- Дивергенция, конвергенция и параллелизм

Содержание
- 2. Понятие дивергенции Дивергенция – явление расхождения признаков у близких форм в процессе эволюции. Термин «дивергенция» (в
- 3. Адаптивная радиация Генри Осборн расширил понятие дивергенции, введя термин «адаптивная радиация». По Осборну, это процесс, при
- 4. Как работает дивергенция у прокариот Вообще сложно говорить о дивергенции у прокариот, так как они практикуют
- 5. Как работает дивергенция у эукариот Основным фактором расхождения признаков, а в дальнейшем – видообразования у эукариот,
- 6. Как работает дивергенция у эукариот По-видимому, предки современного человека и шимпанзе разделились в какой-то степени географически.
- 7. Как работает дивергенция у эукариот Экологическая изоляция происходит в пределах одной территории, когда часть популяции по
- 8. Как работает дивергенция у эукариот Репродуктивная изоляция – самый главный фактор, способствующий видообразованию. Подразделяется на презиготическую
- 9. Конвергенция и параллелизм Конвергенция – независимое приобретение неродственными организмами сходных признаков; поверхностна, затрагивает немногие черты организации
- 10. Глубокая гомология Глубокая гомология – явление, при котором негомологичные органы развиваются сходным путём, на основе работы
- 11. Список литературы Дарвин, Ч. Происхождение видов путём естественного отбора. В 2 кн. Кн 1: Происхождение видов:
- 13. Скачать презентацию

Понятие дивергенции
Дивергенция – явление расхождения признаков у близких форм в
Понятие дивергенции
Дивергенция – явление расхождения признаков у близких форм в

Адаптивная радиация
Генри Осборн расширил понятие дивергенции, введя термин «адаптивная радиация».
Адаптивная радиация
Генри Осборн расширил понятие дивергенции, введя термин «адаптивная радиация».
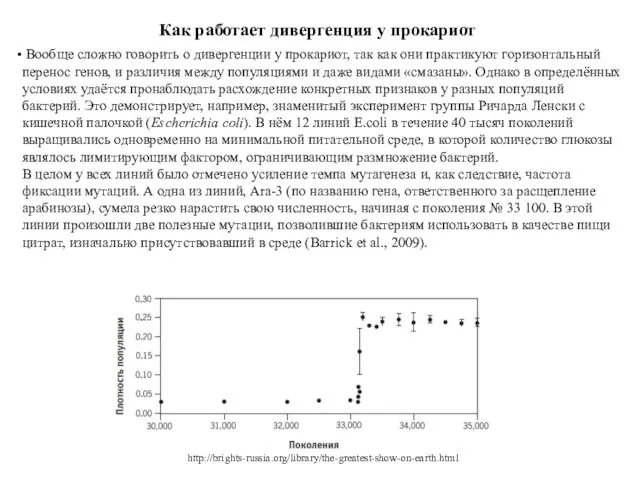
Как работает дивергенция у прокариот
Вообще сложно говорить о дивергенции у
Как работает дивергенция у прокариот
Вообще сложно говорить о дивергенции у

Как работает дивергенция у эукариот
Основным фактором расхождения признаков, а в
Как работает дивергенция у эукариот
Основным фактором расхождения признаков, а в

Как работает дивергенция у эукариот
По-видимому, предки современного человека и шимпанзе разделились
Как работает дивергенция у эукариот
По-видимому, предки современного человека и шимпанзе разделились

Как работает дивергенция у эукариот
Экологическая изоляция происходит в пределах одной
Как работает дивергенция у эукариот
Экологическая изоляция происходит в пределах одной

Как работает дивергенция у эукариот
Репродуктивная изоляция – самый главный фактор,
Как работает дивергенция у эукариот
Репродуктивная изоляция – самый главный фактор,
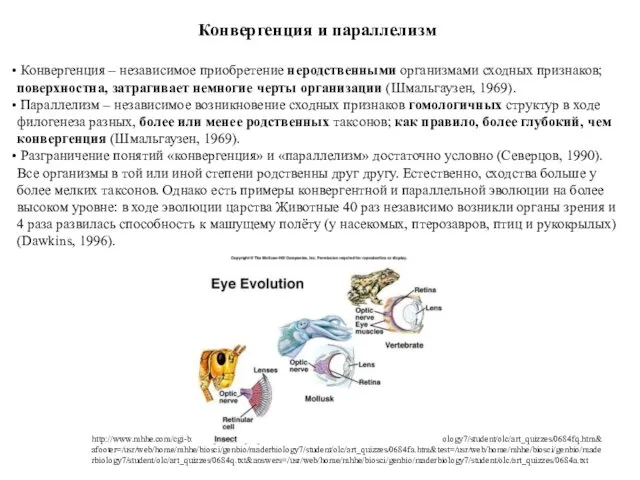
Конвергенция и параллелизм
Конвергенция – независимое приобретение неродственными организмами сходных признаков;
Конвергенция и параллелизм
Конвергенция – независимое приобретение неродственными организмами сходных признаков;
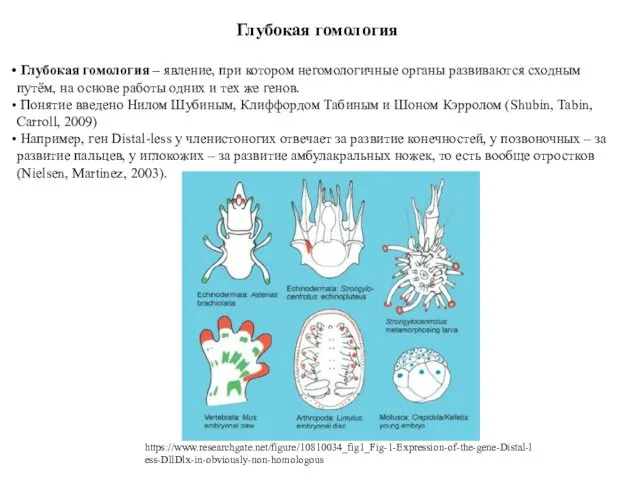
Глубокая гомология
Глубокая гомология – явление, при котором негомологичные органы развиваются
Глубокая гомология
Глубокая гомология – явление, при котором негомологичные органы развиваются

Список литературы
Дарвин, Ч. Происхождение видов путём естественного отбора. В 2 кн.
Список литературы
Дарвин, Ч. Происхождение видов путём естественного отбора. В 2 кн.
 Оборудование для испытания пластов. Семинар 15
Оборудование для испытания пластов. Семинар 15 Презентация А. Барто
Презентация А. Барто Обобщение и систематизация знаний по теме Спирты и фенолы
Обобщение и систематизация знаний по теме Спирты и фенолы Обзор Дорожной карты по формированию и развитию благоприятных условий для духовного просвещения детей
Обзор Дорожной карты по формированию и развитию благоприятных условий для духовного просвещения детей Тексты и изображения в памяти компьютера
Тексты и изображения в памяти компьютера 29.01 Двугранный угол1_
29.01 Двугранный угол1_ Здоровый образ жизни
Здоровый образ жизни Защита диплома
Защита диплома Что мы знаем о правилах дорожного движения. Игра для детей 3-4 лет
Что мы знаем о правилах дорожного движения. Игра для детей 3-4 лет Теорія граничної корисності і поведінка споживача
Теорія граничної корисності і поведінка споживача Крупнощитовая разборно-переставная опалубка
Крупнощитовая разборно-переставная опалубка 20231024_pavlyuchenko_v.yu_
20231024_pavlyuchenko_v.yu_ Конспект урока технологии в 4 классе по теме: Реалистическая лепка
Конспект урока технологии в 4 классе по теме: Реалистическая лепка Скоро-скоро Новый год
Скоро-скоро Новый год Fauna of Kazakhstan
Fauna of Kazakhstan Развитие хозяйства
Развитие хозяйства Показатели характеризующие производственный микроклимат, их гигиеническая оценка
Показатели характеризующие производственный микроклимат, их гигиеническая оценка Викторина по теме озеро Байкал.
Викторина по теме озеро Байкал. Факторинговые компании
Факторинговые компании Агропромышленный комплекс. Растениеводство
Агропромышленный комплекс. Растениеводство Технология выполнения успешного исследования в области английского языка. Структура защиты
Технология выполнения успешного исследования в области английского языка. Структура защиты ХОЗЯЙСТВЕННАЯ ДЕЯТЕЛЬНОСТЬ ДАЛЬНЕГО ВОСТОКА
ХОЗЯЙСТВЕННАЯ ДЕЯТЕЛЬНОСТЬ ДАЛЬНЕГО ВОСТОКА прзтц
прзтц Оборудование для добычи нефти и газа. Оборудование для эксплуатации скважин фонтанным и газлифтным способами
Оборудование для добычи нефти и газа. Оборудование для эксплуатации скважин фонтанным и газлифтным способами Число и цифра 5. Состав числа 5. Сложение и вычитание в пределах 5. Пятиугольник
Число и цифра 5. Состав числа 5. Сложение и вычитание в пределах 5. Пятиугольник Презентация Концепция Курта Левина.
Презентация Концепция Курта Левина. Автоматизация водопроводных и канализационных насосных станции
Автоматизация водопроводных и канализационных насосных станции Цифровые многоканальные телекоммуникационные системы
Цифровые многоканальные телекоммуникационные системы