- Биосинтез липидов

Содержание
- 2. Необходим для создания запасных форм. Биосинтез липидов (липогенез) начинается с биосинтеза жирных кислот. Синтез жирных кислот
- 3. Особенности биосинтеза жирных кислот Синтез жирных кислот не является простым обращением реакций β-окисления. Наиболее важными особенностями
- 4. Растущая цепь жирной кислоты удлинятся путем последовательного присоединения двухуглеродных компонентов, происходящих из ацетил-КоА. Активированным донором двухуглеродных
- 5. ЭТАПЫ БИОСИНТЕЗА ЖИРНЫХ КИСЛОТ I этап – транспорт ацетил-КоА из митохондрий в цитоплазму. Жирные кислоты синтезируются
- 6. МИТОХОНДРИЯ ЦИТОПЛАЗМА O O = C –COOH ║ + H3C – C ~ SKoA CH2 –
- 7. Цитрат образуется в митохондриальном матриксе путем конденсации ацетил-КоА и оксалоацетата. Затем диффундирует в цитоплазму, где расщепляется
- 8. Источники НАДФН2 для биосинтеза жирных кислот Оксалоацетат, образовавшийся в результате переноса ацетил-КОА в цитоплазму должен быть
- 9. МДГ 1.Оксалоацетат + НАДН*Н Малат + НАД+ 2. Малат + НАДФ+ Пируват + СО2 + НАДФ
- 10. Образовавшийся пируват легко диффундирует в митохондрии, где он карбоксилируется в оксалоацетат под действием пируваткарбоксилазы (с затратой
- 11. Таким образом, на каждую молекулу ацетил-КоА, которая переходит из митохондрий в цитозоль, образуется одна молекула НАДФН2.
- 12. II - этап – образование малонил-КоА. Является первой реакцией биосинтеза ж.к. Катализируется ферментом ацетил-КоА-карбоксилазой. Коферментом является
- 13. H3C COOH │ _ Mg2+ │ C = O + HCO3 + АТФ E – биотин
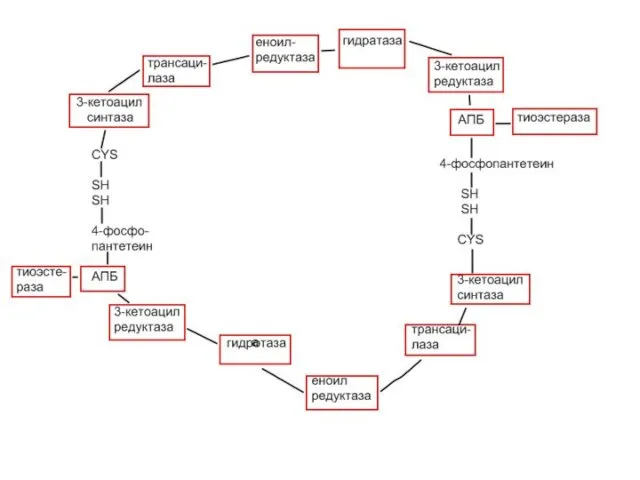
- 14. III этап – биосинтез жирных кислот. Строение синтазы жирных кислот представляет из себя полиферментный комплекс, катализирующий
- 15. HS – CYS – 1 – ФП – SH HS – ФП – 2 – CYS
- 16. Каждый мономер включает 6 индивидуальных ферментов и ацилпереносящий белок (АПБ). Cys-SH-тиоловая группа цистеина. Сульфгидрильная группа 4-фосфопантетеина
- 17. HS – CYS Е HS – ФП
- 18. СИНТЕЗ ЖИРНЫХ КИСЛОТ 1-ая реакция: ацетил-КоА под действием ацетилтрансферазы переносится на SH-группу цистеина. 2-ая реакция: малонил-КоА
- 19. 5-ая реакция: заключается в дегидратации 3-гидроксиацил-Е ферментом гидратазой, при этом образуется 2,3-ненасыщенный ацил-Е (кротонил-Е). 6-ая реакция:
- 21. Биосинтез жирных кислот HS – CYS HS – ФП Трансацилаза Ацетил – КоА (ацетилтрансацилаза) HS -
- 22. 3 – кетоацилсинтаза CO2 HS H3C – C – CH2 – C –S Ацетоацетил - фермент
- 23. HS 2,3 – Ненасыщенный ацил фермент H3C – CH = CH – C – S (кротонил
- 24. Пальмитил – Е Тиоэстераза Н2О Е Пальмитиновая кислота
- 25. Цикл синтеза повторяется. Для синтеза пальмитиновой кислоты требуется 7 таких циклов, соответственно требуется 7 остатков малонил-КоА
- 26. Суммарное уравнение синтеза пальмитиновой кислоты H3C – C(О) ~ SKOA + 7HOOC – CH2 – C(О)
- 27. Различия процессов окисления и биосинтеза
- 28. СИСТЕМА УДЛИНЕНИЯ (ЭЛОНГАЦИИ) ЖИРНЫХ КИСЛОТ Данный процесс в основном происходит в микросомах. Факторы обеспечивающие элонгацию жирных
- 29. При голодании элонгация жирных кислот затормаживается. При образовании миелиновых оболочек нервных клеток в мозгу резко усиливается
- 30. СИНТЕЗ НЕНАСЫЩЕННЫХ ЖИРНЫХ КИСЛОТ Катализируются микросомальными системами путем введения двойной связи в КоА-производных жирных кислот с
- 31. Стеароил – КоА + НАДН2 + О2 (НАДФН2) Олеоил – КоА + НАД+ + 2Н2О (НАДФ+)
- 32. Таким образом идет синтез из пальмитиновой - пальмитиолеиновой кислоты, а из стеариновой – олеиновой. У млекопитающих
- 33. Ацетил КоА Пальмитиновая кислота - 2Н + С2 Пальмитоолеиновая Стеариновая кислота кислота (16 : 1) (18
- 34. Линолевая кислота (поступает с пищей) 18 : 2 (9, 12) - 2Н Линоленовая кислота 18 :
- 35. БИОСИНТЕЗ КЕТОНОВЫХ ТЕЛ (АЦЕТОНОВЫХ) К ацетоновым телами относятся ацетоуксусная кислота, β- оксимасленая кислота, ацетон. Кетоновые тела
- 36. O O Н3С – С ~ SKоA + Н3С – С ~ SKоA HS – KoA
- 37. OH O HOOC – CH2 – C - CH2 – C ~ SKoA CH3 β -
- 38. В организме кетоновые тела выполняют важную функцию по поддержанию энергетического гомеостаза. Являются поставщиками энергии для мышц,
- 39. СН3 СOOH СН3 COOH Е С = О СН2 ТРАНСФЕРАЗА С = О СН2 энергия СН2
- 40. В тех случаях когда скорость образовании кетоновых тел превышает их потребление развиваются кетозы. Они могут быть
- 41. Патологический кетоз возникает при тяжелой форме сахарного диабета. Выделение кетоновых тел с мочой – кетонурия. Опасность
- 43. Скачать презентацию

Необходим для создания запасных форм. Биосинтез липидов (липогенез) начинается с биосинтеза
Необходим для создания запасных форм. Биосинтез липидов (липогенез) начинается с биосинтеза

Особенности биосинтеза жирных кислот
Синтез жирных кислот не является простым обращением реакций
Особенности биосинтеза жирных кислот
Синтез жирных кислот не является простым обращением реакций

Растущая цепь жирной кислоты удлинятся путем последовательного присоединения двухуглеродных компонентов, происходящих
Растущая цепь жирной кислоты удлинятся путем последовательного присоединения двухуглеродных компонентов, происходящих

ЭТАПЫ БИОСИНТЕЗА ЖИРНЫХ КИСЛОТ
I этап – транспорт ацетил-КоА из митохондрий в
ЭТАПЫ БИОСИНТЕЗА ЖИРНЫХ КИСЛОТ
I этап – транспорт ацетил-КоА из митохондрий в
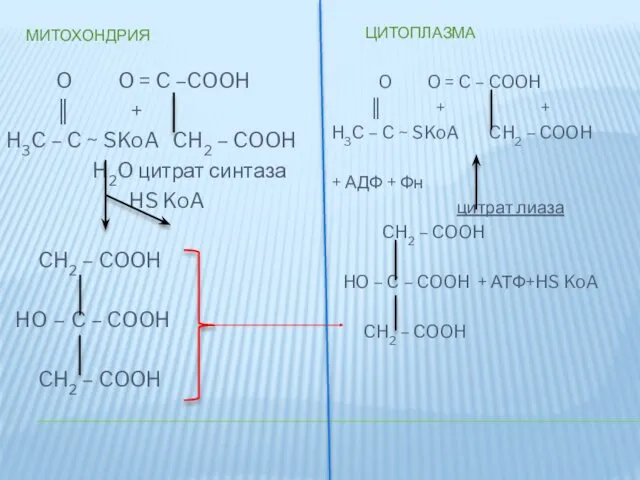
МИТОХОНДРИЯ
ЦИТОПЛАЗМА
O O = C –COOH
║ +
H3C –
МИТОХОНДРИЯ
ЦИТОПЛАЗМА
O O = C –COOH
║ +
H3C –

Цитрат образуется в митохондриальном матриксе путем конденсации ацетил-КоА и оксалоацетата. Затем
Цитрат образуется в митохондриальном матриксе путем конденсации ацетил-КоА и оксалоацетата. Затем

Источники НАДФН2 для биосинтеза жирных кислот
Оксалоацетат, образовавшийся в результате переноса ацетил-КОА
Источники НАДФН2 для биосинтеза жирных кислот
Оксалоацетат, образовавшийся в результате переноса ацетил-КОА
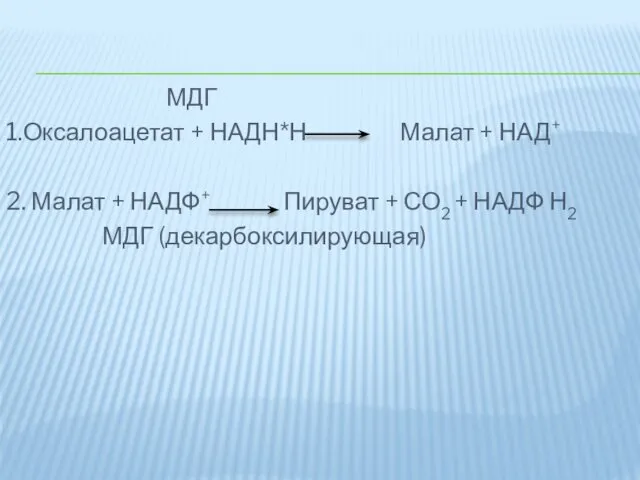
МДГ
1.Оксалоацетат + НАДН*Н Малат + НАД+
2. Малат + НАДФ+ Пируват +
1.Оксалоацетат + НАДН*Н Малат + НАД+
2. Малат + НАДФ+ Пируват +

Образовавшийся пируват легко диффундирует в митохондрии, где он карбоксилируется в оксалоацетат
Образовавшийся пируват легко диффундирует в митохондрии, где он карбоксилируется в оксалоацетат

Таким образом, на каждую молекулу ацетил-КоА, которая переходит из митохондрий в
Таким образом, на каждую молекулу ацетил-КоА, которая переходит из митохондрий в

II - этап – образование малонил-КоА.
Является первой реакцией биосинтеза ж.к. Катализируется
II - этап – образование малонил-КоА.
Является первой реакцией биосинтеза ж.к. Катализируется
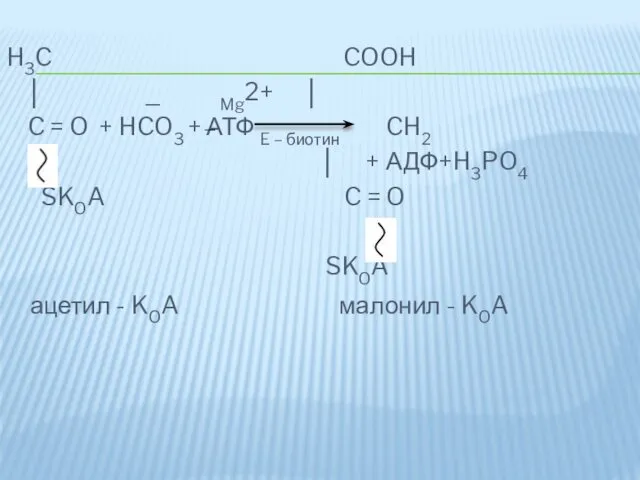
H3C COOH
│ _ Mg2+ │
C = O + HCO3
│ _ Mg2+ │
C = O + HCO3

III этап – биосинтез жирных кислот.
Строение синтазы жирных кислот представляет из
III этап – биосинтез жирных кислот.
Строение синтазы жирных кислот представляет из

HS – CYS – 1 – ФП – SH
HS – ФП
HS – ФП

Каждый мономер включает 6 индивидуальных ферментов и ацилпереносящий белок (АПБ).
Cys-SH-тиоловая группа
Каждый мономер включает 6 индивидуальных ферментов и ацилпереносящий белок (АПБ).
Cys-SH-тиоловая группа
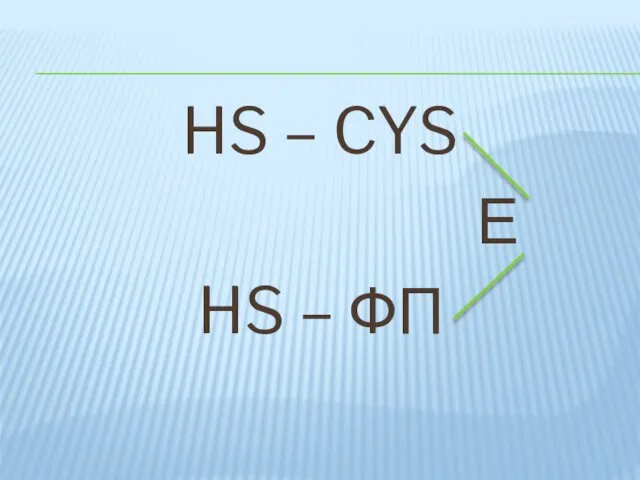
HS – CYS
Е
HS – ФП
Е
HS – ФП

СИНТЕЗ ЖИРНЫХ КИСЛОТ
1-ая реакция: ацетил-КоА под действием ацетилтрансферазы переносится на SH-группу
СИНТЕЗ ЖИРНЫХ КИСЛОТ
1-ая реакция: ацетил-КоА под действием ацетилтрансферазы переносится на SH-группу

5-ая реакция: заключается в дегидратации 3-гидроксиацил-Е ферментом гидратазой, при этом образуется
5-ая реакция: заключается в дегидратации 3-гидроксиацил-Е ферментом гидратазой, при этом образуется
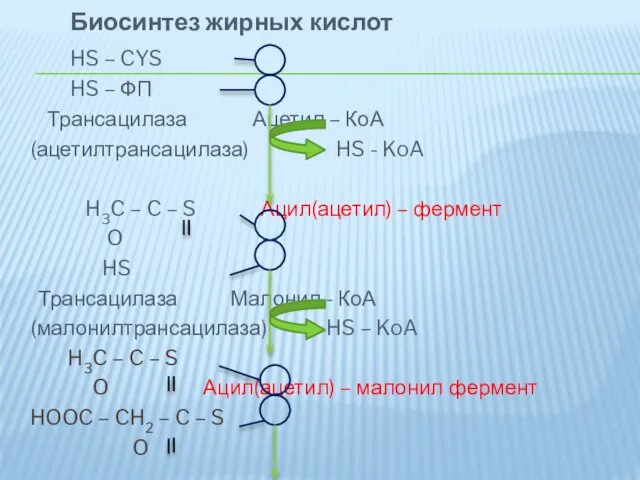
Биосинтез жирных кислот
HS – CYS
HS – ФП
Трансацилаза Ацетил –
Биосинтез жирных кислот
HS – CYS
HS – ФП
Трансацилаза Ацетил –
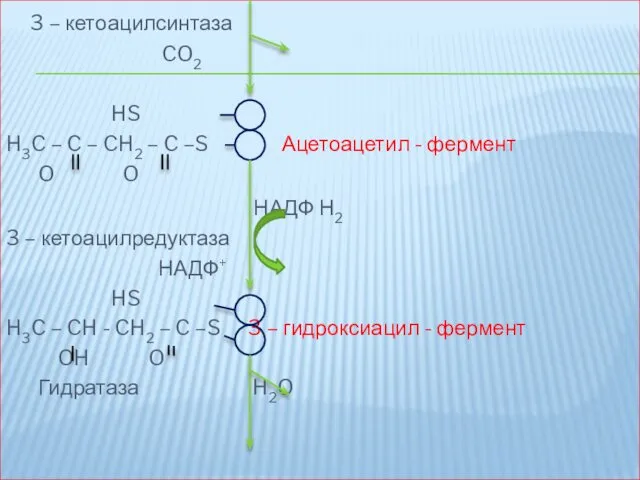
3 – кетоацилсинтаза
CO2
HS
H3C – C – CH2 – C
3 – кетоацилсинтаза
CO2
HS
H3C – C – CH2 – C
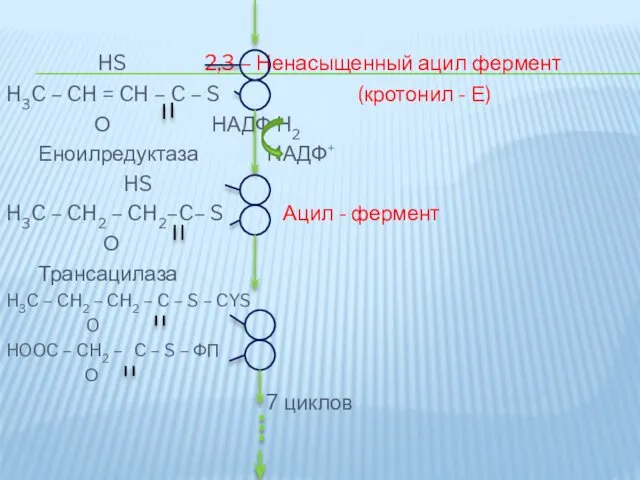
HS 2,3 – Ненасыщенный ацил фермент
H3C – CH = CH –
H3C – CH = CH –

Пальмитил – Е
Тиоэстераза
Н2О
Е
Пальмитиновая кислота
Тиоэстераза
Н2О
Е
Пальмитиновая кислота

Цикл синтеза повторяется.
Для синтеза пальмитиновой кислоты требуется 7 таких циклов, соответственно
Цикл синтеза повторяется.
Для синтеза пальмитиновой кислоты требуется 7 таких циклов, соответственно

Суммарное уравнение синтеза пальмитиновой кислоты
H3C – C(О) ~ SKOA + 7HOOC
H3C – C(О) ~ SKOA + 7HOOC

Различия процессов окисления и биосинтеза
Различия процессов окисления и биосинтеза

СИСТЕМА УДЛИНЕНИЯ (ЭЛОНГАЦИИ) ЖИРНЫХ КИСЛОТ
Данный процесс в основном происходит в микросомах.
СИСТЕМА УДЛИНЕНИЯ (ЭЛОНГАЦИИ) ЖИРНЫХ КИСЛОТ
Данный процесс в основном происходит в микросомах.

При голодании элонгация жирных кислот затормаживается. При образовании миелиновых оболочек нервных
При голодании элонгация жирных кислот затормаживается. При образовании миелиновых оболочек нервных

СИНТЕЗ НЕНАСЫЩЕННЫХ ЖИРНЫХ КИСЛОТ
Катализируются микросомальными системами путем введения двойной связи в
СИНТЕЗ НЕНАСЫЩЕННЫХ ЖИРНЫХ КИСЛОТ
Катализируются микросомальными системами путем введения двойной связи в

Стеароил – КоА + НАДН2 + О2
(НАДФН2)
Олеоил – КоА +
(НАДФН2)
Олеоил – КоА +

Таким образом идет синтез из пальмитиновой - пальмитиолеиновой кислоты, а из
Таким образом идет синтез из пальмитиновой - пальмитиолеиновой кислоты, а из
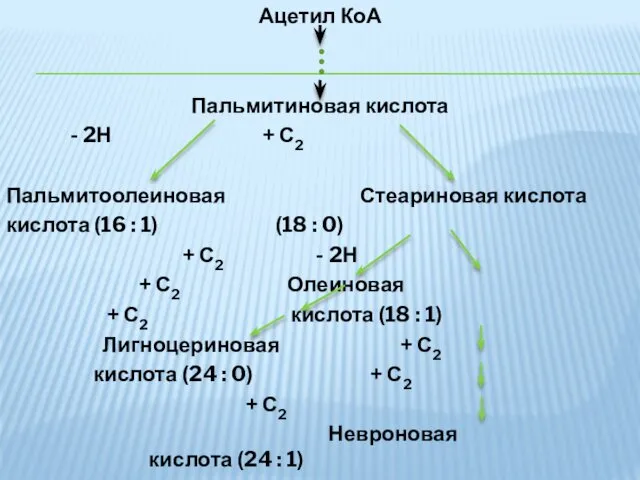
Ацетил КоА
Пальмитиновая кислота
- 2Н + С2
Пальмитоолеиновая Стеариновая кислота
кислота (16 : 1) (18
Ацетил КоА
Пальмитиновая кислота
- 2Н + С2
Пальмитоолеиновая Стеариновая кислота
кислота (16 : 1) (18
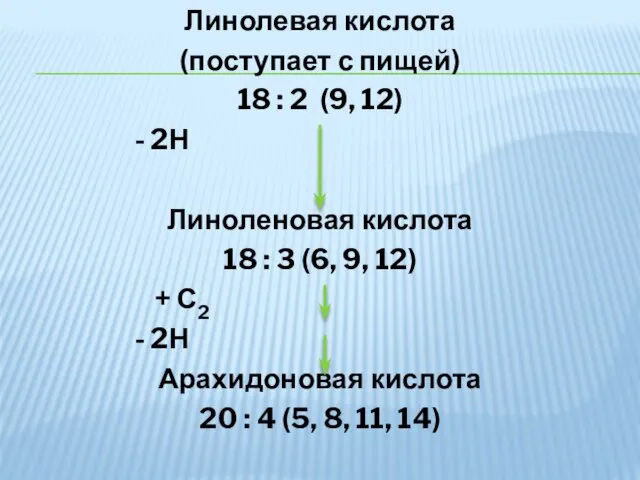
Линолевая кислота
(поступает с пищей)
18 : 2 (9, 12)
- 2Н
Линоленовая кислота
18 :
Линолевая кислота
(поступает с пищей)
18 : 2 (9, 12)
- 2Н
Линоленовая кислота
18 :

БИОСИНТЕЗ КЕТОНОВЫХ ТЕЛ (АЦЕТОНОВЫХ)
К ацетоновым телами относятся ацетоуксусная кислота, β- оксимасленая
БИОСИНТЕЗ КЕТОНОВЫХ ТЕЛ (АЦЕТОНОВЫХ)
К ацетоновым телами относятся ацетоуксусная кислота, β- оксимасленая
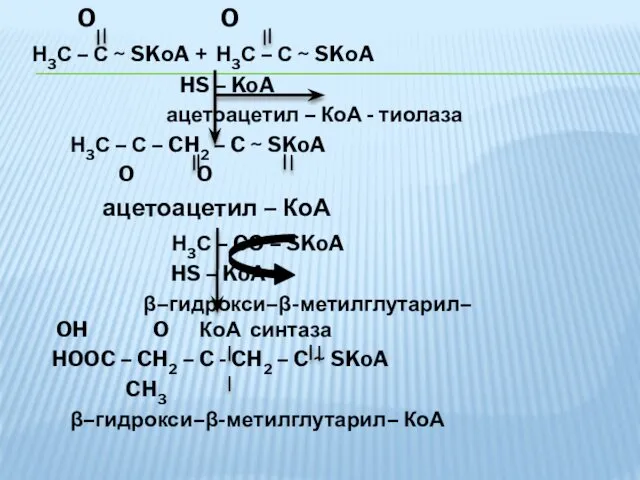
O O
Н3С – С ~ SKоA + Н3С –
O O
Н3С – С ~ SKоA + Н3С –
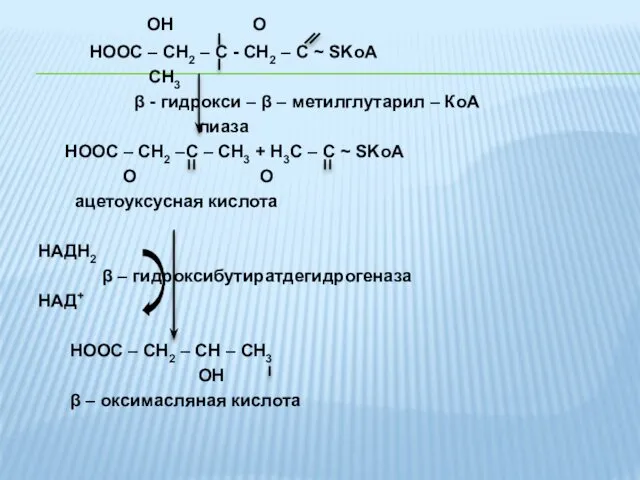
OH O
HOOC – CH2 – C - CH2 –
OH O
HOOC – CH2 – C - CH2 –

В организме кетоновые тела выполняют важную функцию по поддержанию энергетического гомеостаза.
В организме кетоновые тела выполняют важную функцию по поддержанию энергетического гомеостаза.
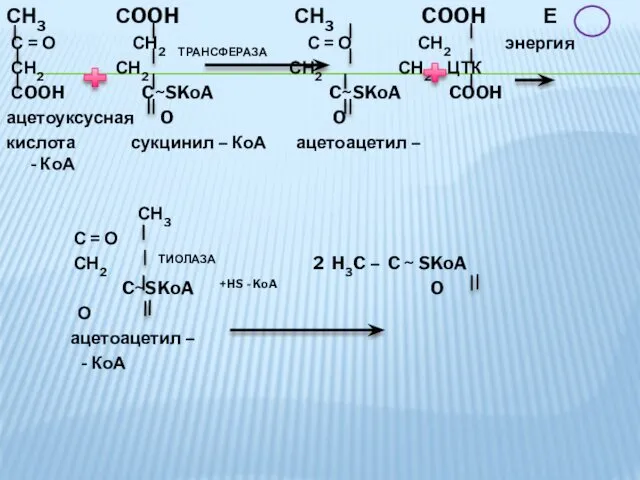
СН3 СOOH СН3 COOH Е
С = О СН2 ТРАНСФЕРАЗА
СН3 СOOH СН3 COOH Е
С = О СН2 ТРАНСФЕРАЗА

В тех случаях когда скорость образовании кетоновых тел превышает их потребление
В тех случаях когда скорость образовании кетоновых тел превышает их потребление

Патологический кетоз возникает при тяжелой форме сахарного диабета. Выделение кетоновых тел
Патологический кетоз возникает при тяжелой форме сахарного диабета. Выделение кетоновых тел
 Инерция. Билет 2
Инерция. Билет 2 Корпоративные ценности
Корпоративные ценности Смешарики. Война 12 эпизод
Смешарики. Война 12 эпизод Признаки текста. Анализ текста
Признаки текста. Анализ текста Методы краткосрочного прогнозирования экономических явлений. Сглаживание и экстраполяция
Методы краткосрочного прогнозирования экономических явлений. Сглаживание и экстраполяция Экспериментальная психология. Влияние экспериментатора и испытуемых на результаты эксперимента. (Лекция 5)
Экспериментальная психология. Влияние экспериментатора и испытуемых на результаты эксперимента. (Лекция 5) Марокко. Основные сведения о Марокко
Марокко. Основные сведения о Марокко Технико-эксплуатационные показатели использования пассажирского автотранспорта
Технико-эксплуатационные показатели использования пассажирского автотранспорта Храм Преображения Господня на Преображенской площади
Храм Преображения Господня на Преображенской площади Проектирование разработки сеноманских отложений на Заполярном месторождении
Проектирование разработки сеноманских отложений на Заполярном месторождении Организация производства на предприятии TOYOTA
Организация производства на предприятии TOYOTA Звіт за результатами роботи відділу здоров’я та спорту та перспективи на 2016 рік
Звіт за результатами роботи відділу здоров’я та спорту та перспективи на 2016 рік Журнал Актуальные проблемы российского права
Журнал Актуальные проблемы российского права Память о войне нам книга оживляет
Память о войне нам книга оживляет Урок-инструктаж Как подготовить реферат и презентацию по обществознанию
Урок-инструктаж Как подготовить реферат и презентацию по обществознанию Общая характеристика основных видов транспорта нефти, нефтепродуктов и газа. Лекция №1
Общая характеристика основных видов транспорта нефти, нефтепродуктов и газа. Лекция №1 Осветление воды методами фильтрования
Осветление воды методами фильтрования Чтение топографических карт
Чтение топографических карт Ускорение. Торможение
Ускорение. Торможение Легисакционный процесс
Легисакционный процесс Гимнастическая терминология
Гимнастическая терминология Предметно-развивающая среда
Предметно-развивающая среда Sergis General Information
Sergis General Information Многостраничный дизайн
Многостраничный дизайн Системы вентиляции и кондиционирования воздуха
Системы вентиляции и кондиционирования воздуха Культура первобытного общества
Культура первобытного общества Детский церебральный паралич
Детский церебральный паралич Мальчики или девочки: кто умнее?
Мальчики или девочки: кто умнее?