- Физиологическая роль мышц

Содержание
- 2. Физиологическая роль мышц Поддержание позы и равновесия тела Произвольные движения Воспроизведение речи Работа сердца Регуляция тонуса
- 3. У детей В постнатальном периоде до реализации позы, функция мышечной системы в значительной степени связана с
- 4. Двигательная единица Комплекс, включающий один мотонейрон и иннервируемые им мышечные волокна, называется ДВИГАТЕЛЬНОЙ ИЛИ НЕЙРОМОТОРНОЙ ЕДИНИЦЕЙ
- 5. Два варианта классификации скелетных мышц Анатомическая По плотности иннервации (количество двигательных единиц в мышце) Функциональная По
- 6. Классификация по количеству двигательных единиц в мышце Много двигательных единиц на одну мышцу Плотность иннервации высокая
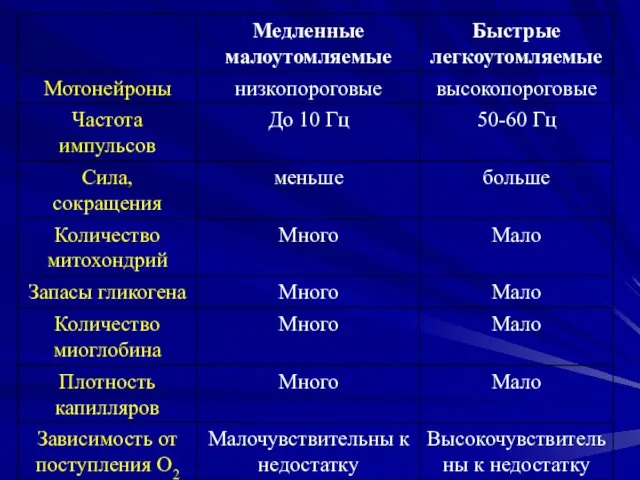
- 7. Классификация по работоспособности Медленные малоутомляемые Быстрые легкоутомляемые
- 9. Свойства мышцы Возбудимость Проводимость Сократимость Эластичность – способность сокращаться при растягивании. Тонус
- 10. Мышечное волокно
- 11. Саркомер
- 12. Возбуждение мышечных волокон
- 13. Одно мышечное волокно получает нервный импульс от одного синапса
- 14. Нервно-мышечный синапс (концевая пластинка)
- 15. Медиатор – ацетилхолин На постсинаптической мембране Н-холинорецепторы
- 16. Потенциал концевой пластинки
- 17. Особенности нервно-мышечного синапса Большая поверхность пресинаптической мембраны Синаптическая щель содержит много ГАГ, митохондрий Большая складчатость постсинаптической
- 18. У детей постсинаптическая мембрана не сформирована, мышечное волокно чувствительно к ацетилхолину на всем его протяжении, а
- 19. ПД распространяется по мембране мышечного волокна
- 20. Результат возбуждения - увеличение концентрации внутриклеточного кальция
- 21. Потенциал концевой пластинки (ПКП) и потенциал действия (ПД) мышечного волокна ПКП ПД Синапс ПД Са++ СПР
- 22. У детей Возбудимость мышц плода и детей низка, т.к. мембранный потенциал покоя значительно менее отрицателен, чем
- 23. Последовательность событий при возбуждении ПД пресинаптического окончания приводит к выделению медиатора. Возникновение ПКП на постсинаптической мембране.
- 24. Сокращение Сократительные и регуляторные белки саркомера
- 25. Саркомер – функциональная единица сократительного аппарата мышечной клетки. Длина саркомера 2,5 мкм, поперечник – 1мкм .
- 28. Толстые миофиламенты
- 29. Свойства миозина Толстые миофиламенты – образованы молекулами миозина, Тяжелые цепи миозина - головка миозина и шейка.
- 30. Тонкие миофиламенты
- 31. Свойства актина Тонкие миофиламенты построены из глобулярных молекул белка актина. Актиновые филаменты представляют собой закрученную двойную
- 32. Регуляторные белки В продольных бороздках актиновой спирали располагаются нитевидные молекулы белка тропомиозина. Тропомиозин закрывает активные центры
- 34. Контакт актина с миозином возможен, если тропомиозин сдвинется и откроет активные центры на тонких – актиновых
- 35. Ключевой момент – переход отвозбуждения к сокращению Связывание ионов кальция молекулами тропонина Изменение конформации тропомиозина Сдвиг
- 37. «гребок» В момент контакта головка миозина совершает «гребковое» движение и передвигает тонкую миофиламенту вдоль толстой по
- 38. Укорочение саркомера
- 45. Последовательность событий при сокращении. молекулы тропомиозина опускаются в желобки между цепочками мономеров актина, открывая участки прикрепления
- 46. Расслабление Работа Са++ АТФ-азы Снижение концентрации внутриклеточного Са++ Восстановление конформации тропомиозина Активные центры тонких миофиламентов закрыты.
- 47. Временная характеристика процесса сокращения
- 48. Сопоставление во времени сокращения мышцы и возбудимости Возбудимость мышцы во время сокращения нормальна, следовательно она может
- 49. Запись мышечных сокращений
- 50. Суммация сокращений
- 51. Режимы мышечного сокращения Одиночное сокращение Тетанус – слитное сокращение без расслабления Зубчатый – импульс в фазу
- 52. Режимы сокращения Одиночное Зубчатый тетанус Гладкий тетанус Оптимум
- 53. СКЕЛЕТНЫЕ МЫШЦЫ НОВОРОЖДЕННЫХ НЕ СПОСОБНЫ РАЗВИВАТЬ ТЕТАНУС - низкая функциональная лабильность ( высокая продолжительность ПД).
- 54. Виды мышечного сокращения Виды Изометрическое Изотоническое Ауксотоническое
- 55. Регуляция силы сокращения мышцы Сила сокращения зависит от числа включенных мышечных волокон частоты импульсов возбуждения –
- 57. Скачать презентацию

Физиологическая роль мышц
Поддержание позы и равновесия тела
Произвольные движения
Воспроизведение речи
Работа
Физиологическая роль мышц
Поддержание позы и равновесия тела
Произвольные движения
Воспроизведение речи
Работа

У детей
В постнатальном периоде до реализации позы, функция мышечной системы в
У детей
В постнатальном периоде до реализации позы, функция мышечной системы в
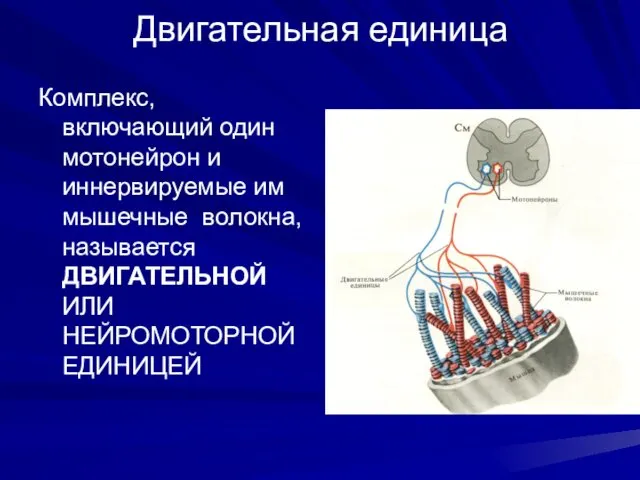
Двигательная единица
Комплекс, включающий один мотонейрон и иннервируемые им мышечные волокна, называется
Двигательная единица
Комплекс, включающий один мотонейрон и иннервируемые им мышечные волокна, называется

Два варианта классификации
скелетных мышц
Анатомическая По плотности иннервации (количество двигательных единиц в
Два варианта классификации
скелетных мышц
Анатомическая По плотности иннервации (количество двигательных единиц в

Классификация по количеству двигательных единиц в мышце
Много двигательных единиц на одну
Классификация по количеству двигательных единиц в мышце
Много двигательных единиц на одну

Классификация
по работоспособности
Медленные малоутомляемые
Быстрые легкоутомляемые
Классификация
по работоспособности
Медленные малоутомляемые
Быстрые легкоутомляемые

Свойства мышцы
Возбудимость
Проводимость
Сократимость
Эластичность – способность сокращаться при растягивании.
Тонус
Свойства мышцы
Возбудимость
Проводимость
Сократимость
Эластичность – способность сокращаться при растягивании.
Тонус
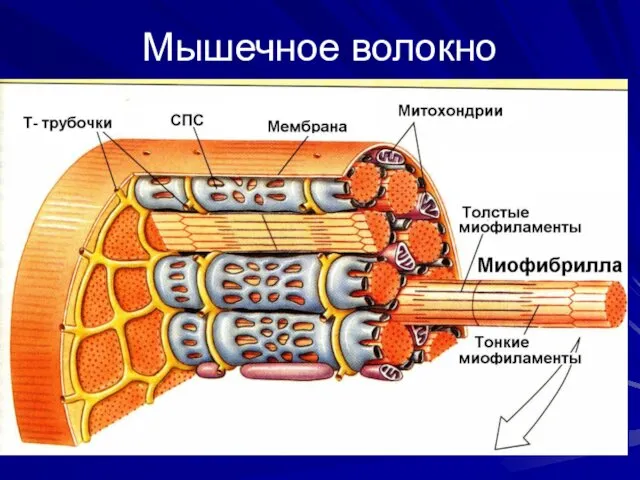
Мышечное волокно
Мышечное волокно
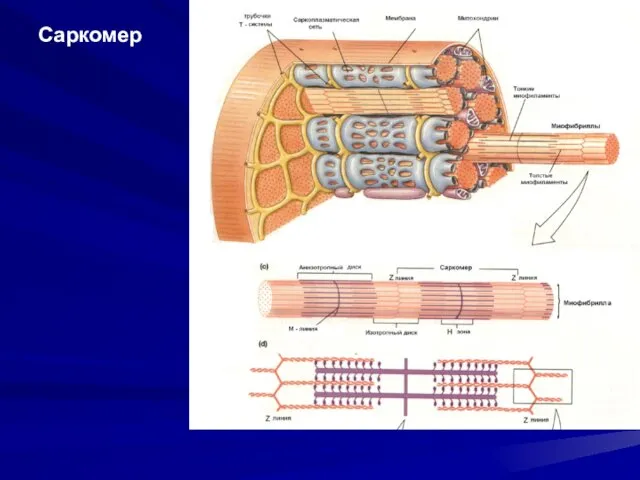
Саркомер
Саркомер

Возбуждение мышечных волокон
Возбуждение мышечных волокон
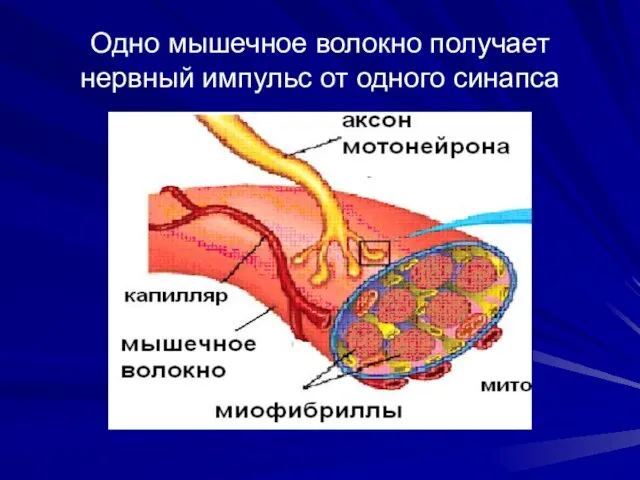
Одно мышечное волокно получает нервный импульс от одного синапса
Одно мышечное волокно получает нервный импульс от одного синапса
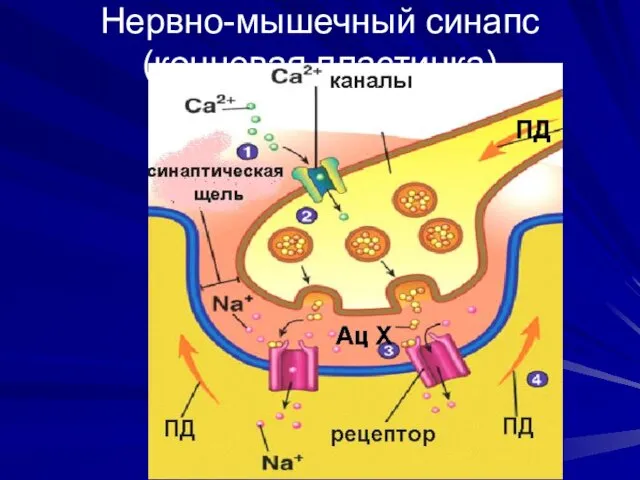
Нервно-мышечный синапс (концевая пластинка)
Нервно-мышечный синапс (концевая пластинка)
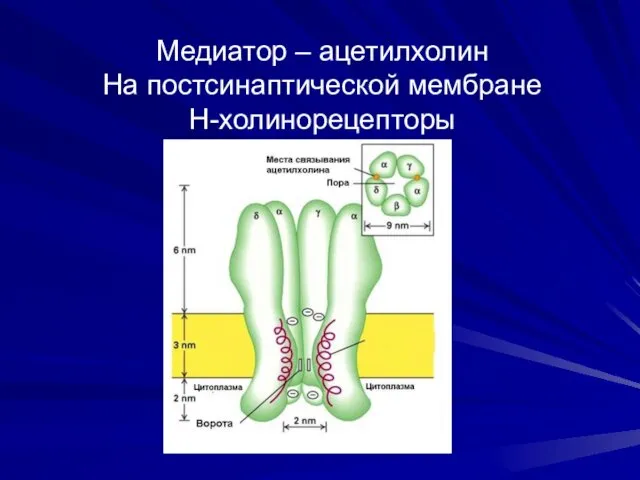
Медиатор – ацетилхолин
На постсинаптической мембране
Н-холинорецепторы
Медиатор – ацетилхолин
На постсинаптической мембране
Н-холинорецепторы
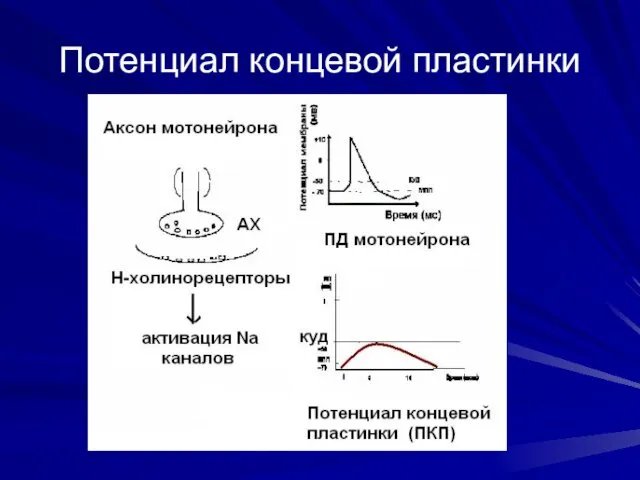
Потенциал концевой пластинки
Потенциал концевой пластинки

Особенности
нервно-мышечного синапса
Большая поверхность пресинаптической мембраны
Синаптическая щель содержит много ГАГ, митохондрий
Большая
Особенности
нервно-мышечного синапса
Большая поверхность пресинаптической мембраны
Синаптическая щель содержит много ГАГ, митохондрий
Большая

У детей
постсинаптическая мембрана не сформирована, мышечное волокно чувствительно к ацетилхолину на
У детей
постсинаптическая мембрана не сформирована, мышечное волокно чувствительно к ацетилхолину на
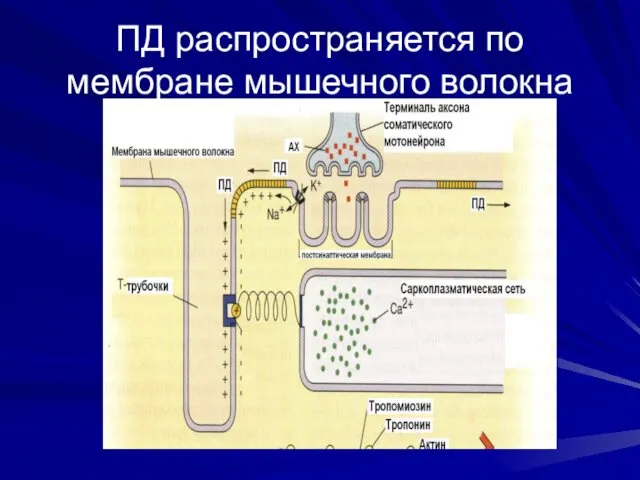
ПД распространяется по мембране мышечного волокна
ПД распространяется по мембране мышечного волокна
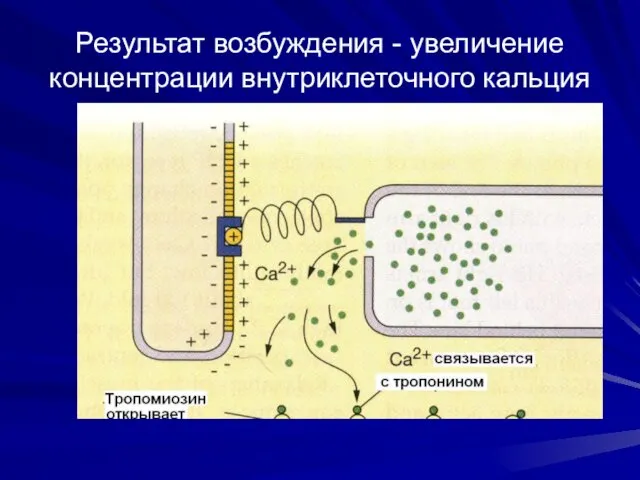
Результат возбуждения - увеличение концентрации внутриклеточного кальция
Результат возбуждения - увеличение концентрации внутриклеточного кальция
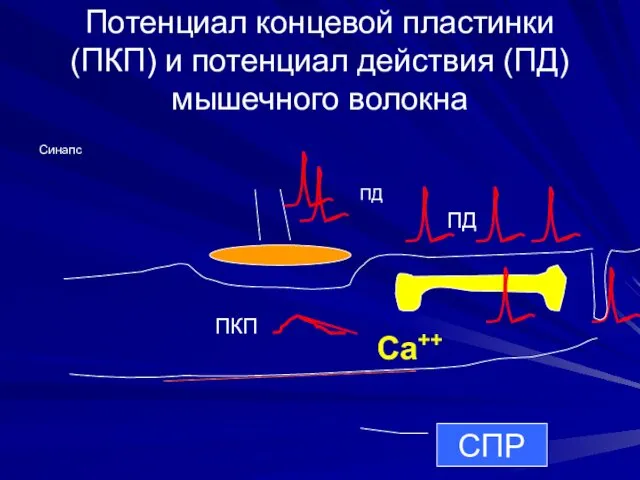
Потенциал концевой пластинки (ПКП) и потенциал действия (ПД) мышечного волокна
ПКП
ПД
Синапс
ПД
Са++
СПР
Потенциал концевой пластинки (ПКП) и потенциал действия (ПД) мышечного волокна
ПКП
ПД
Синапс
ПД
Са++
СПР

У детей
Возбудимость мышц плода и детей низка, т.к. мембранный потенциал покоя
У детей
Возбудимость мышц плода и детей низка, т.к. мембранный потенциал покоя

Последовательность событий при возбуждении
ПД пресинаптического окончания приводит к выделению медиатора.
Возникновение
Последовательность событий при возбуждении
ПД пресинаптического окончания приводит к выделению медиатора.
Возникновение

Сокращение
Сократительные и регуляторные белки саркомера
Сокращение
Сократительные и регуляторные белки саркомера
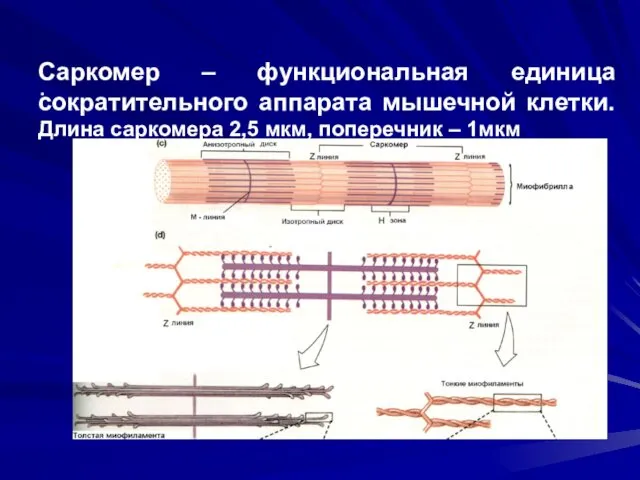
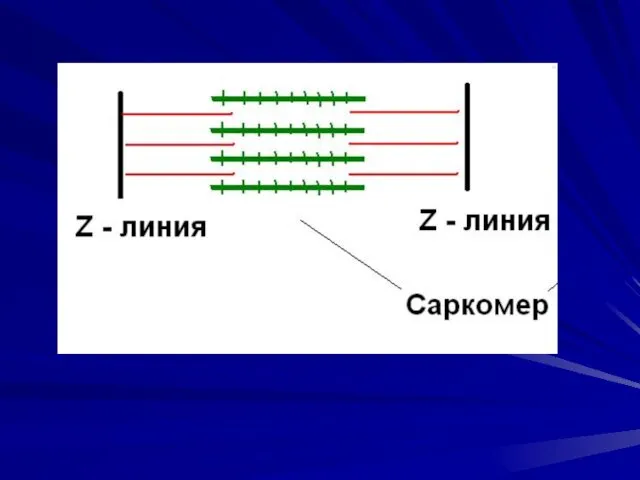
Саркомер – функциональная единица сократительного аппарата мышечной клетки.
Длина саркомера 2,5 мкм,
Саркомер – функциональная единица сократительного аппарата мышечной клетки. Длина саркомера 2,5 мкм,
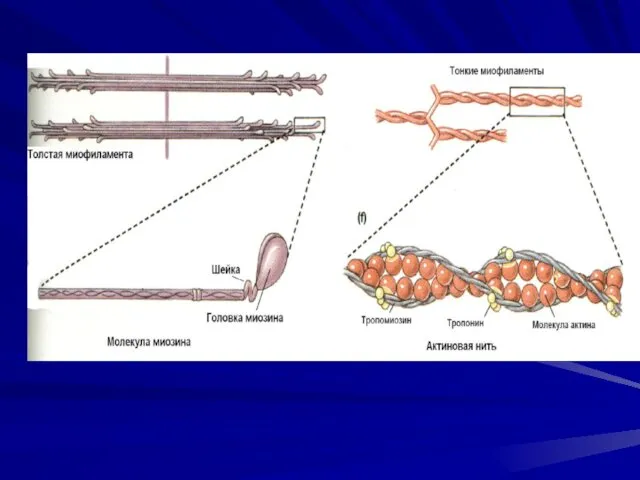
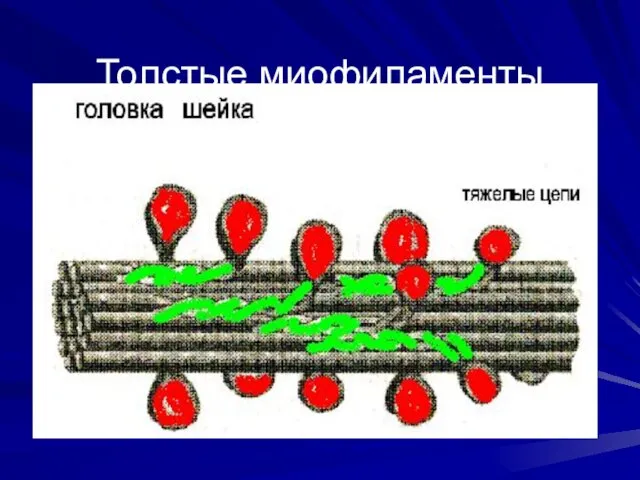
Толстые миофиламенты
Толстые миофиламенты

Свойства миозина
Толстые миофиламенты – образованы молекулами миозина, Тяжелые цепи миозина -
Свойства миозина
Толстые миофиламенты – образованы молекулами миозина, Тяжелые цепи миозина -
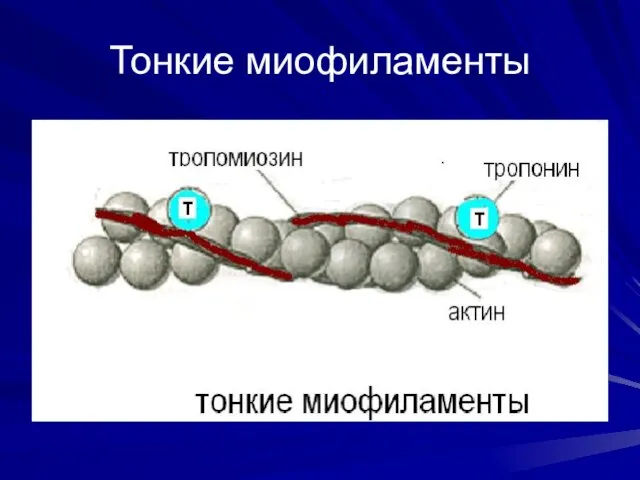
Тонкие миофиламенты
Тонкие миофиламенты

Свойства актина
Тонкие миофиламенты построены из глобулярных молекул белка актина. Актиновые филаменты
Свойства актина
Тонкие миофиламенты построены из глобулярных молекул белка актина. Актиновые филаменты

Регуляторные белки
В продольных бороздках актиновой спирали располагаются нитевидные молекулы белка тропомиозина.
Регуляторные белки
В продольных бороздках актиновой спирали располагаются нитевидные молекулы белка тропомиозина.
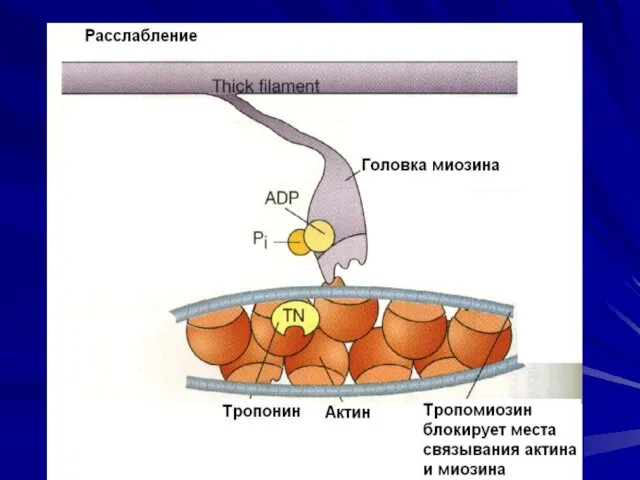

Контакт актина с миозином возможен, если тропомиозин сдвинется и откроет активные
Контакт актина с миозином возможен, если тропомиозин сдвинется и откроет активные

Ключевой момент – переход отвозбуждения к сокращению
Связывание ионов кальция молекулами тропонина
Изменение
Ключевой момент – переход отвозбуждения к сокращению
Связывание ионов кальция молекулами тропонина
Изменение
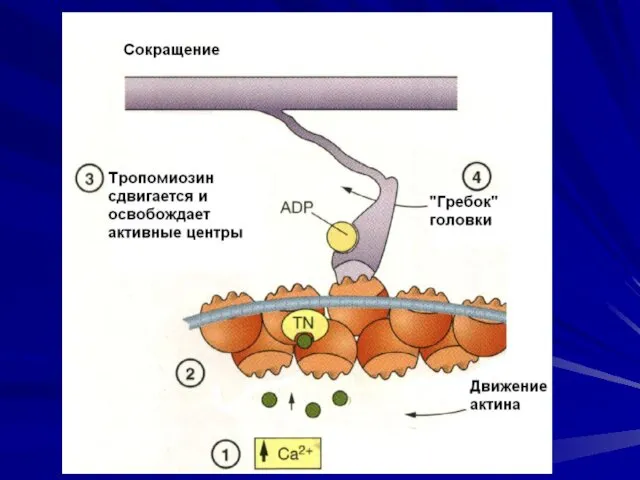

«гребок»
В момент контакта головка миозина совершает «гребковое» движение и передвигает тонкую
«гребок»
В момент контакта головка миозина совершает «гребковое» движение и передвигает тонкую
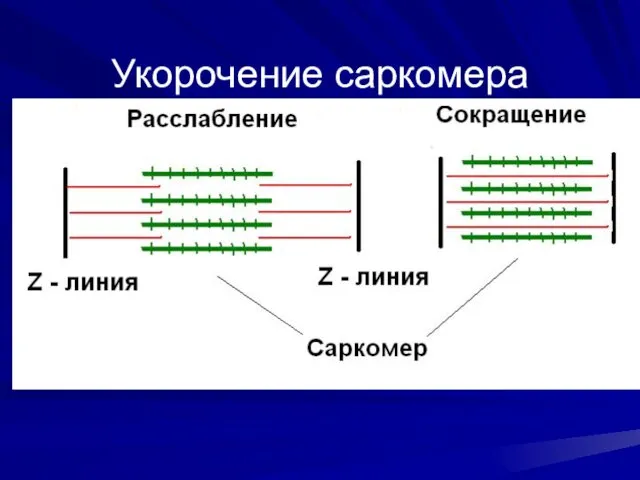
Укорочение саркомера
Укорочение саркомера
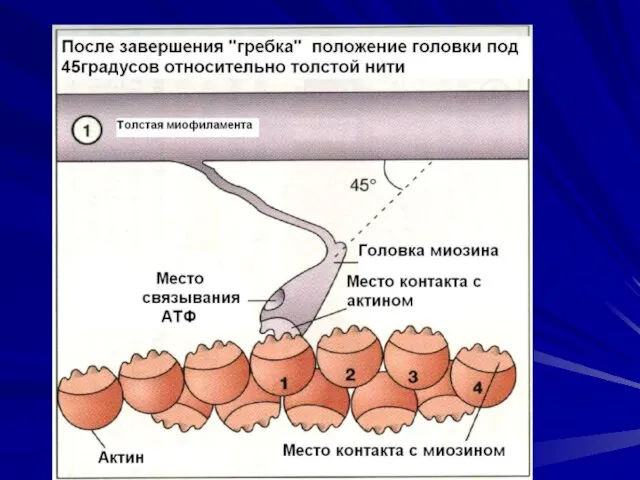
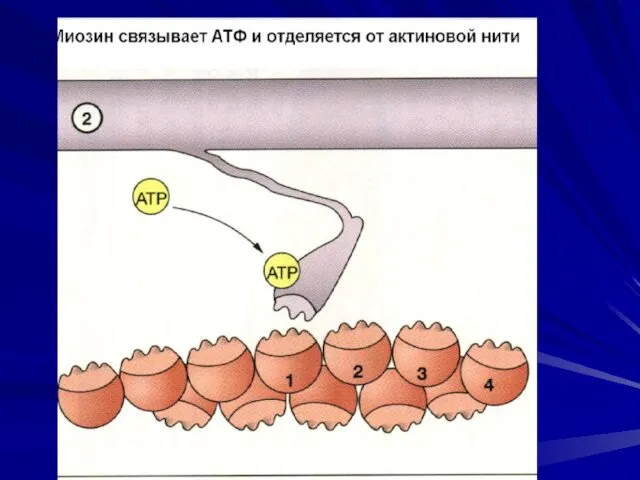
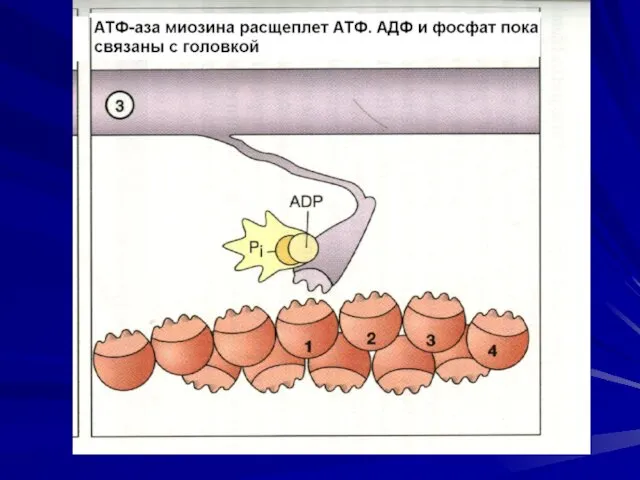
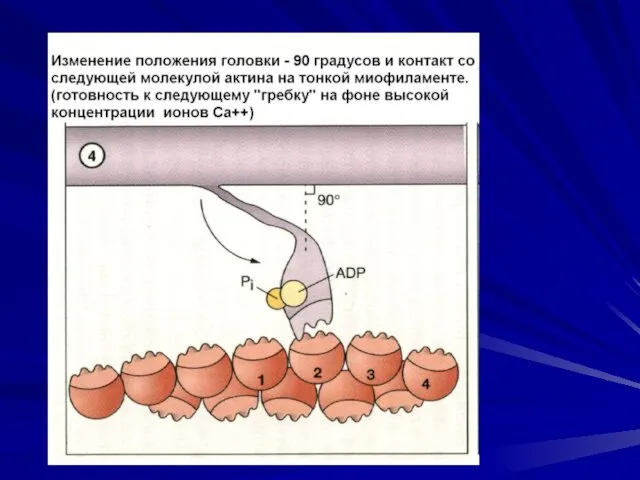
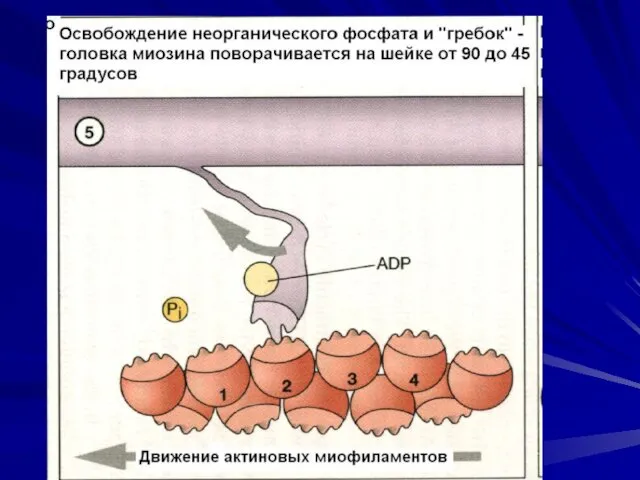
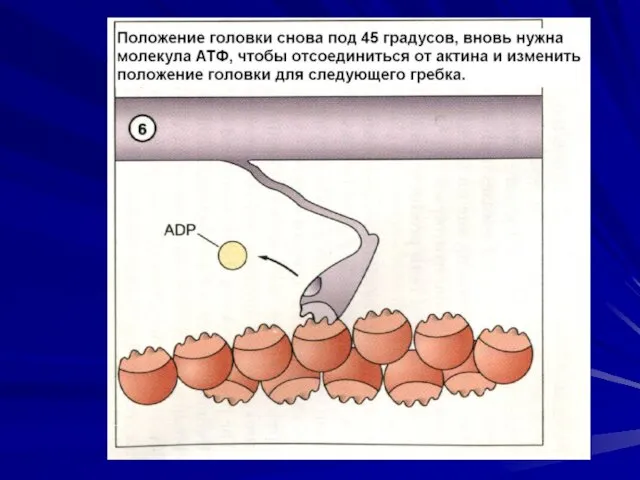

Последовательность событий при сокращении.
молекулы тропомиозина опускаются в желобки между цепочками
Последовательность событий при сокращении.
молекулы тропомиозина опускаются в желобки между цепочками

Расслабление
Работа Са++ АТФ-азы
Снижение концентрации внутриклеточного Са++
Восстановление конформации тропомиозина
Активные центры тонких миофиламентов
Расслабление
Работа Са++ АТФ-азы
Снижение концентрации внутриклеточного Са++
Восстановление конформации тропомиозина
Активные центры тонких миофиламентов
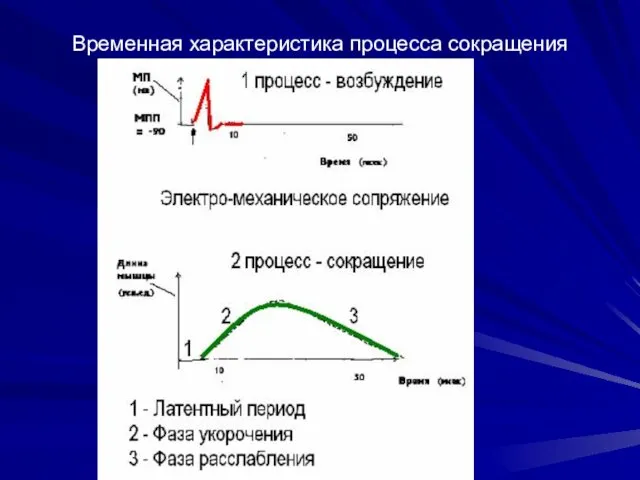
Временная характеристика процесса сокращения
Временная характеристика процесса сокращения

Сопоставление во времени сокращения мышцы и возбудимости
Возбудимость мышцы во время сокращения
Сопоставление во времени сокращения мышцы и возбудимости
Возбудимость мышцы во время сокращения
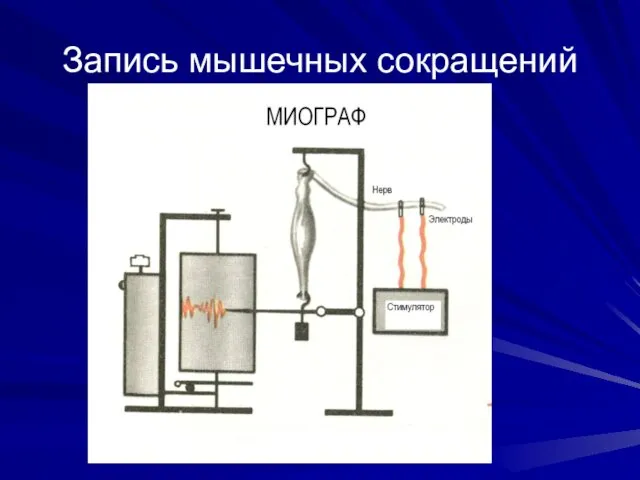
Запись мышечных сокращений
Запись мышечных сокращений
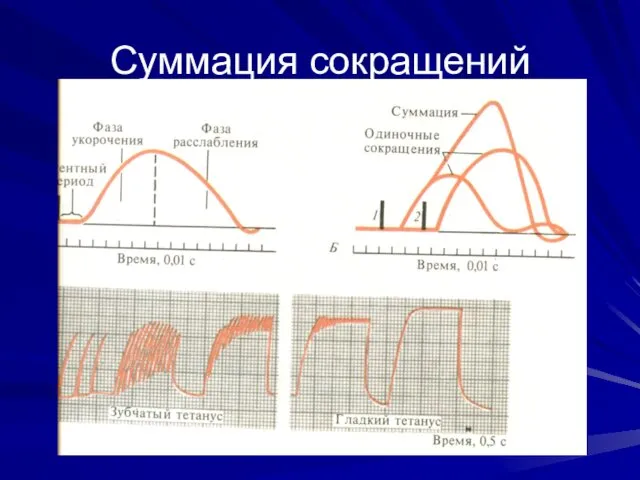
Суммация сокращений
Суммация сокращений

Режимы
мышечного сокращения
Одиночное сокращение
Тетанус – слитное сокращение без расслабления
Зубчатый – импульс в
Режимы
мышечного сокращения
Одиночное сокращение
Тетанус – слитное сокращение без расслабления
Зубчатый – импульс в
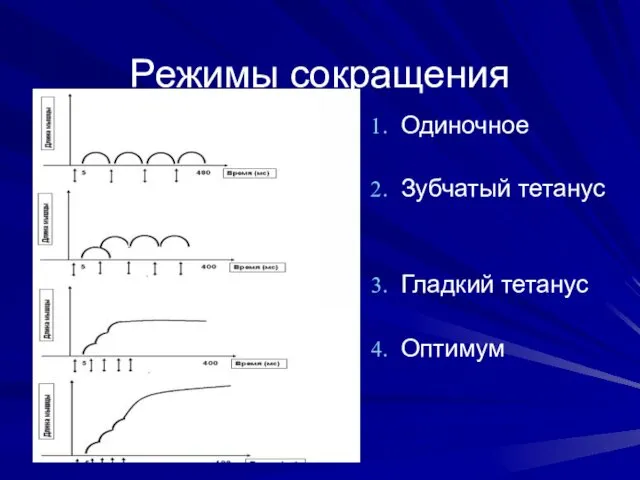
Режимы сокращения
Одиночное
Зубчатый тетанус
Гладкий тетанус
Оптимум
Режимы сокращения
Одиночное
Зубчатый тетанус
Гладкий тетанус
Оптимум

СКЕЛЕТНЫЕ МЫШЦЫ НОВОРОЖДЕННЫХ НЕ СПОСОБНЫ РАЗВИВАТЬ ТЕТАНУС - низкая функциональная лабильность
СКЕЛЕТНЫЕ МЫШЦЫ НОВОРОЖДЕННЫХ НЕ СПОСОБНЫ РАЗВИВАТЬ ТЕТАНУС - низкая функциональная лабильность

Виды мышечного сокращения
Виды
Изометрическое
Изотоническое
Ауксотоническое
Виды мышечного сокращения
Виды
Изометрическое
Изотоническое
Ауксотоническое

Регуляция
силы сокращения мышцы
Сила сокращения зависит от
числа включенных мышечных волокон
частоты
Регуляция
силы сокращения мышцы
Сила сокращения зависит от
числа включенных мышечных волокон
частоты
 Мои любимые произведения
Мои любимые произведения Функциональная анатомия жевательных мышц и дна полости рта
Функциональная анатомия жевательных мышц и дна полости рта Виртуальная экскурсия по городу Славгороду
Виртуальная экскурсия по городу Славгороду Внутренняя политика Николая I
Внутренняя политика Николая I Сергей Александрович Есенин
Сергей Александрович Есенин Технология конструкционных материалов
Технология конструкционных материалов Construction of Reservoir, Sedimentation Tank & Irrigation System at Berry Farms in Uzbekistan
Construction of Reservoir, Sedimentation Tank & Irrigation System at Berry Farms in Uzbekistan Проект Контакт
Проект Контакт Капля бережет море
Капля бережет море Инвестиции и инвестиционные решения
Инвестиции и инвестиционные решения МСР. Южный округ Москвы. Чертаново-Центральное.
МСР. Южный округ Москвы. Чертаново-Центральное. Christina Stürmer
Christina Stürmer ЭХОКГ в пульмонологии
ЭХОКГ в пульмонологии Проект Борьба за чистоту русского языка
Проект Борьба за чистоту русского языка Харківський регіональний центр оцінювання якості освіти
Харківський регіональний центр оцінювання якості освіти ПОРТФОЛИО УЧИТЕЛЯ
ПОРТФОЛИО УЧИТЕЛЯ Уровни параллелизма. Классификации архитектур ВС
Уровни параллелизма. Классификации архитектур ВС История письменности на Руси
История письменности на Руси Игра лазертаг
Игра лазертаг Rio-de-Janeiro República Federativa do Brasil
Rio-de-Janeiro República Federativa do Brasil Агентство креативных решений
Агентство креативных решений Пруды города Димитровграда
Пруды города Димитровграда Что у осени в корзинке. Викторина
Что у осени в корзинке. Викторина Разработка технологического процесса изготовления упаковочной продукции флексографским способом печати
Разработка технологического процесса изготовления упаковочной продукции флексографским способом печати Хозяйство Центрального района
Хозяйство Центрального района Книга Памяти п.ст. Тарасиха, д.Тарасиха Нижегородской области
Книга Памяти п.ст. Тарасиха, д.Тарасиха Нижегородской области Наукометрия и библиометрия
Наукометрия и библиометрия Вычислительные приборы
Вычислительные приборы