуктаза
02.-
СОД
 Команди МП 8088/86. Зміст команд. Лекция 8
Команди МП 8088/86. Зміст команд. Лекция 8 Информация. Виды и свойства
Информация. Виды и свойства Концепции естественнонаучной картины мира
Концепции естественнонаучной картины мира Самопрезентация педагога
Самопрезентация педагога Урок бабочки
Урок бабочки Ритм мен өткізгіштігінің бұзылысы синдромы
Ритм мен өткізгіштігінің бұзылысы синдромы Теплофикационная установка (ТФУ)
Теплофикационная установка (ТФУ) Презентация по ПДД для детей средней группы
Презентация по ПДД для детей средней группы Цифровые системы передачи мультимедийной информации
Цифровые системы передачи мультимедийной информации Виды статистических методов контроля качества: гистограмма и расслоение
Виды статистических методов контроля качества: гистограмма и расслоение Quel temps fait-il? Leçon 8
Quel temps fait-il? Leçon 8 СНГ: проблемы и перспективы дальнейшего развития
СНГ: проблемы и перспективы дальнейшего развития Предварительное техническое предложение на поставку беспилотного вертолетного комплекса “INDELA”
Предварительное техническое предложение на поставку беспилотного вертолетного комплекса “INDELA” Интеллектуальная система охраны периметра объекта
Интеллектуальная система охраны периметра объекта Мир деревьев
Мир деревьев 19 января – Крещение Господне
19 января – Крещение Господне Архитектура компьютера. Технические средства реализации информационных процессов
Архитектура компьютера. Технические средства реализации информационных процессов Многообразие биохимических реакторов. Классификация, конструктивные особенности
Многообразие биохимических реакторов. Классификация, конструктивные особенности Понятие и сущность международного права. (Лекция 1)
Понятие и сущность международного права. (Лекция 1) Молодая, но известная... или Путешествие по Ботландии
Молодая, но известная... или Путешествие по Ботландии Составление рассказа по картине Кошка с котятами
Составление рассказа по картине Кошка с котятами Силовые трансформаторы и автотрансформаторы
Силовые трансформаторы и автотрансформаторы Моя работа - мое творчество
Моя работа - мое творчество The production base of the group of companies is located in Ukraine LTD “Novator Pallet”
The production base of the group of companies is located in Ukraine LTD “Novator Pallet” презентация на тему Игрушки Диск
презентация на тему Игрушки Диск Портфолио студента
Портфолио студента Микропроцессорлық жүйе
Микропроцессорлық жүйе Изменчивость как причина наследственных заболеваний у человека
Изменчивость как причина наследственных заболеваний у человека