- Генетическая изменчивость

Содержание
- 2. Понятие «мутация» Термин мутация предложен голландским генетиком Гуго Де Фризом в 1901 году в результате экспериментов
- 3. Классификация мутаций По уровню дискретности генотипа: генные (изменение первичной структуры ДНК), хромосомные (изменение структуры хромосомы), геномные
- 4. Спонтанные мутации Спонтанные мутации возникают постоянно как у самоопыляющихся так и у перекрестноопыляющихся растений (у перекрестников
- 5. Типы мутагенов Физические а) электромагнитные излучения : рентгеновские лучи, гамма-лучи, ультрафиолетовый свет, инфракрасный свет б) корпускулярные
- 6. Химические а) ингибиторы и аналоги азотистых оснований ДНК и РНК: 5-бромурацил, 2-аминопурин б) алкилирующие агенты: иприт,
- 7. Генные мутации Ген – отрезок ДНК, который состоит из определенного числа пар нуклеотидов, кодирующий синтез полипептида.
- 8. Хромосомные мутации - аберрации Аберрации - изменения структуры хромосом, связанные с разрывами хромосом при воздействии тех
- 9. Механизмы возникновения основных типов аберраций: каждая из четырех типов аберраций может возникнуть как в результате разрыва
- 10. Геномные мутации - полиплоидия Явление изменения числа хромосом в клетках называется полиплоидией (термин предложен Т.Винклером в
- 11. Типы полиплоидии Автополиплоидия – кратное гаплоидному числу увеличение числа хромосом внутри одного вида Аллополиплоидия – увеличение
- 12. Пути возникновения полиплоидов Митотический (бесполый, митотическая полиплоидизация) - в результате нарушений митоза: в анафазе хроматиды не
- 13. Использование колхицина для увеличения плоидности растений Колхицин нарушает веретено деления делящихся клеток меристемы. Хромосомы не расходятся
- 14. Автополиплоидия Автополиплоиды, как правило, имеют увеличенные размеры ядер и клеток, что ведет к увеличению размеров листьев,
- 15. картофель (Solanum) 12, 24, 36, 48, 60, 72, 96, 108, 144 щавель (Rumex) 20, 40, 60,
- 16. Цветки тетраплоидной (4х) и диплоидной (2х) петунии Автополиплоидия Плоды октоплоидной (8х) и диплоидной (2х) земляники Диплоидный
- 17. АВТОПОЛИПЛОИДИЯ – ФОРМИРОВАНИЕ ТРИПЛОИДНЫХ ОРГАНИЗМОВ При скрещивании автотетраплоидов с диплоидами возникают триплоидные стерильные организмы, но иногда
- 18. Наследование при автоплоидии У автотетраполидов имеются 5 возможных генотипов по 1 гену (у диплоидов – 3),
- 19. Аллоплоидия Аллополиплоидия – увеличение числа хромосом за счет объединения геномов разных видов
- 20. Сравнение автополиплоидии, аллоплоидии и аллополиплоидии
- 21. Получение искусственного вида Triticale
- 22. Анеуплоидия Образование анеуплоидных гамет в результате нерасхождения хромосом при первом и втором мейотическом делении Анеуплоидия –
- 23. Анеуплоидия Нерасхождение и потеря хромосом во время митотического деления соматических клеток Нерасхождение в митозе приводит к
- 24. Колосья нуллисомиков мягкой пшеницы сорта «Чайниз Спринг»
- 25. Семенные коробочки трисомиков дурмана по каждой из 12 хромосом гаплоидного набора
- 26. Гаплоидия Гаплоид – организм, имеющий в соматических клетках полный для данного вида набор негомологичных хромосом (n);
- 27. Узел с 1 бутоном женского цветка и несколькими мужскими Бутоны огурца, от самого маленького (4,6мм) до
- 28. Стадии развития микроспор (окрашивание ацетокармином и красителем DAPI) Тетрады микроспор (пыльник 2,51 мм, чашелистики 7,65 мм,
- 29. Стадии развития микроспор (окрашивание ацетокармином и красителем DAPI) Средняя одноядерная стадия развития микроспор (количество ядер в
- 30. Стадии развития микроспор (окрашивание ацетокармином и красителем DAPI) Двухъядерная стадия развития микроспор (количество ядер в ацетокармине
- 31. Стадии развития каллуса. А – желтоватый, прозрачный эмбриоид, возникающий из эмбрионального каллуса через 30 дней после
- 33. Гаплоидное (а) и диплоидное (б) растения томата Особенности гаплоидных растений По фенотипу гаплоиды похожи на диплоиды,
- 34. Инбридинг и гетерозис. Получение потомства от скрещивания родственных между собой особей называется инбридингом. В результате самоопыления,
- 35. Гипотеза доминирования основана на том, что большинство рецессивных генов отрицательно влияет на организм, поэтому накопление у
- 36. D. Shull и E.East и Шелл, Ист и Хейс развивали генетическую гипотезу сверхдоминирования, согласно которой доминантные
- 38. Скачать презентацию

Понятие «мутация»
Термин мутация предложен голландским генетиком Гуго Де Фризом в 1901
Понятие «мутация»
Термин мутация предложен голландским генетиком Гуго Де Фризом в 1901

Классификация мутаций
По уровню дискретности генотипа:
генные (изменение первичной структуры ДНК),
хромосомные
Классификация мутаций
По уровню дискретности генотипа:
генные (изменение первичной структуры ДНК),
хромосомные

Спонтанные мутации
Спонтанные мутации возникают постоянно как у самоопыляющихся так и у
Спонтанные мутации
Спонтанные мутации возникают постоянно как у самоопыляющихся так и у
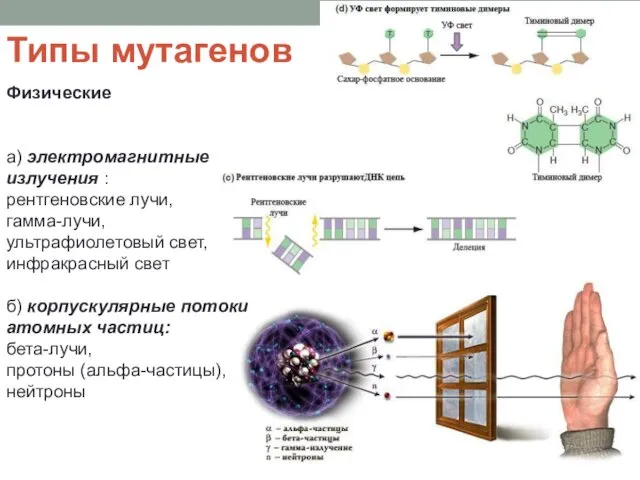
Типы мутагенов
Физические
а) электромагнитные излучения :
рентгеновские лучи,
гамма-лучи,
ультрафиолетовый свет,
Типы мутагенов
Физические
а) электромагнитные излучения :
рентгеновские лучи,
гамма-лучи,
ультрафиолетовый свет,
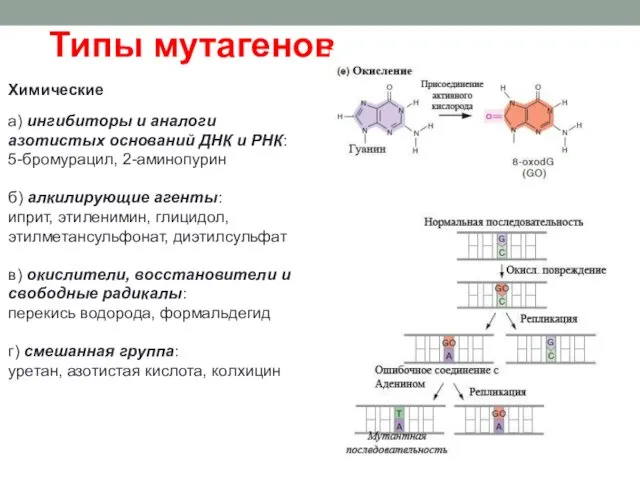
Химические
а) ингибиторы и аналоги азотистых оснований ДНК и РНК:
5-бромурацил,
Химические
а) ингибиторы и аналоги азотистых оснований ДНК и РНК:
5-бромурацил,

Генные мутации
Ген – отрезок ДНК, который состоит из определенного числа пар
Генные мутации
Ген – отрезок ДНК, который состоит из определенного числа пар

Хромосомные мутации - аберрации
Аберрации - изменения структуры хромосом, связанные с разрывами
Хромосомные мутации - аберрации
Аберрации - изменения структуры хромосом, связанные с разрывами
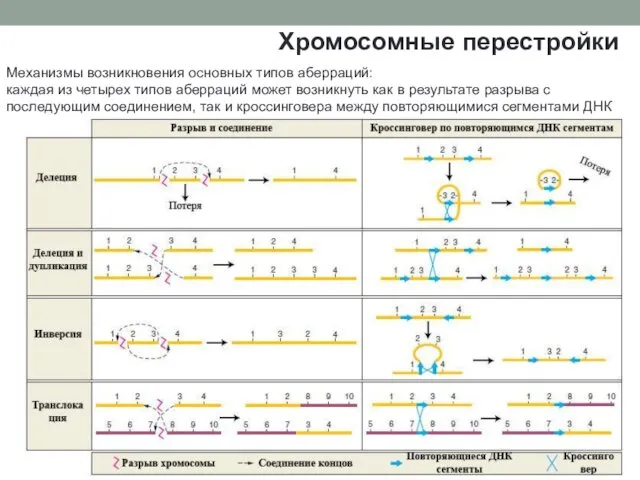
Механизмы возникновения основных типов аберраций:
каждая из четырех типов аберраций может возникнуть
Механизмы возникновения основных типов аберраций: каждая из четырех типов аберраций может возникнуть

Геномные мутации - полиплоидия
Явление изменения числа хромосом в клетках называется полиплоидией
Геномные мутации - полиплоидия
Явление изменения числа хромосом в клетках называется полиплоидией

Типы полиплоидии
Автополиплоидия – кратное гаплоидному числу увеличение числа хромосом внутри одного
Типы полиплоидии
Автополиплоидия – кратное гаплоидному числу увеличение числа хромосом внутри одного

Пути возникновения полиплоидов
Митотический (бесполый, митотическая полиплоидизация) - в результате нарушений митоза:
Пути возникновения полиплоидов
Митотический (бесполый, митотическая полиплоидизация) - в результате нарушений митоза:
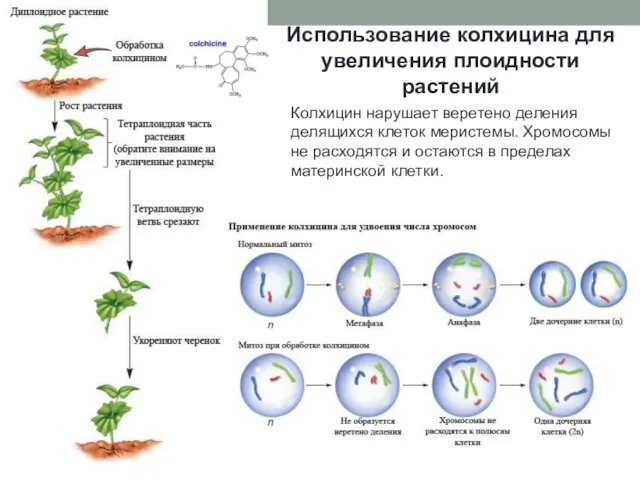
Использование колхицина для увеличения плоидности растений
Колхицин нарушает веретено деления делящихся клеток
Использование колхицина для увеличения плоидности растений
Колхицин нарушает веретено деления делящихся клеток

Автополиплоидия
Автополиплоиды, как правило, имеют увеличенные размеры ядер и клеток, что ведет
Автополиплоидия
Автополиплоиды, как правило, имеют увеличенные размеры ядер и клеток, что ведет

картофель (Solanum) 12, 24, 36, 48, 60, 72, 96, 108, 144
щавель
картофель (Solanum) 12, 24, 36, 48, 60, 72, 96, 108, 144
щавель

Цветки тетраплоидной (4х) и диплоидной (2х) петунии
Автополиплоидия
Плоды октоплоидной (8х) и диплоидной
Цветки тетраплоидной (4х) и диплоидной (2х) петунии
Автополиплоидия
Плоды октоплоидной (8х) и диплоидной
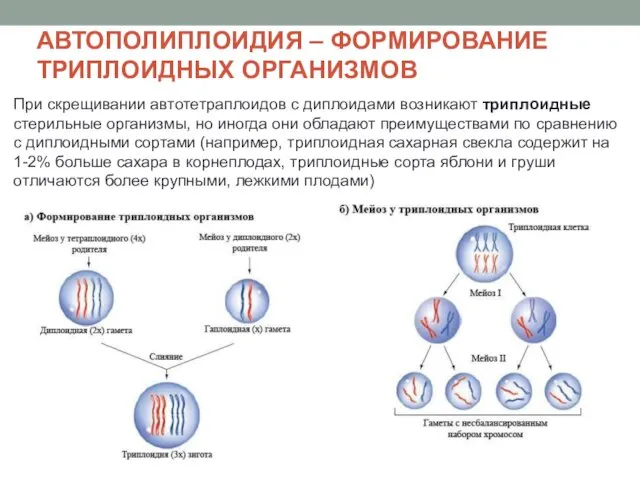
АВТОПОЛИПЛОИДИЯ – ФОРМИРОВАНИЕ ТРИПЛОИДНЫХ ОРГАНИЗМОВ
При скрещивании автотетраплоидов с диплоидами возникают триплоидные
АВТОПОЛИПЛОИДИЯ – ФОРМИРОВАНИЕ ТРИПЛОИДНЫХ ОРГАНИЗМОВ
При скрещивании автотетраплоидов с диплоидами возникают триплоидные

Наследование при автоплоидии
У автотетраполидов имеются 5 возможных генотипов по 1
Наследование при автоплоидии
У автотетраполидов имеются 5 возможных генотипов по 1
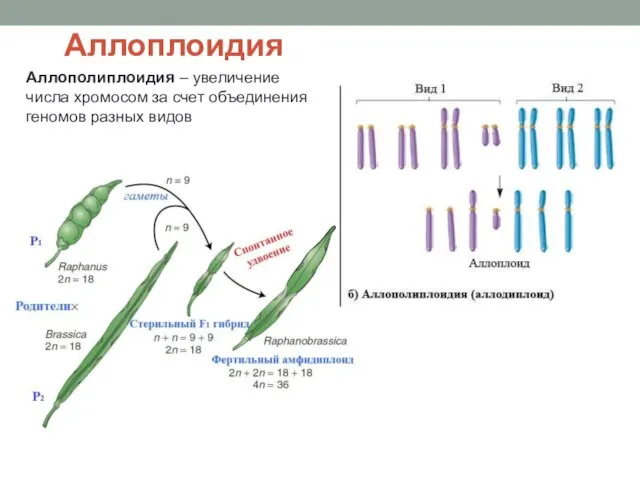
Аллоплоидия
Аллополиплоидия – увеличение числа хромосом за счет объединения геномов разных видов
Аллоплоидия
Аллополиплоидия – увеличение числа хромосом за счет объединения геномов разных видов
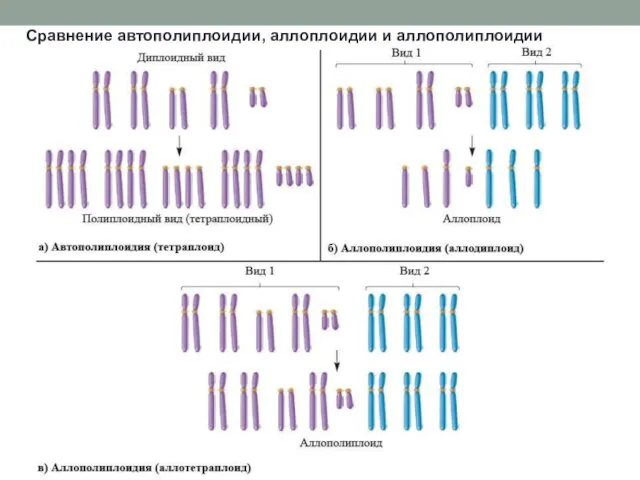
Сравнение автополиплоидии, аллоплоидии и аллополиплоидии
Сравнение автополиплоидии, аллоплоидии и аллополиплоидии
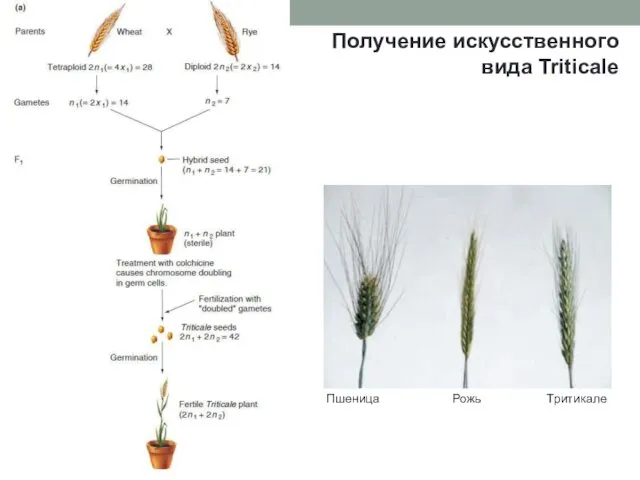
Получение искусственного вида Triticale
Получение искусственного вида Triticale
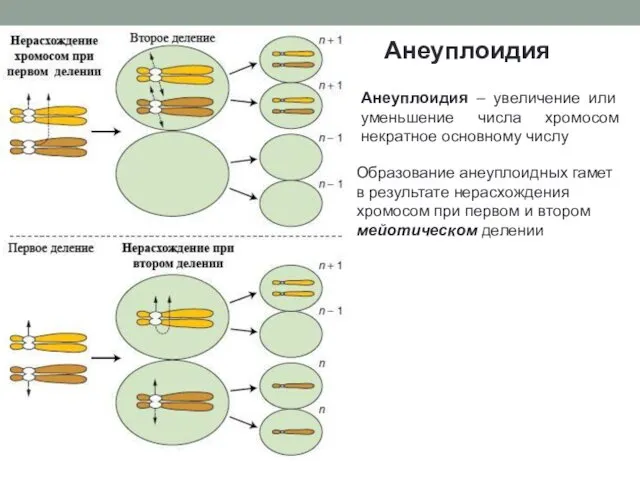
Анеуплоидия
Образование анеуплоидных гамет в результате нерасхождения хромосом при первом и втором
Анеуплоидия
Образование анеуплоидных гамет в результате нерасхождения хромосом при первом и втором
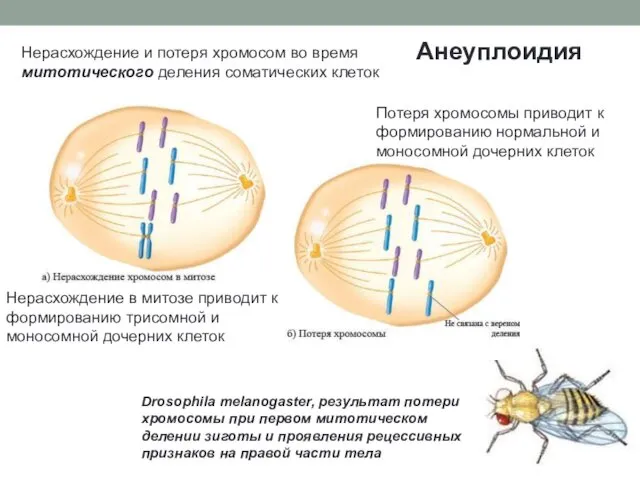
Анеуплоидия
Нерасхождение и потеря хромосом во время митотического деления соматических клеток
Нерасхождение в
Анеуплоидия
Нерасхождение и потеря хромосом во время митотического деления соматических клеток
Нерасхождение в

Колосья нуллисомиков мягкой пшеницы сорта «Чайниз Спринг»
Колосья нуллисомиков мягкой пшеницы сорта «Чайниз Спринг»

Семенные коробочки трисомиков дурмана по каждой из 12 хромосом гаплоидного набора
Семенные коробочки трисомиков дурмана по каждой из 12 хромосом гаплоидного набора

Гаплоидия
Гаплоид – организм, имеющий в соматических клетках полный для данного вида
Гаплоидия
Гаплоид – организм, имеющий в соматических клетках полный для данного вида

Узел с 1 бутоном женского
цветка и несколькими
мужскими
Бутоны огурца, от самого
Узел с 1 бутоном женского
цветка и несколькими
мужскими
Бутоны огурца, от самого
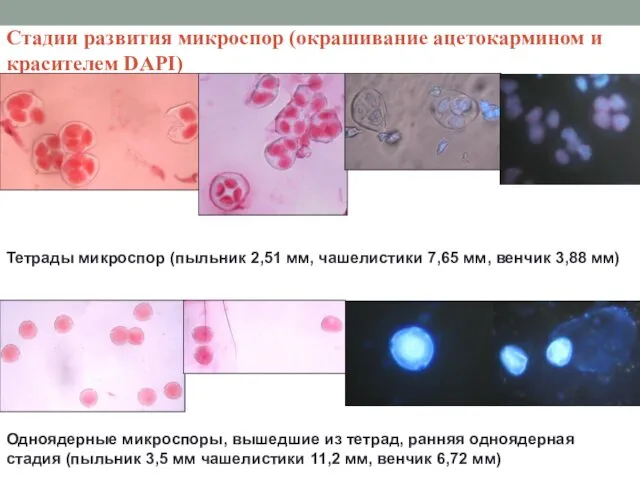
Стадии развития микроспор (окрашивание ацетокармином и красителем DAPI)
Тетрады микроспор (пыльник 2,51
Стадии развития микроспор (окрашивание ацетокармином и красителем DAPI)
Тетрады микроспор (пыльник 2,51
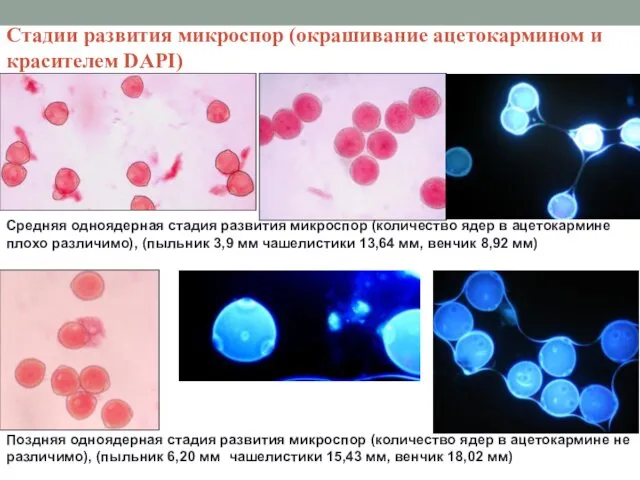
Стадии развития микроспор (окрашивание ацетокармином и красителем DAPI)
Средняя одноядерная стадия развития
Стадии развития микроспор (окрашивание ацетокармином и красителем DAPI)
Средняя одноядерная стадия развития
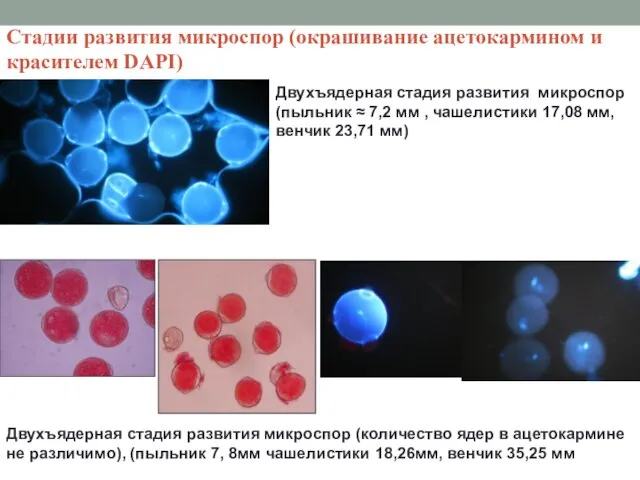
Стадии развития микроспор (окрашивание ацетокармином и красителем DAPI)
Двухъядерная стадия развития микроспор
Стадии развития микроспор (окрашивание ацетокармином и красителем DAPI)
Двухъядерная стадия развития микроспор

Стадии развития каллуса.
А – желтоватый, прозрачный эмбриоид, возникающий из эмбрионального каллуса
Стадии развития каллуса.
А – желтоватый, прозрачный эмбриоид, возникающий из эмбрионального каллуса


Гаплоидное (а) и диплоидное (б) растения томата
Особенности гаплоидных растений
По фенотипу гаплоиды
Гаплоидное (а) и диплоидное (б) растения томата
Особенности гаплоидных растений
По фенотипу гаплоиды

Инбридинг и гетерозис.
Получение потомства от скрещивания родственных между собой особей называется
Инбридинг и гетерозис.
Получение потомства от скрещивания родственных между собой особей называется

Гипотеза доминирования основана на том, что большинство рецессивных генов отрицательно влияет на
Гипотеза доминирования основана на том, что большинство рецессивных генов отрицательно влияет на

D. Shull и E.East и Шелл, Ист и Хейс развивали генетическую гипотезу
D. Shull и E.East и Шелл, Ист и Хейс развивали генетическую гипотезу
 Маркировка чугуна
Маркировка чугуна Презентация ЦИПР
Презентация ЦИПР Презентация аналитического отчета за 2014-2015 год
Презентация аналитического отчета за 2014-2015 год Электрическое сопротивление проводника. Удельное сопротивление
Электрическое сопротивление проводника. Удельное сопротивление научно-практическая конференция
научно-практическая конференция презентация Возникновение жизни (4)
презентация Возникновение жизни (4) Вирусы
Вирусы Аутсорсинг
Аутсорсинг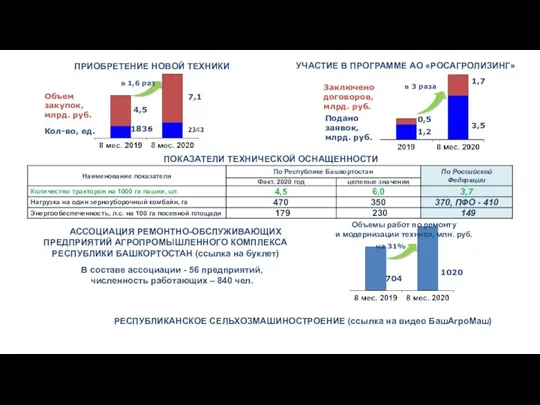 Программа АО Росагролизинг. Приобретение новой техники
Программа АО Росагролизинг. Приобретение новой техники Презентация к уроку Страны мира 7 класс
Презентация к уроку Страны мира 7 класс Жер бедері
Жер бедері Оригами. Проект
Оригами. Проект Снеговик. Новогодняя поделка
Снеговик. Новогодняя поделка 9 мая - День Победы. Презентация к лассному часу
9 мая - День Победы. Презентация к лассному часу Изобразительное искусство России начала XX века
Изобразительное искусство России начала XX века Коллективный проект, технология, 3 класс, Подарок ветерану
Коллективный проект, технология, 3 класс, Подарок ветерану Презентация по методической теме: Балаларда туган җиребезгә мәхәббәт, аның табигатенә карата сакчыл караш тәрбияләү һәм экологик тәрбия бирү 2011-2015 год
Презентация по методической теме: Балаларда туган җиребезгә мәхәббәт, аның табигатенә карата сакчыл караш тәрбияләү һәм экологик тәрбия бирү 2011-2015 год Перпендикуляр и наклонная
Перпендикуляр и наклонная Презентация Школа сегодня и вчера
Презентация Школа сегодня и вчера Рахіт. Етіологія, патогенез, клініка, діагностика, лікування, профілактика
Рахіт. Етіологія, патогенез, клініка, діагностика, лікування, профілактика Автоматизированное рабочее место врача-терапевта (АРМ)
Автоматизированное рабочее место врача-терапевта (АРМ) Фронтальные одноковшовые погрузчики
Фронтальные одноковшовые погрузчики Дозирующие устройства жидкости
Дозирующие устройства жидкости Системы впрыска бензина
Системы впрыска бензина Dark wash-Finish
Dark wash-Finish Градусная сетка. Географические координаты
Градусная сетка. Географические координаты Экскурсия
Экскурсия Расчет радиовещательного приемника высшего класса
Расчет радиовещательного приемника высшего класса