- Физиология переднего мозга. Физиология вегетативной нервной системы. Лекция 10

Содержание
- 2. По А. Р. Лурия, реальным анатомическим и функциональным образованием, включенным в реализацию двигательного акта, помимо собственно
- 3. Моделью работы рано созревающего и еще не контролируемого корой паллидарного комплекса являются беспорядочные, нецеленаправленные движения бодрствующего
- 4. Рис. 12. Структура эстрапирамидной системы
- 5. При поражении старого отдела экстрапирамидной системы возникает противоположная картина. У больных появляется скованность (ригидность), бедность и
- 6. Пирамидная система (кортико-спинальный путь) начинается от моторных (крупных пирамидных) клеток Беца, находящихся в основном в 5-м
- 7. Третья группа волокон пирамидной системы, после частичного перекреста на уровне среднего мозга, варолиева моста и продолговатого
- 8. 10.2. ЛИМБИЧЕСКАЯ СИСТЕМА: СТРУКТУРЫ И ФУНКЦИИ. Лимбическая система. Передний мозг, интегративная деятельность которого обеспечивает целенаправленное поведение,
- 9. В настоящее время хорошо известны связи между структурами мозга, организующие круги, имеющие свою функциональную спе¬цифику. К
- 10. Лимбическая система имеет непосредственное отношение к эмоциям, как чувствам и настроениям, проявляющимся в поведении и реакциях
- 12. Повреждение миндалины у животных снижает адекватную подготовку автономной нервной системы к организации и реализации поведенческих ре-акций,
- 13. 10.3. ФУНКЦИИ НОВОЙ КОРЫ, ФУНКЦИОНАЛЬНОЕ ЗНАЧЕНИЕ СОМАТОСЕНСОРНЫХ И МОТОРНЫХ ЗОН КОРЫ БОЛЬШИХ ПОЛУШАРИЙ. Кора больших полушарий
- 14. Таламокортикальные пути проецируются на заднюю центральную извилину. Здесь имеется строгое соматотопическое деление. На верхние отделы этой
- 15. Вкусовая система проецируется в гиппокампальной извилине по соседству с обонятельной областью коры (поле 43). Первичная соматосенсорная
- 16. Рис. 14. Локализационная карта Клейста
- 17. Моторные области. Впервые Фритч и Гитциг (1870) показали, что раздражение передней центральной извилины мозга (поле 4)
- 18. Вторая особенность ассоциативной области коры заключается в способ-ности к пластическим перестройкам в зависимости от значимости поступаю-щей
- 19. 10.4. ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА: ОСОБЕННОСТИ СИМПАТИЧЕСКОГО И ПАРАСИМПАТИЧЕСКОГО ОТДЕЛОВ, ИХ ВЛИЯНИЯ. В обычных условиях внутренние органы
- 20. На органы, иннервируемые ВНС, перерезка передних корешков оказывает меньшее влияние, так как дегенерации подвергается только преганглионарное
- 21. Центры ВНС. Низшими центрами, относящимися к центральному отделу ВНС, являются парасимпатические ядра кранио-бульбарного и сакрального отделов,
- 22. Помимо прямых нервных влияний на деятельность внутренних органов, гипоталамус может оказывать на них воздействие и гуморальным
- 23. Функции вегетативной нервной системы. И.П.Павловым было сформулировано понятие о тройственном характере функциональных влияний, которые может оказывать
- 24. Универсальный характер симпатической иннервации. Симпатическая система иннервирует все внутренние органы. Кроме того, она иннервирует разные отделы
- 26. Скачать презентацию

По А. Р. Лурия, реальным анатомическим и функциональным образованием, включенным в

Моделью работы рано созревающего и еще не контролируемого корой паллидарного комплекса
Моделью работы рано созревающего и еще не контролируемого корой паллидарного комплекса
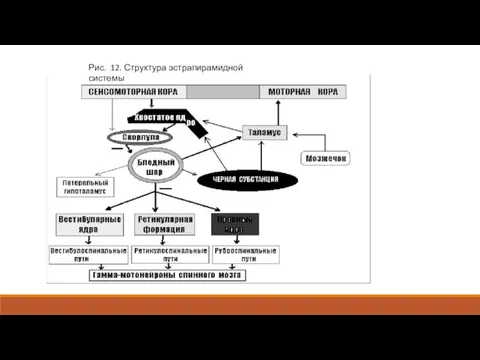
Рис. 12. Структура эстрапирамидной системы
Рис. 12. Структура эстрапирамидной системы

При поражении старого отдела экстрапирамидной системы возникает противоположная картина. У больных
При поражении старого отдела экстрапирамидной системы возникает противоположная картина. У больных

Пирамидная система (кортико-спинальный путь) начинается от моторных (крупных пирамидных) клеток Беца,
Пирамидная система (кортико-спинальный путь) начинается от моторных (крупных пирамидных) клеток Беца,

Третья группа волокон пирамидной системы, после частичного перекреста на уровне среднего
Третья группа волокон пирамидной системы, после частичного перекреста на уровне среднего

10.2. ЛИМБИЧЕСКАЯ СИСТЕМА: СТРУКТУРЫ И ФУНКЦИИ.
Лимбическая система. Передний мозг, интегративная деятельность
10.2. ЛИМБИЧЕСКАЯ СИСТЕМА: СТРУКТУРЫ И ФУНКЦИИ.
Лимбическая система. Передний мозг, интегративная деятельность

В настоящее время хорошо известны связи между структурами мозга, организующие круги,
В настоящее время хорошо известны связи между структурами мозга, организующие круги,

Лимбическая система имеет непосредственное отношение к эмоциям, как чувствам и настроениям,
Лимбическая система имеет непосредственное отношение к эмоциям, как чувствам и настроениям,


Повреждение миндалины у животных снижает адекватную подготовку автономной нервной системы к
Повреждение миндалины у животных снижает адекватную подготовку автономной нервной системы к

10.3. ФУНКЦИИ НОВОЙ КОРЫ, ФУНКЦИОНАЛЬНОЕ ЗНАЧЕНИЕ СОМАТОСЕНСОРНЫХ И МОТОРНЫХ ЗОН
10.3. ФУНКЦИИ НОВОЙ КОРЫ, ФУНКЦИОНАЛЬНОЕ ЗНАЧЕНИЕ СОМАТОСЕНСОРНЫХ И МОТОРНЫХ ЗОН

Таламокортикальные пути проецируются на заднюю центральную извилину. Здесь имеется строгое соматотопическое
Таламокортикальные пути проецируются на заднюю центральную извилину. Здесь имеется строгое соматотопическое

Вкусовая система проецируется в гиппокампальной извилине по соседству с обонятельной областью
Вкусовая система проецируется в гиппокампальной извилине по соседству с обонятельной областью
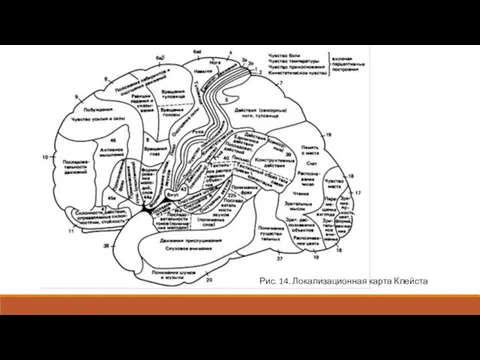
Рис. 14. Локализационная карта Клейста
Рис. 14. Локализационная карта Клейста

Моторные области. Впервые Фритч и Гитциг (1870) показали, что раздражение передней
Моторные области. Впервые Фритч и Гитциг (1870) показали, что раздражение передней

Вторая особенность ассоциативной области коры заключается в способ-ности к пластическим перестройкам
Вторая особенность ассоциативной области коры заключается в способ-ности к пластическим перестройкам

10.4. ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА: ОСОБЕННОСТИ СИМПАТИЧЕСКОГО И ПАРАСИМПАТИЧЕСКОГО ОТДЕЛОВ, ИХ ВЛИЯНИЯ.
10.4. ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА: ОСОБЕННОСТИ СИМПАТИЧЕСКОГО И ПАРАСИМПАТИЧЕСКОГО ОТДЕЛОВ, ИХ ВЛИЯНИЯ.

На органы, иннервируемые ВНС, перерезка передних корешков оказывает меньшее влияние, так
На органы, иннервируемые ВНС, перерезка передних корешков оказывает меньшее влияние, так

Центры ВНС. Низшими центрами, относящимися к центральному отделу ВНС, являются парасимпатические
Центры ВНС. Низшими центрами, относящимися к центральному отделу ВНС, являются парасимпатические

Помимо прямых нервных влияний на деятельность внутренних органов, гипоталамус может оказывать
Помимо прямых нервных влияний на деятельность внутренних органов, гипоталамус может оказывать

Функции вегетативной нервной системы. И.П.Павловым было сформулировано понятие о тройственном характере
Функции вегетативной нервной системы. И.П.Павловым было сформулировано понятие о тройственном характере

Универсальный характер симпатической иннервации. Симпатическая система иннервирует все внутренние органы. Кроме
Универсальный характер симпатической иннервации. Симпатическая система иннервирует все внутренние органы. Кроме
 Электрические колебания. Переменный ток
Электрические колебания. Переменный ток История туризма и гостеприимства
История туризма и гостеприимства Классный час Химки
Классный час Химки Мы граждане - России!!! Презентация.
Мы граждане - России!!! Презентация. Технология машинных швейных работ
Технология машинных швейных работ Методы расчета потерь электроэнергии для различных сетей
Методы расчета потерь электроэнергии для различных сетей Мусульманская правовая система
Мусульманская правовая система занятие
занятие Для Вас, выпускники: весна 2 класс
Для Вас, выпускники: весна 2 класс User Awareness and Practices
User Awareness and Practices Требования ФГОС ВО 3+ и варианты решения. Матрица компетенций
Требования ФГОС ВО 3+ и варианты решения. Матрица компетенций Корпоративное венчурные фонды, как новый субъект отношений в сфере интеллектуальной собственности
Корпоративное венчурные фонды, как новый субъект отношений в сфере интеллектуальной собственности Презентация к консультации для педагогов ГБОУ.Формирование знаний о правилах дорожного движения у детей дошкольного возраста.
Презентация к консультации для педагогов ГБОУ.Формирование знаний о правилах дорожного движения у детей дошкольного возраста. Воздействие сигнала и шума на линейный АД
Воздействие сигнала и шума на линейный АД Автоматический контроль качества
Автоматический контроль качества Гемоглобинопатии и их диагностика
Гемоглобинопатии и их диагностика Огонь-друг или враг? Диск
Огонь-друг или враг? Диск Психология малых групп
Психология малых групп Презентация Путешествие в страну Здоровья
Презентация Путешествие в страну Здоровья Презентация к классному часу Семья
Презентация к классному часу Семья Алексей Константинович Толстой (1817-1875)
Алексей Константинович Толстой (1817-1875) Asking for/Giving directions
Asking for/Giving directions Интерактивная игра Кино и книги
Интерактивная игра Кино и книги Культура. Культурное наследие
Культура. Культурное наследие Histology of the circulatory system
Histology of the circulatory system Общая программа мониторинга загрязнения почв
Общая программа мониторинга загрязнения почв Творческий проект Подарок маме
Творческий проект Подарок маме Должностные положения бухгалтерского, финансового и экономического отделов
Должностные положения бухгалтерского, финансового и экономического отделов