- Иммунология. В и Т-лимфоциты и кооперация клеток в иммунном ответе. (Лекция 3)

Содержание
- 2. повторяем ….Основные АПК
- 3. ДИФФЕРЕНЦИРОВОЧНЫЕ АНТИГЕНЫ ЛИМФОЦИТОВ
- 4. клеточные ФАКТОРЫ ПРИОБРЕТЕННОГО ИММУНИТЕТА Т-ЛИМФОЦИТЫ В- ЛИМФОЦИТЫ CD*4 CD*8 дендритные клетки плазмоциты Клетки памяти
- 5. Лейкоциты всех типов происходят от единой костномозговой полипотентной стволовой кроветворной клетки, популяция которой является самоподдерживающейся. Направление
- 6. Во вторичных лимфоидных органах - Т- и В-лимфоциты впервые контактируют с чужеродными для организма антигенами. Происходит
- 7. Т-лимфоциты Т- лимфоциты – сложная по составу группа клеток. На их долю приходится 75% всей популяции
- 8. Т- лимфоциты Функции Т- лимфоцитов : - распознают процессированный и представленный на поверхности антиген-представляющих клеток (АПК)
- 9. В тимусе (thymus) проходит лимфопоэз значительной части T-лимфоцитов В каждой дольке тимуса выделяют 2 зоны: по
- 10. Роль гуморальных и клеточных факторов тимуса Звездчатые эпителиальные клетки (клетки- няньки): секретируют растворимые тимические (или тимусные)
- 11. Происхождение и созревание Т- лимфоцитов. Родоначальницей всех клеток крови является единая стволовая клетка костного мозга. Образование
- 12. В зависимости от того какую молекулу АГ+МНС1 или МНС2 «узнает» Т-лимфоцит (его TCR) он дифференцируется в
- 13. Рецепторы Т-лимфоцита
- 14. Антигенраспознающие участки рецепторов В лимфоцитов (ВCR) и рецепторов Т-лимфоцитов (TCR) Антиген-распознающие участки рецепторов как Т, так
- 15. Основные рецепторы на примере Т-хелпера Существует два типа TCR- Т- клеточн.рецептор, каждый из которых присуствует на
- 16. Популяции лимфоцитов
- 17. Т-лимфоциты Тимус Контакт с антигеном Th-fn T- reg
- 18. T-цитотоксические Т-киллеры (CTL или CD8+)
- 19. T-цитотоксические лимф. (кратко) Поверхностный маркер – СD3 (сигнальный комплекс, связанный с ТСР), CD8 (рецептор к МНС
- 20. Т-киллер атакует клетку, на поверхности которой экспрессирован ассоциированный с MHC-I антиген, к которому комплементарен Т- клеточный
- 21. Т-хелперы и их роль в иммунном ответе
- 22. Th0-лимфоциты (наивные) Поверхностный маркер – СD3, CD4 (рецептор к МНС II) Происхождение – из клеток-предшественников в
- 23. T-хелперы CD4+ В зависмости от характера и презентации антигена (кто презентировал - макрофаги , В-клетки и
- 24. Парадигма о Т-хелперах до 2012 года Публикация : CD4 T cells: fates, functions, and faults Jinfang
- 25. Парадигма о Т-хелперах после 2012 года Активация разных эффекторных клеток цитокинами разных субпопуляций Т-хелперов: ЗАЩИТА /
- 26. Т-лимфоциты-участники адаптивного (антигенспецифического) ответа
- 27. Дополнительный матерал- вне рамок курса
- 28. Другие субпопуляции Т-хелперов (материал не входит в рамки программы курса- дополнительно) Th17 Если в микроокружении наивного
- 29. Functions of TH17 cells Fig. 10-10
- 30. Субпопуляции Т-хелперов: регуляторные клетки - T reg (CD4+CD25+FOXP3 клетки) «Т- реги» или Т-супрессоры Эти клетки способны
- 31. Развитие натуральной регуляторной Т-клетки в тимусе (Treg:CD4+CD25+FOXP3 )
- 32. Окончание дополнительного материала
- 33. Иммунологическая память. Субопуляции клеток памяти
- 34. Иммунологическая память. Субопуляции клеток памяти Иммунологическая память – феномен более раннего и более сильного иммунного ответа
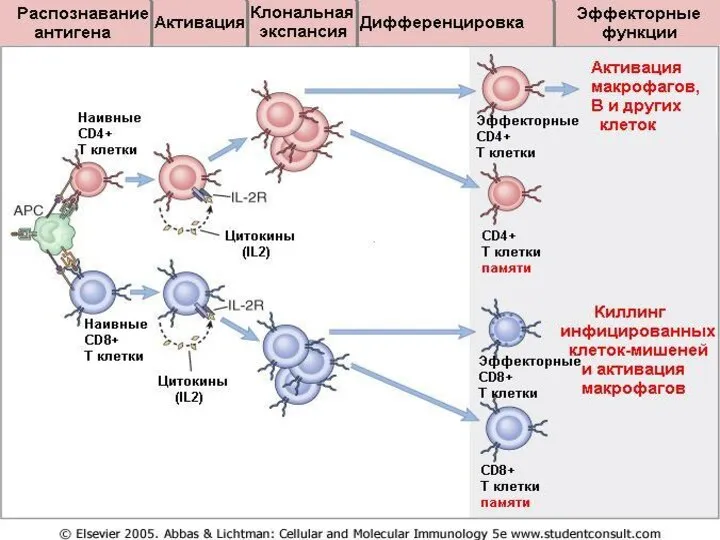
- 35. Этапы иммунного ответа распознавание антигена клональная экспансия Т- и В-лимфоцитов. дифференцировка эффекторных клеток продукция антител активация
- 36. Основные субпопуляции Т-клеток памяти в лимфоидной ткани локализуются Т клетки центральной памяти Т клетки центральной памяти
- 37. Основные субпопуляции Т-клеток памяти: Т-клетки эффекторной памяти (ТЕМ) Т клетки эффекторной памяти ТЕМ- являются терминально дифференцированными
- 38. Миграция Т-клеток памяти в ткани: кожу, легкие, кишечник лимфоузел посткапиллярные венулы кожа легкие жкт Тн 0
- 39. Число Т-клеток памяти в организме увеличивается с возрастом
- 40. Что способствует формированию долговременной памяти на патоген? Свойства самого антигена (иммуногенность), а также свойства иммунной системы
- 41. ФАКТОРЫ ПРИОБРЕТЕННОГО ИММУНИТЕТА Т-ЛИМФОЦИТЫ В- ЛИМФОЦИТЫ дендритные клетки
- 42. клеточные ФАКТОРЫ ПРИОБРЕТЕННОГО ИММУНИТЕТА В- ЛИМФОЦИТЫ В1-клетки В2 клетки плазмоциты Клетки памяти
- 43. В-лимфоциты (bursa fabricius) Y Y клетки иммунной системы, через которые реализуются свойства гуморального иммунного ответа (
- 44. Характеристика В-клеток: функции: способность дифференцироваться в плазматические клетки и продуцировать антитела; выступать в роли антигенпредставляющих клеток
- 45. Типы В-клеток В1-лимфоциты отвечают на Т независимые антигены (нативный или растворимый антиген), не нуждаются в помощи
- 46. Рецепторы B-лимфоцитов узнающим антиген, является IgM/D рецептор. Это мономерный иммуноглобулин. Число молекул рецептора достигает 10 -
- 47. Рецепторы В-лимфоцитов BCR CD19 - самый ранний маркер В-клеток, . CD20 - маркер В-клеток, характеризующий поздние
- 48. B-клеточный распознающий рецептор (BCR) Состоит из мембранной формы Ig D или Ig M в комплексе с
- 49. Онтогенез В-лимфоцитов Предшественники В-лимфоцитов обнаружены в островках гемопоэтической ткани эмбриональной печени на 8-9-й неделе эмбрионального развития
- 50. Плазмоцит
- 51. Плазматические клетки (“МАМы антител”) нет в кровотоке локализация – ЛУ, селезенка и лимфоидные образования в слизистых
- 52. Сопоставление характеристик В-лимфоцита и плазматической клетки уровень экспрессии соматические Ig MHC II Ig секреция мутации Высокий
- 53. Центр размножения ЛУ(зародышевые центры) Особая область в лимфатических узлах или селезенке, в которой обнаруживается интенсивная пролиферация
- 54. Зародышевые центры – ЗЦ (дифференцировка и селекция В-лимфоцитов) ТЕМНАЯ ЗОНА: В-л связавшие АГ и получившие дополнительный
- 55. Рециркулирующие В – клетки встречаются со «ИСКОМЫМ» антигеном в лимфоузле
- 56. роль Tfh (фолликулярных Т хелперов) На территории лимфоидного фолликула в ЛУ под контролем Tfh происходит :
- 57. Взаимодействие В-клеток с микроокружением Итог: ОТБОР КЛЕТОК АНТИТЕЛОПРОДУЦЕНТОВ 1. гибель клеток несущих низкоафинный рецептор ИГ или
- 58. Первичный иммунный ответ Плазматические клетки при первичном ответе синтезируют антитела с неизмененным, не прошедшим гиперсоматической мутации
- 59. Вторичный иммунный ответ (повторное попадание антигена) Под контролем Т-хелперов фолликулярных происходит: быстрая дифференцировка клеток памяти- В
- 60. Цитокины (смотри лекцию №2)
- 61. Цитокины – небольшие белки (8-80 кДа), действующие аутокринно или паракринно. Виды: Интерлейкины Интерфероны Колониестимулирующие факторы Факторы
- 62. Функции цитокинов регуляция гемопоэза регуляция иммунного ответа и воспалительных процессов участие в ангиогенезе апоптоз хемотаксис эмбриогенез
- 63. Взаимодействие клеток иммунной системы в ходе иммунного ответа(общие закономерности) КООПЕРАЦИЯ КЛЕТОК В ИММУННОМ ОТВЕТЕ
- 64. Иммунный синапс – основа кооперации клеток Синапс – область соприкосновения клеток, где происходит обмен сигнальными молекулами.
- 65. СИГНАЛ 1 (антигенное распознавание): группа взаимодействующих молекул на АПК (макрофаг или ДК) и Т лимфоците, обеспечивающих
- 66. «синапс»: В лимфоцит –T-хелпер IL21R IL-21 IL21R CXCR5 CXCL13 CXCL13 PD-1 PD-1L
- 67. Отсутствие экспрессии костимулирующих молекул антгенпрезетрующими клетками приводит к тому, что иммунный ответ не развивается -это является
- 68. Значение костимуляции для активации Т-клеток Fig. 9-3 Анергия и гибель Выживание клона клеток и пролиферация
- 69. Роль CD40 рецептора активации T клеток Fig. 9-6 Выживание клона клеток и пролиферация и дифференцировка
- 70. Это интересно… Помимо стимулирующих воздействий в иммунной системе предусмотрены и подавляющие воздействия. Рецептор PD1 и его
- 71. Клеточный тип иммунного ответа КООПЕРАЦИЯ КЛЕТОК В ИММУННОМ ОТВЕТЕ
- 72. Клеточный тип иммунного ответа ( инициаторы: макрофаги и дендр. клетки) Переход Th 0 в Th 1
- 73. Клеточный тип иммунного ответа (ИО) Основные цитокины, определяющие тип иммунного ответа производятся АПК (ДК). Основные продуценты
- 74. экспрессия β и γ цепей IL-2 рецептора, но не α цепи в обычном состоянии рецептор к
- 75. Клеточный тип иммунного ответа (ИО)
- 76. Этапы клеточного типа иммунного ответа (ИО) Происходит взаимодействие: комплекса MHC I I +антигенный пептид на поверхности
- 77. Общая схема клеточного им ответа
- 79. Клеточный тип иммунного ответа(схема)
- 80. Гуморальный тип иммунного ответа КООПЕРАЦИЯ КЛЕТОК В ИММУННОМ ОТВЕТЕ
- 81. Гуморальный тип иммунного ответа ( инициаторы: В-лимфоциты и дендр. клетки) Переход Th 0 в Th fn,
- 82. Основные «участники» синапсов в зависимости от типа и вида распознаваемого Аг это либо : Дуэт :В-лимфоцит
- 83. В зависимости от состава антигена выделяю ТИМУСзависимые АГ и Тимус не зависимые
- 84. Тимусзависимые АГ Нужна помощь хелперов для В-лимфоцитов для распознавания АГ
- 85. Тимусзависимые АГ Антигены, которым необходима помощь со стороны Т-клеток, получили название тимусзависимых антигенов. При адекватном включении
- 86. Взаимодействие между Т и В клетками это двунаправленный процесс (костимуляция) Th продуцирует ИЛ 21, 4, 6
- 87. T клетки помогают B клеткам сигнал 1 антиген и рецептор к антигену 1. T клеточный рецептор
- 88. Тимусзависимые АГ Зачем такие сложности????? Если В-клетки взаимодействовали с антигеном (АГ), но не получили помощи со
- 89. Тимусзависимые АГ для распознавания АГ хелпер не нужен В-лимфоцит справится САМ в одиночку!
- 90. Распознавание нативных антигенов лимфоцитами: Нативный –т.е. природный, не расщепленный антиген, способны самостоятельно распознавать только В лимфоциты
- 91. Тимуснезависимые АГ без участия хелперов Например АГ с митогенным участком (МУ), который формирует второй сигнал для
- 92. Первичный гуморальный ответ на тимуснезависимые АГ (Тi)немного слабее, чем на Т- зависимые АГ , но достигает
- 93. T клеточная помощь - сигнал 2 Сигналы1 и 2 повышают предотвращают апоптоз В- лимфоцита сигналы 1
- 94. Роль цитокинов в гуморальном иммунитете
- 95. Этапы гуморального ИО Наивные В- клетки сами распознают растворимый антиген (АГ) (без участия макрофага) или связанный
- 96. Общая схема гуморалного иммунного ответа (Т-независимые АГ)
- 97. Общая схема гуморалного иммунного ответа (Т зависимые АГ)
- 98. Фазы развития гуморального иммунного ответа По Ярилину А.А. Учебник ГОЭТАР 2010
- 99. Схема гуморального иммунного ответа Tfn IL-21,6 IL-21
- 100. КООПЕРАЦИЯ КЛЕТОК В ИММУННОМ ОТВЕТЕ (общая схема)
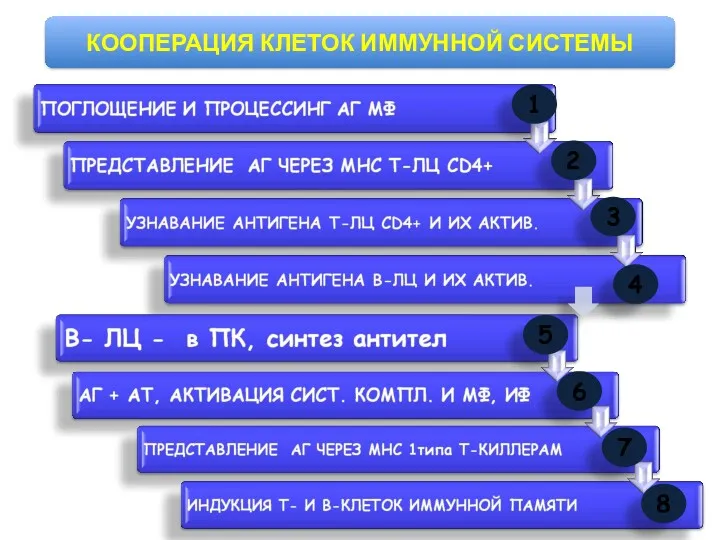
- 101. КООПЕРАЦИЯ КЛЕТОК ИММУННОЙ СИСТЕМЫ 1 2 3 4 5 6 7 8
- 103. Скачать презентацию
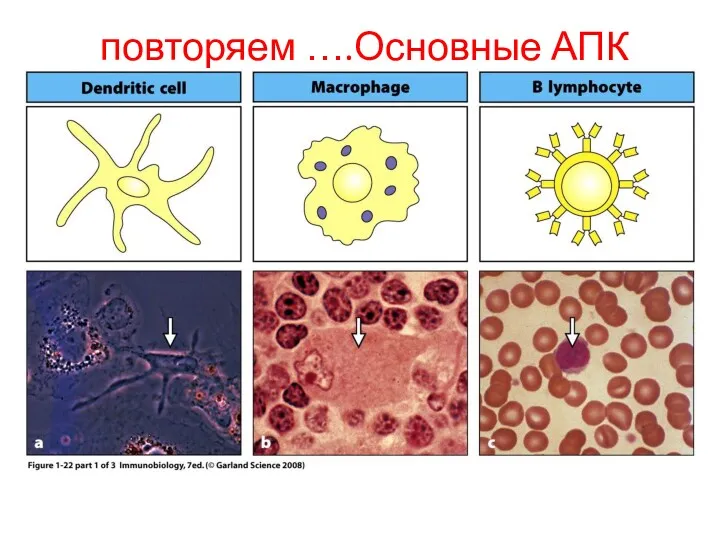
повторяем ….Основные АПК
повторяем ….Основные АПК
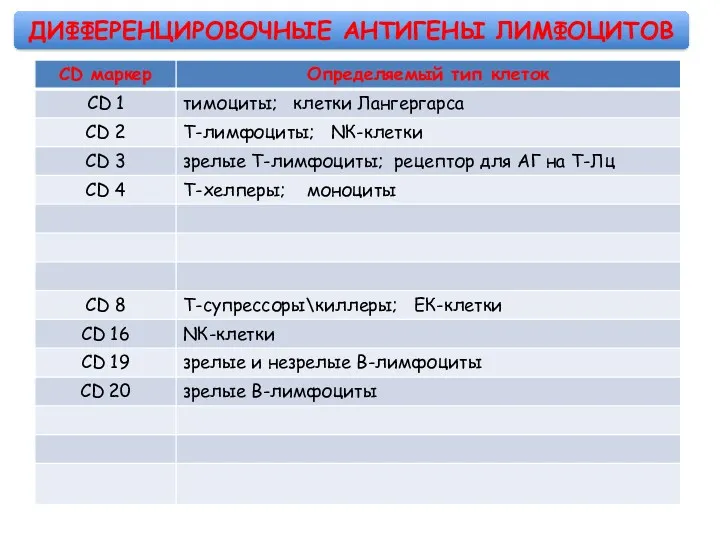
ДИФФЕРЕНЦИРОВОЧНЫЕ АНТИГЕНЫ ЛИМФОЦИТОВ
ДИФФЕРЕНЦИРОВОЧНЫЕ АНТИГЕНЫ ЛИМФОЦИТОВ
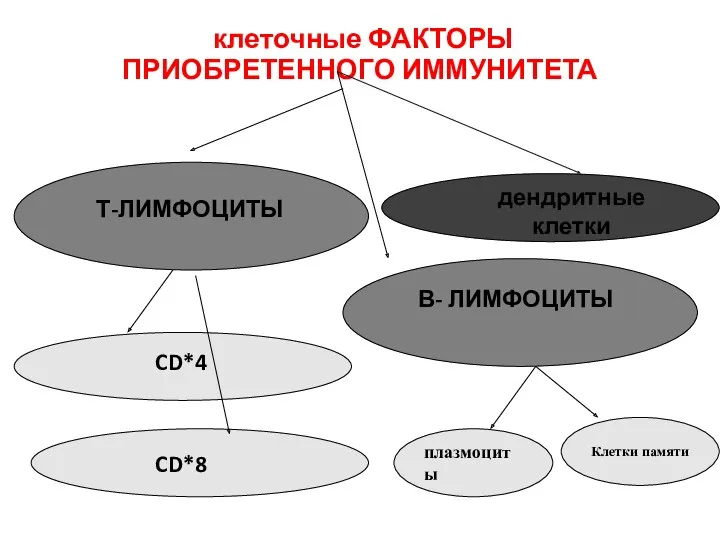
клеточные ФАКТОРЫ
ПРИОБРЕТЕННОГО ИММУНИТЕТА
Т-ЛИМФОЦИТЫ
В- ЛИМФОЦИТЫ
CD*4
CD*8
дендритные клетки
плазмоциты
Клетки памяти
клеточные ФАКТОРЫ
ПРИОБРЕТЕННОГО ИММУНИТЕТА
Т-ЛИМФОЦИТЫ
В- ЛИМФОЦИТЫ
CD*4
CD*8
дендритные клетки
плазмоциты
Клетки памяти

Лейкоциты всех типов происходят от единой костномозговой полипотентной стволовой кроветворной клетки,
Лейкоциты всех типов происходят от единой костномозговой полипотентной стволовой кроветворной клетки,

Во вторичных лимфоидных органах - Т- и В-лимфоциты впервые контактируют с
Во вторичных лимфоидных органах - Т- и В-лимфоциты впервые контактируют с

Т-лимфоциты
Т- лимфоциты – сложная по составу группа клеток.
На их долю
Т-лимфоциты
Т- лимфоциты – сложная по составу группа клеток.
На их долю

Т- лимфоциты
Функции Т- лимфоцитов :
- распознают процессированный и представленный
Т- лимфоциты
Функции Т- лимфоцитов :
- распознают процессированный и представленный
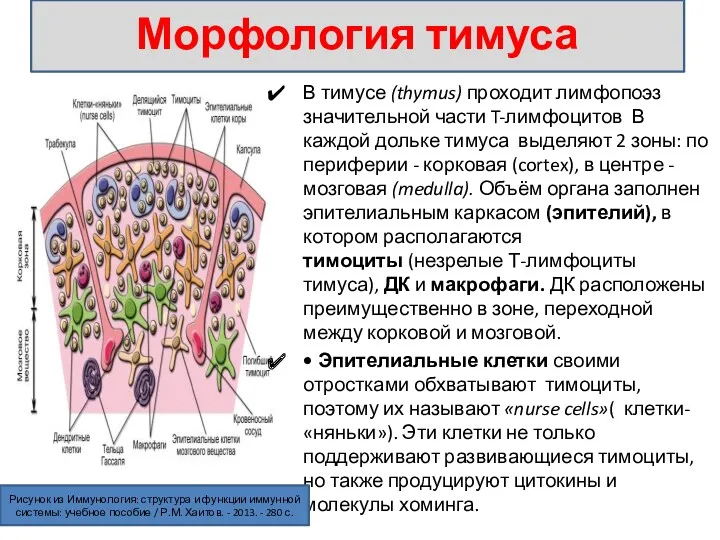
В тимусе (thymus) проходит лимфопоэз значительной части T-лимфоцитов В каждой дольке тимуса выделяют
В тимусе (thymus) проходит лимфопоэз значительной части T-лимфоцитов В каждой дольке тимуса выделяют

Роль гуморальных и клеточных факторов тимуса
Звездчатые эпителиальные клетки (клетки- няньки): секретируют
Роль гуморальных и клеточных факторов тимуса
Звездчатые эпителиальные клетки (клетки- няньки): секретируют
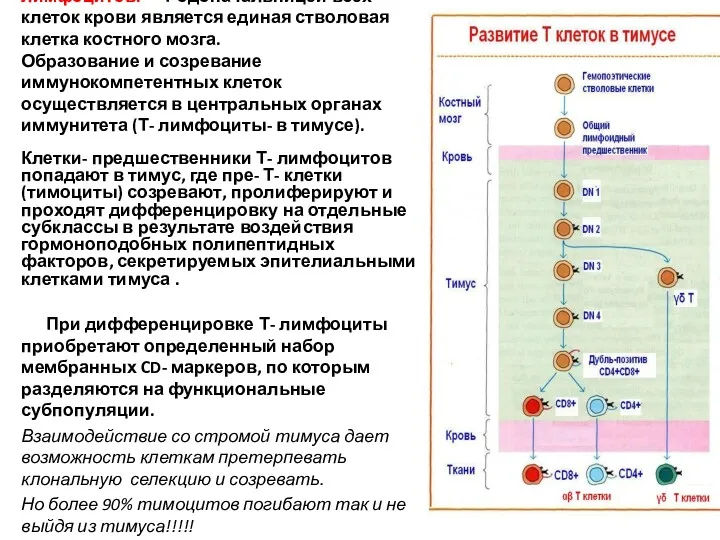
Происхождение и созревание Т- лимфоцитов. Родоначальницей всех клеток крови является единая
Происхождение и созревание Т- лимфоцитов. Родоначальницей всех клеток крови является единая
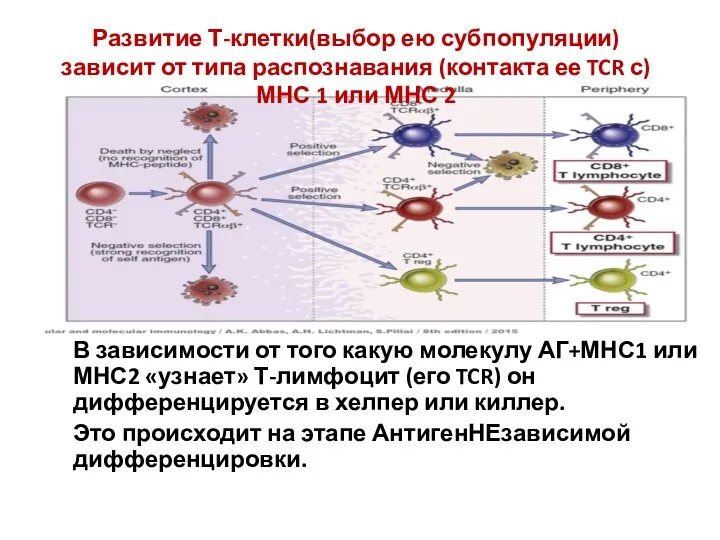
В зависимости от того какую молекулу АГ+МНС1 или МНС2 «узнает» Т-лимфоцит
В зависимости от того какую молекулу АГ+МНС1 или МНС2 «узнает» Т-лимфоцит

Рецепторы Т-лимфоцита
Рецепторы Т-лимфоцита
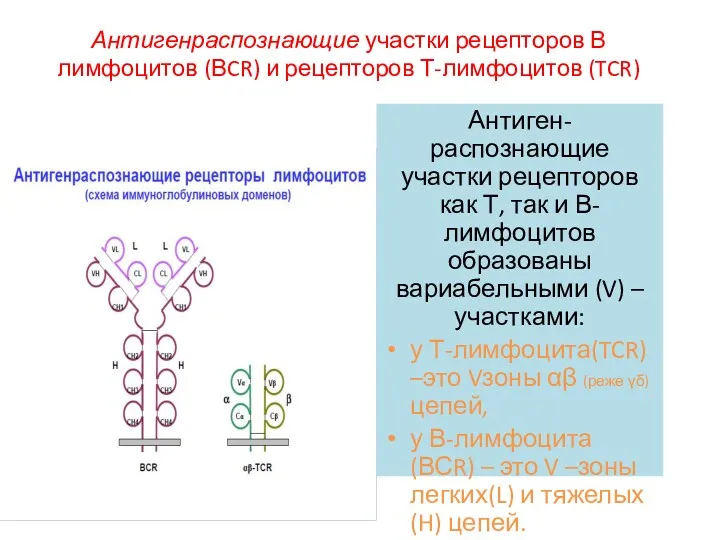
Антигенраспознающие участки рецепторов В лимфоцитов (ВCR) и рецепторов Т-лимфоцитов (TCR)
Антиген-распознающие участки
Антигенраспознающие участки рецепторов В лимфоцитов (ВCR) и рецепторов Т-лимфоцитов (TCR)
Антиген-распознающие участки
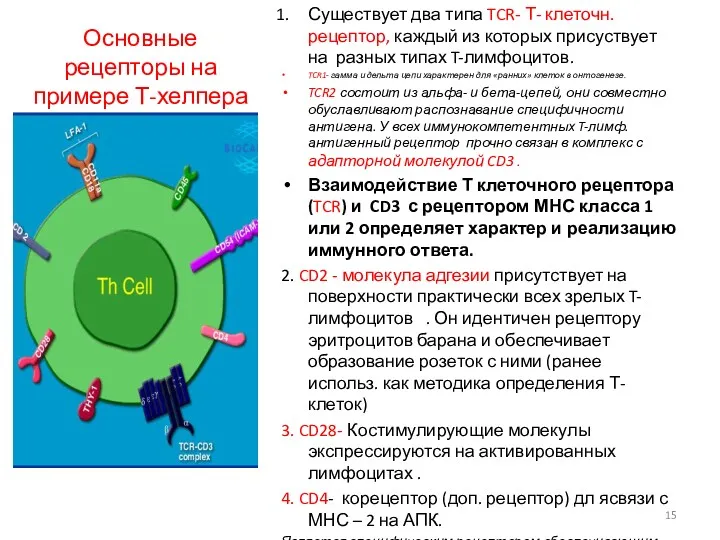
Основные рецепторы на примере Т-хелпера
Существует два типа TCR- Т- клеточн.рецептор, каждый
Основные рецепторы на примере Т-хелпера
Существует два типа TCR- Т- клеточн.рецептор, каждый

Популяции лимфоцитов
Популяции лимфоцитов
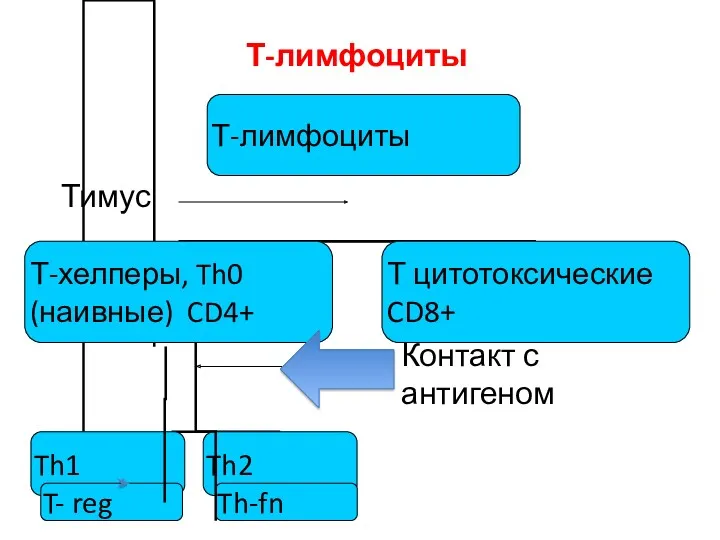
Т-лимфоциты
Тимус
Контакт с антигеном
Th-fn
T- reg
Т-лимфоциты
Тимус
Контакт с антигеном
Th-fn
T- reg

T-цитотоксические
Т-киллеры
(CTL или CD8+)
T-цитотоксические
Т-киллеры
(CTL или CD8+)

T-цитотоксические лимф. (кратко)
Поверхностный маркер – СD3 (сигнальный комплекс, связанный с
T-цитотоксические лимф. (кратко)
Поверхностный маркер – СD3 (сигнальный комплекс, связанный с

Т-киллер атакует клетку, на поверхности которой экспрессирован ассоциированный с MHC-I антиген,
Т-киллер атакует клетку, на поверхности которой экспрессирован ассоциированный с MHC-I антиген,

Т-хелперы и
их роль в иммунном ответе
Т-хелперы и
их роль в иммунном ответе

Th0-лимфоциты (наивные)
Поверхностный маркер – СD3, CD4 (рецептор к МНС II)
Th0-лимфоциты (наивные)
Поверхностный маркер – СD3, CD4 (рецептор к МНС II)

T-хелперы CD4+
В зависмости от характера и презентации антигена (кто презентировал
T-хелперы CD4+
В зависмости от характера и презентации антигена (кто презентировал
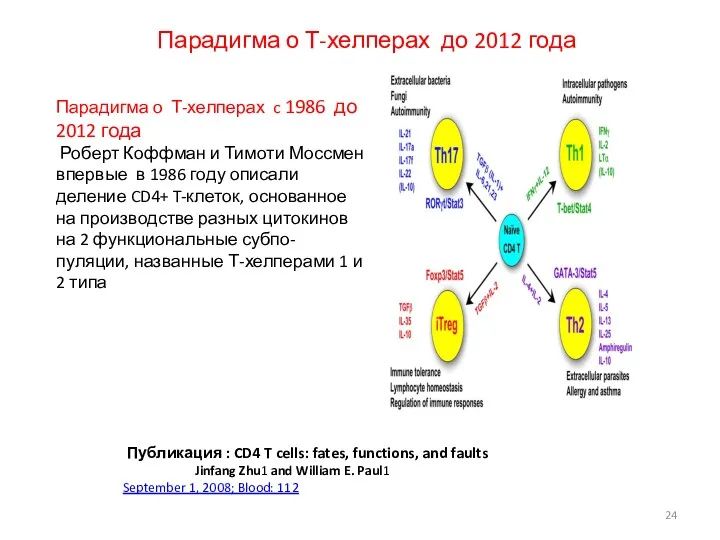
Парадигма о Т-хелперах до 2012 года
Публикация : CD4 T cells:
Парадигма о Т-хелперах до 2012 года
Публикация : CD4 T cells:
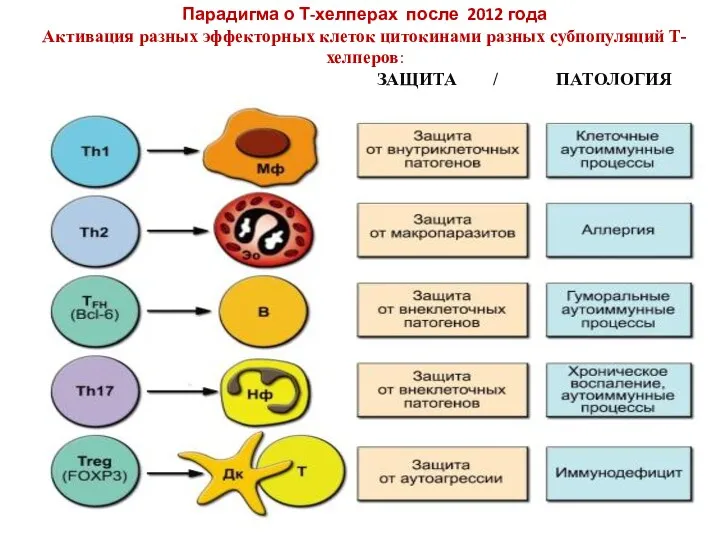
Парадигма о Т-хелперах после 2012 года
Активация разных эффекторных клеток цитокинами разных
Парадигма о Т-хелперах после 2012 года Активация разных эффекторных клеток цитокинами разных

Т-лимфоциты-участники адаптивного (антигенспецифического) ответа
Т-лимфоциты-участники адаптивного (антигенспецифического) ответа

Дополнительный матерал- вне рамок курса
Дополнительный матерал- вне рамок курса

Другие субпопуляции Т-хелперов
(материал не входит в рамки программы курса- дополнительно)
Th17
Если
Другие субпопуляции Т-хелперов
(материал не входит в рамки программы курса- дополнительно)
Th17
Если
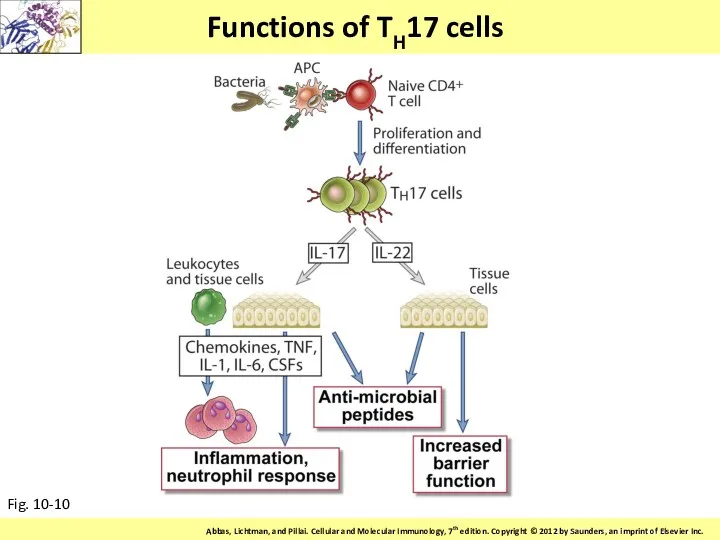
Functions of TH17 cells
Fig. 10-10
Functions of TH17 cells
Fig. 10-10

Субпопуляции Т-хелперов: регуляторные клетки - T reg (CD4+CD25+FOXP3 клетки)
«Т- реги» или
Субпопуляции Т-хелперов: регуляторные клетки - T reg (CD4+CD25+FOXP3 клетки) «Т- реги» или
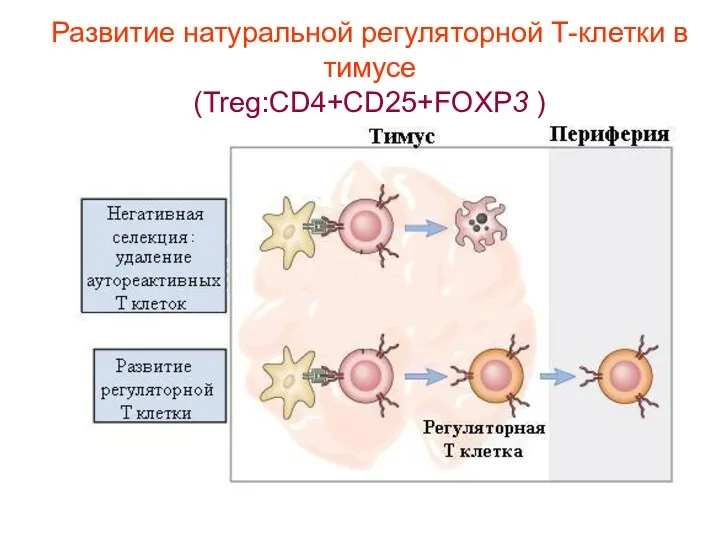
Развитие натуральной регуляторной Т-клетки в тимусе
(Treg:CD4+CD25+FOXP3 )
Развитие натуральной регуляторной Т-клетки в тимусе
(Treg:CD4+CD25+FOXP3 )

Окончание дополнительного материала
Окончание дополнительного материала

Иммунологическая память. Субопуляции клеток памяти
Иммунологическая память. Субопуляции клеток памяти

Иммунологическая память. Субопуляции клеток памяти
Иммунологическая память –
феномен более раннего и
Иммунологическая память. Субопуляции клеток памяти
Иммунологическая память –
феномен более раннего и
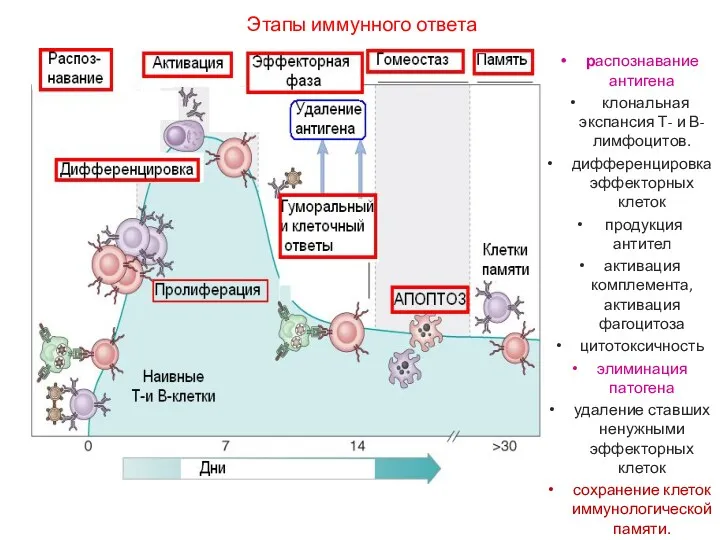
Этапы иммунного ответа
распознавание антигена
клональная экспансия Т- и В-лимфоцитов.
дифференцировка эффекторных
Этапы иммунного ответа
распознавание антигена
клональная экспансия Т- и В-лимфоцитов.
дифференцировка эффекторных

Основные субпопуляции Т-клеток памяти в лимфоидной ткани
локализуются Т клетки центральной
Основные субпопуляции Т-клеток памяти в лимфоидной ткани локализуются Т клетки центральной

Основные субпопуляции Т-клеток памяти:
Т-клетки эффекторной памяти (ТЕМ)
Т клетки эффекторной памяти
Основные субпопуляции Т-клеток памяти:
Т-клетки эффекторной памяти (ТЕМ)
Т клетки эффекторной памяти
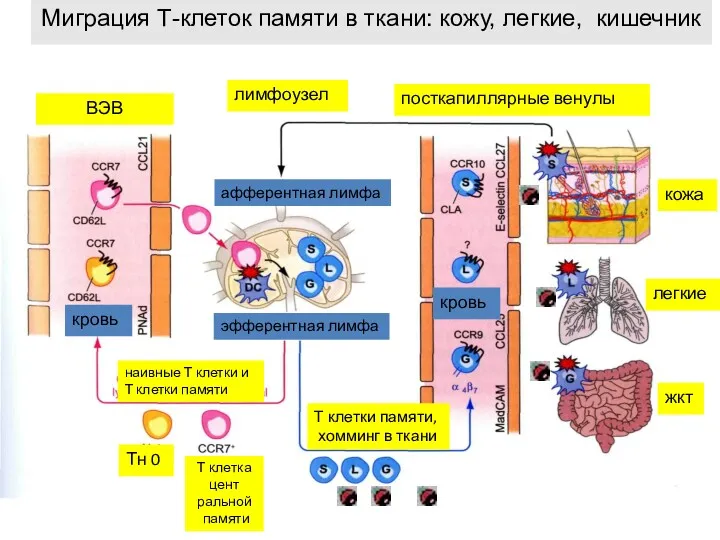
Миграция Т-клеток памяти в ткани: кожу, легкие, кишечник
лимфоузел
посткапиллярные венулы
кожа
легкие
жкт
Тн 0
Т клетка
Миграция Т-клеток памяти в ткани: кожу, легкие, кишечник
лимфоузел
посткапиллярные венулы
кожа
легкие
жкт
Тн 0
Т клетка
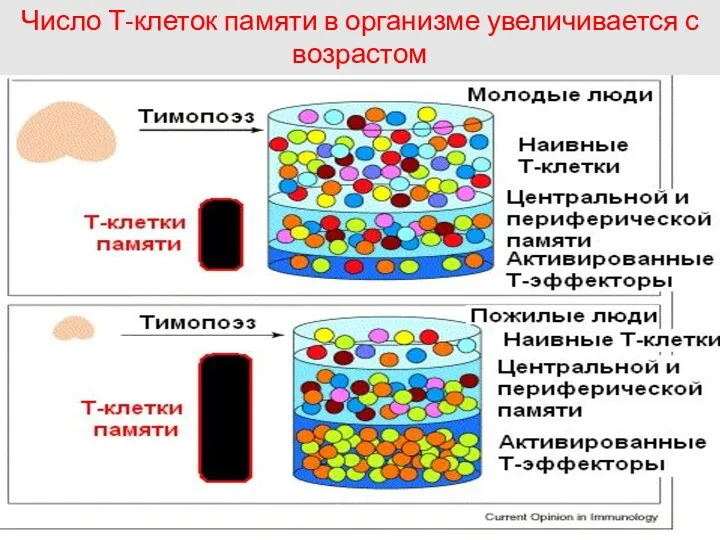
Число Т-клеток памяти в организме увеличивается с возрастом
Число Т-клеток памяти в организме увеличивается с возрастом

Что способствует формированию долговременной памяти на патоген?
Свойства самого антигена (иммуногенность), а
Что способствует формированию долговременной памяти на патоген?
Свойства самого антигена (иммуногенность), а
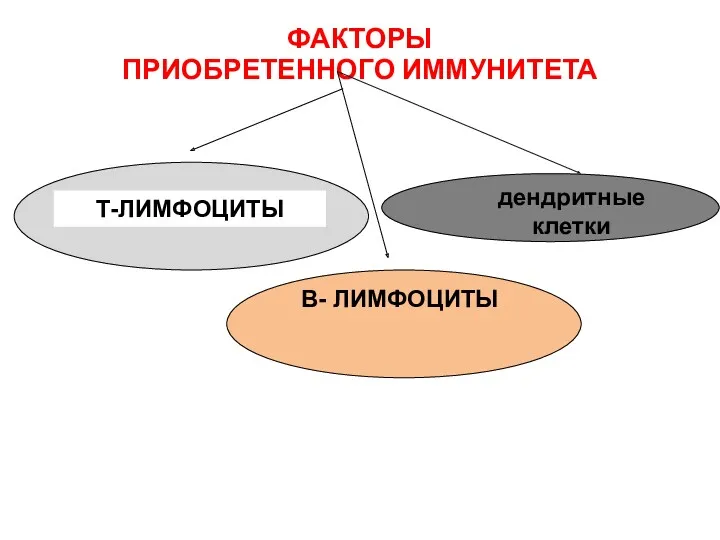
ФАКТОРЫ
ПРИОБРЕТЕННОГО ИММУНИТЕТА
Т-ЛИМФОЦИТЫ
В- ЛИМФОЦИТЫ
дендритные клетки
ФАКТОРЫ
ПРИОБРЕТЕННОГО ИММУНИТЕТА
Т-ЛИМФОЦИТЫ
В- ЛИМФОЦИТЫ
дендритные клетки
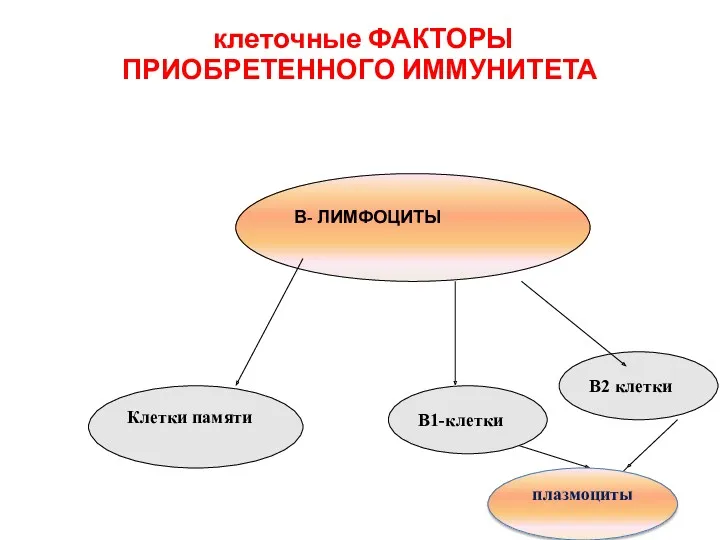
клеточные ФАКТОРЫ
ПРИОБРЕТЕННОГО ИММУНИТЕТА
В- ЛИМФОЦИТЫ
В1-клетки
В2 клетки
плазмоциты
Клетки памяти
клеточные ФАКТОРЫ
ПРИОБРЕТЕННОГО ИММУНИТЕТА
В- ЛИМФОЦИТЫ
В1-клетки
В2 клетки
плазмоциты
Клетки памяти
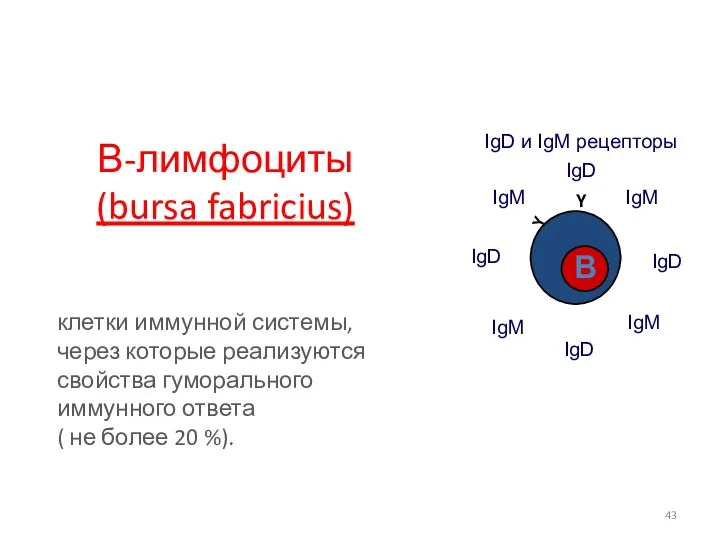
В-лимфоциты
(bursa fabricius)
Y
Y
клетки иммунной системы, через которые реализуются свойства
В-лимфоциты
(bursa fabricius)
Y
Y
клетки иммунной системы, через которые реализуются свойства

Характеристика В-клеток:
функции:
способность дифференцироваться в плазматические клетки и продуцировать антитела;
выступать
Характеристика В-клеток:
функции:
способность дифференцироваться в плазматические клетки и продуцировать антитела;
выступать

Типы В-клеток
В1-лимфоциты отвечают на Т независимые антигены (нативный или растворимый антиген),
Типы В-клеток
В1-лимфоциты отвечают на Т независимые антигены (нативный или растворимый антиген),

Рецепторы B-лимфоцитов
узнающим антиген, является IgM/D рецептор.
Это мономерный иммуноглобулин. Число молекул
Рецепторы B-лимфоцитов
узнающим антиген, является IgM/D рецептор.
Это мономерный иммуноглобулин. Число молекул

Рецепторы В-лимфоцитов
BCR
CD19 - самый ранний маркер В-клеток, .
CD20 - маркер
Рецепторы В-лимфоцитов
BCR
CD19 - самый ранний маркер В-клеток, .
CD20 - маркер

B-клеточный распознающий рецептор (BCR)
Состоит из мембранной формы Ig D или
B-клеточный распознающий рецептор (BCR)
Состоит из мембранной формы Ig D или

Онтогенез В-лимфоцитов
Предшественники В-лимфоцитов обнаружены в островках гемопоэтической ткани эмбриональной печени
Онтогенез В-лимфоцитов
Предшественники В-лимфоцитов обнаружены в островках гемопоэтической ткани эмбриональной печени

Плазмоцит
Плазмоцит

Плазматические клетки (“МАМы антител”)
нет в кровотоке
локализация –
Плазматические клетки (“МАМы антител”)
нет в кровотоке
локализация –
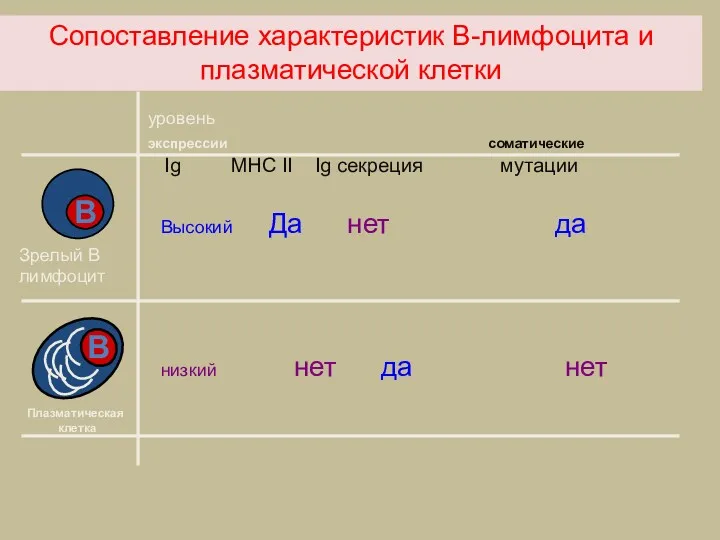
Сопоставление характеристик В-лимфоцита и плазматической клетки
уровень
экспрессии соматические
Ig MHC II
Сопоставление характеристик В-лимфоцита и плазматической клетки
уровень
экспрессии соматические
Ig MHC II

Центр размножения ЛУ(зародышевые центры)
Особая область в лимфатических узлах или селезенке, в
Центр размножения ЛУ(зародышевые центры)
Особая область в лимфатических узлах или селезенке, в
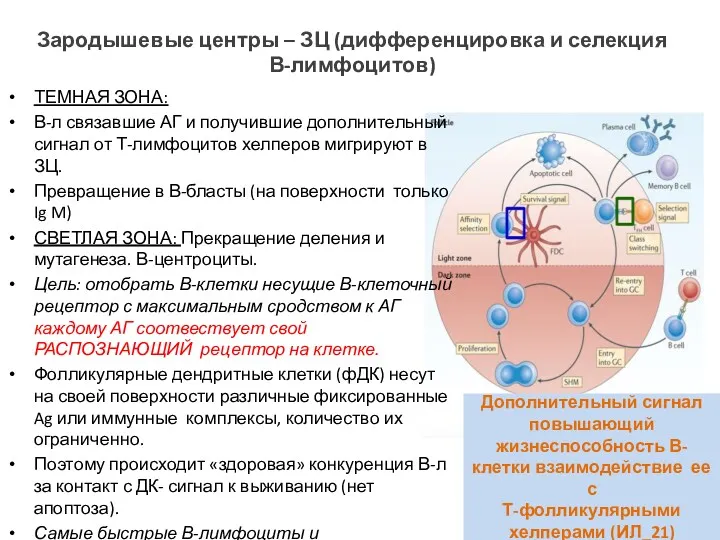
Зародышевые центры – ЗЦ (дифференцировка и селекция В-лимфоцитов)
ТЕМНАЯ ЗОНА:
В-л связавшие АГ
Зародышевые центры – ЗЦ (дифференцировка и селекция В-лимфоцитов)
ТЕМНАЯ ЗОНА:
В-л связавшие АГ
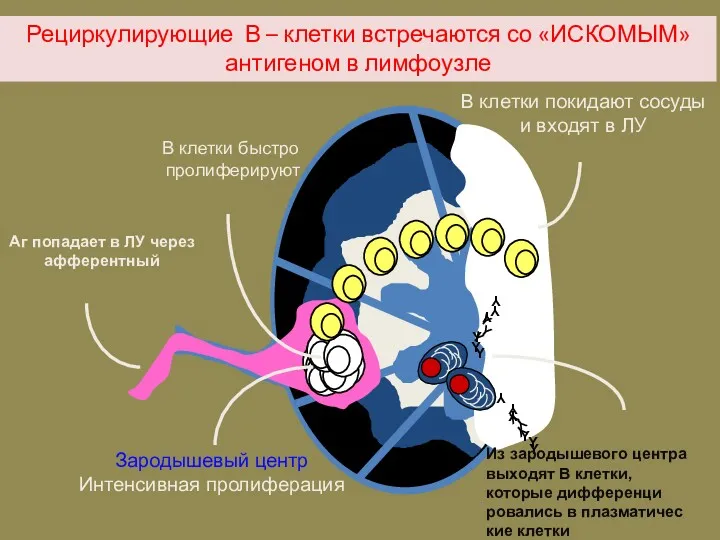
Рециркулирующие В – клетки встречаются со «ИСКОМЫМ» антигеном в лимфоузле
Рециркулирующие В – клетки встречаются со «ИСКОМЫМ» антигеном в лимфоузле
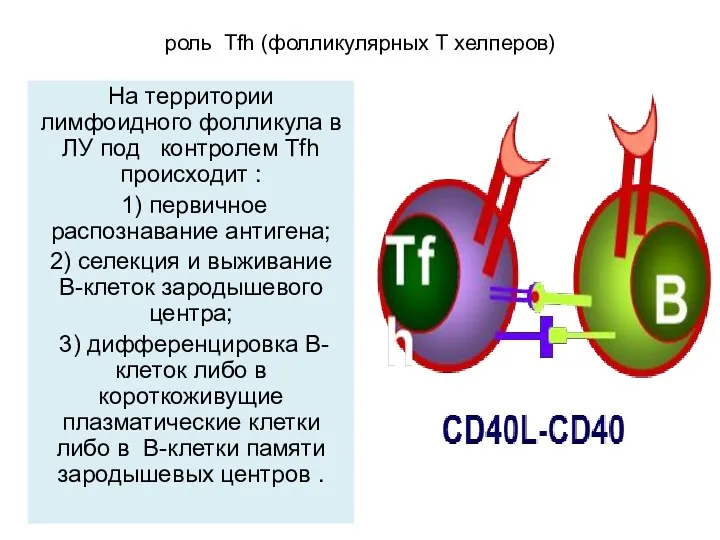
роль Tfh (фолликулярных Т хелперов)
На территории лимфоидного фолликула в
роль Tfh (фолликулярных Т хелперов)
На территории лимфоидного фолликула в
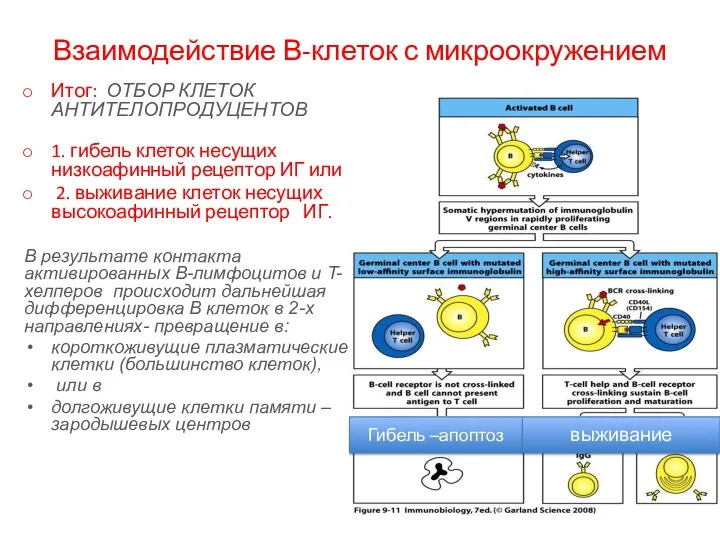
Взаимодействие В-клеток с микроокружением
Итог: ОТБОР КЛЕТОК АНТИТЕЛОПРОДУЦЕНТОВ
1. гибель клеток несущих низкоафинный
Взаимодействие В-клеток с микроокружением
Итог: ОТБОР КЛЕТОК АНТИТЕЛОПРОДУЦЕНТОВ
1. гибель клеток несущих низкоафинный

Первичный иммунный ответ
Плазматические клетки при первичном ответе синтезируют антитела с
Первичный иммунный ответ
Плазматические клетки при первичном ответе синтезируют антитела с
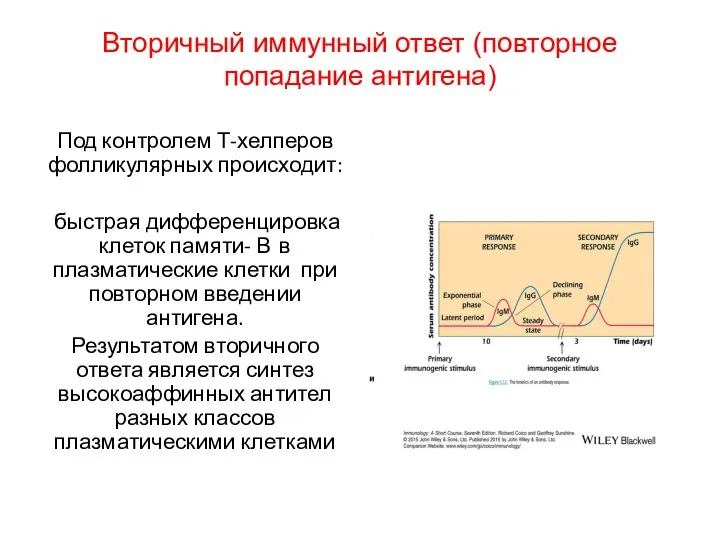
Вторичный иммунный ответ (повторное попадание антигена)
Под контролем Т-хелперов фолликулярных происходит:
быстрая дифференцировка
Вторичный иммунный ответ (повторное попадание антигена)
Под контролем Т-хелперов фолликулярных происходит:
быстрая дифференцировка

Цитокины
(смотри лекцию №2)
Цитокины
(смотри лекцию №2)

Цитокины – небольшие белки
(8-80 кДа), действующие аутокринно или паракринно.
Виды:
Интерлейкины
Цитокины – небольшие белки
(8-80 кДа), действующие аутокринно или паракринно.
Виды:
Интерлейкины

Функции цитокинов
регуляция гемопоэза
регуляция иммунного ответа и воспалительных процессов
участие в ангиогенезе
апоптоз
Функции цитокинов
регуляция гемопоэза
регуляция иммунного ответа и воспалительных процессов
участие в ангиогенезе
апоптоз

Взаимодействие клеток иммунной системы в ходе иммунного ответа(общие закономерности)
КООПЕРАЦИЯ КЛЕТОК
Взаимодействие клеток иммунной системы в ходе иммунного ответа(общие закономерности)
КООПЕРАЦИЯ КЛЕТОК

Иммунный синапс – основа кооперации клеток
Синапс – область соприкосновения клеток, где
Иммунный синапс – основа кооперации клеток
Синапс – область соприкосновения клеток, где
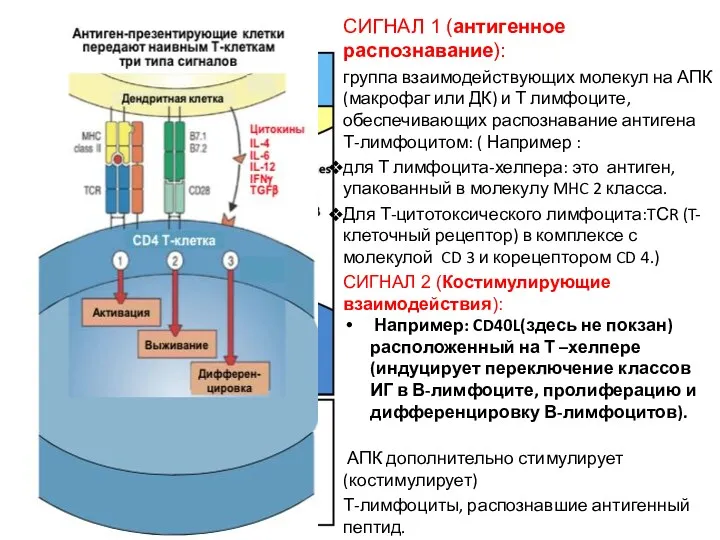
СИГНАЛ 1 (антигенное распознавание):
группа взаимодействующих молекул на АПК (макрофаг или ДК)
СИГНАЛ 1 (антигенное распознавание):
группа взаимодействующих молекул на АПК (макрофаг или ДК)
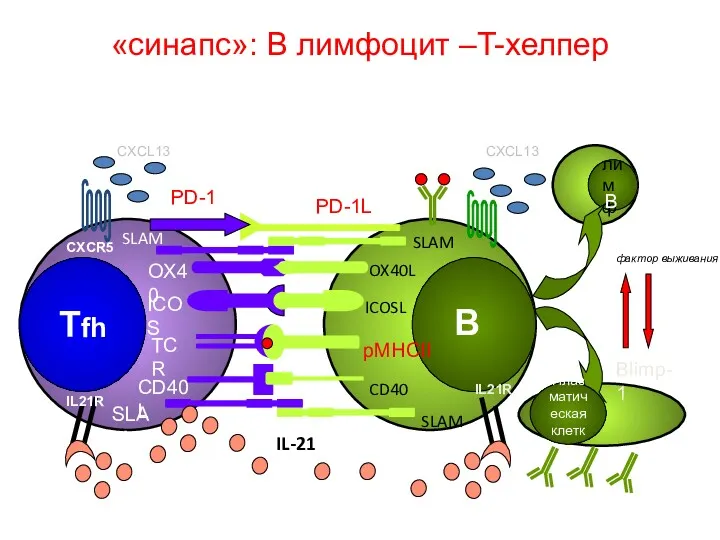
«синапс»: В лимфоцит –T-хелпер
IL21R
IL-21
IL21R
CXCR5
CXCL13
CXCL13
PD-1
PD-1L
«синапс»: В лимфоцит –T-хелпер
IL21R
IL-21
IL21R
CXCR5
CXCL13
CXCL13
PD-1
PD-1L

Отсутствие экспрессии костимулирующих молекул антгенпрезетрующими клетками приводит к тому, что иммунный
Отсутствие экспрессии костимулирующих молекул антгенпрезетрующими клетками приводит к тому, что иммунный
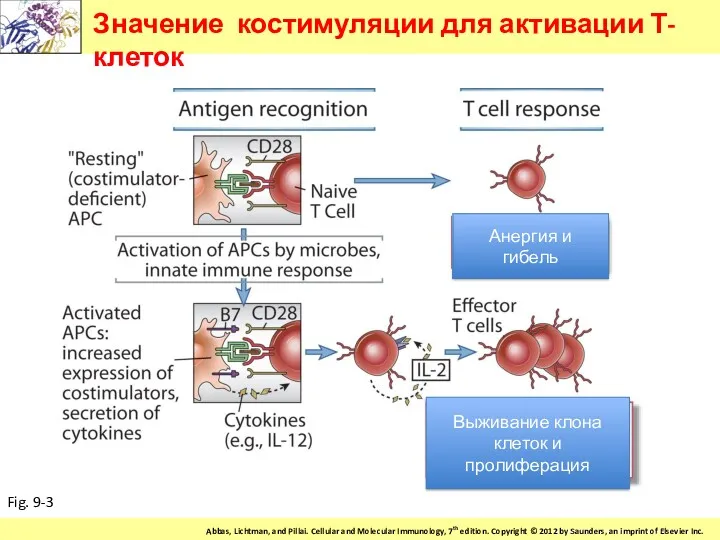
Значение костимуляции для активации Т-клеток
Fig. 9-3
Анергия и гибель
Выживание клона клеток
Значение костимуляции для активации Т-клеток
Fig. 9-3
Анергия и гибель
Выживание клона клеток
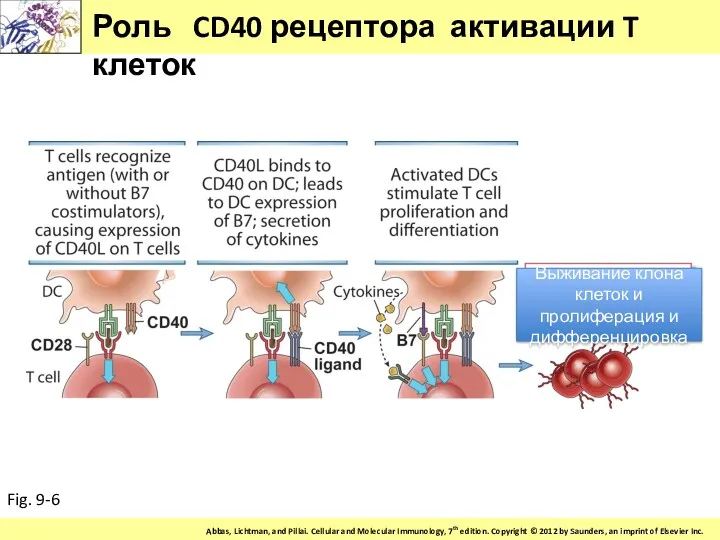
Роль CD40 рецептора активации T клеток
Fig. 9-6
Выживание клона клеток и
Роль CD40 рецептора активации T клеток
Fig. 9-6
Выживание клона клеток и

Это интересно…
Помимо стимулирующих воздействий в иммунной системе предусмотрены и подавляющие
Это интересно…
Помимо стимулирующих воздействий в иммунной системе предусмотрены и подавляющие

Клеточный тип иммунного ответа
КООПЕРАЦИЯ КЛЕТОК В ИММУННОМ ОТВЕТЕ
Клеточный тип иммунного ответа
КООПЕРАЦИЯ КЛЕТОК В ИММУННОМ ОТВЕТЕ

Клеточный тип иммунного ответа
( инициаторы: макрофаги и дендр. клетки)
Клеточный тип иммунного ответа
( инициаторы: макрофаги и дендр. клетки)

Клеточный тип иммунного ответа (ИО)
Основные цитокины, определяющие тип иммунного ответа
Клеточный тип иммунного ответа (ИО)
Основные цитокины, определяющие тип иммунного ответа
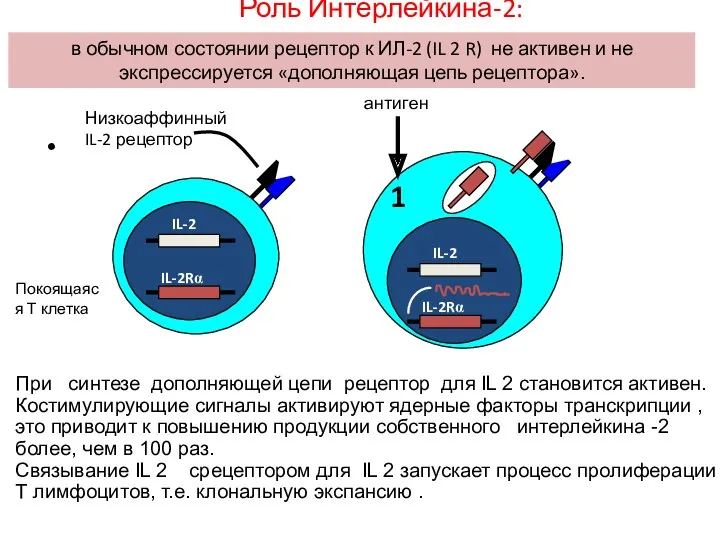
экспрессия
β и γ цепей IL-2 рецептора, но не α цепи
в
экспрессия
β и γ цепей IL-2 рецептора, но не α цепи
в
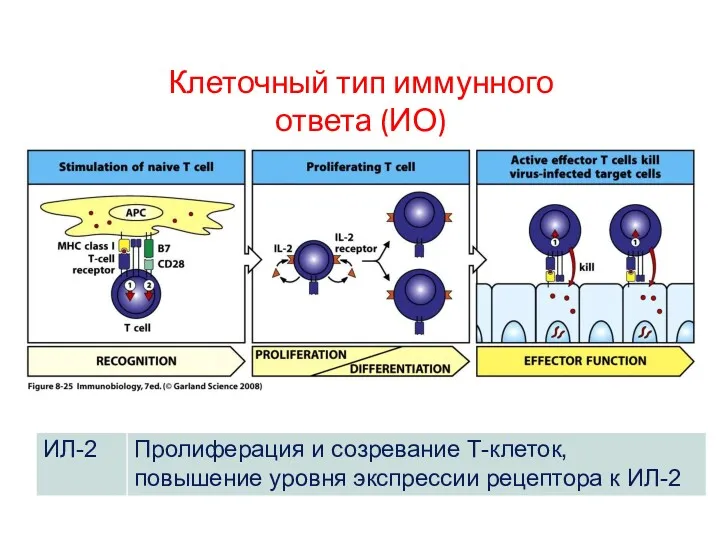
Клеточный тип иммунного ответа (ИО)
Клеточный тип иммунного ответа (ИО)

Этапы клеточного типа иммунного ответа (ИО)
Происходит взаимодействие: комплекса MHC I
Этапы клеточного типа иммунного ответа (ИО)
Происходит взаимодействие: комплекса MHC I
Общая схема клеточного им ответа
Общая схема клеточного им ответа
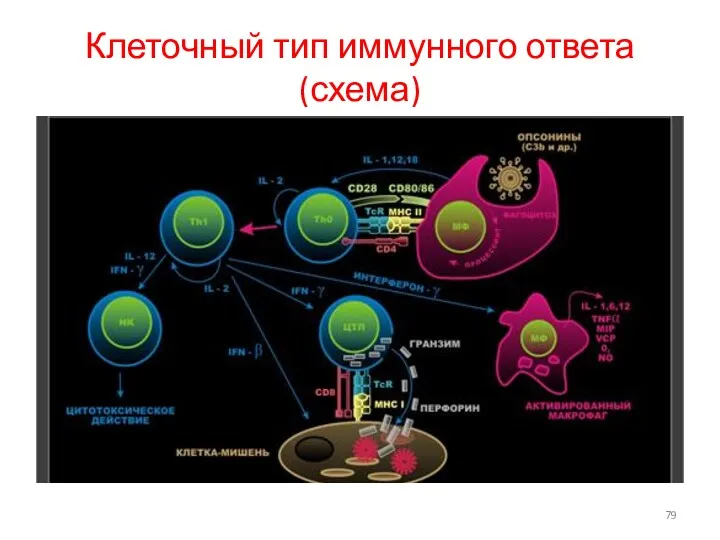
Клеточный тип иммунного ответа(схема)
Клеточный тип иммунного ответа(схема)

Гуморальный тип иммунного ответа
КООПЕРАЦИЯ КЛЕТОК В ИММУННОМ ОТВЕТЕ
Гуморальный тип иммунного ответа
КООПЕРАЦИЯ КЛЕТОК В ИММУННОМ ОТВЕТЕ

Гуморальный тип иммунного ответа ( инициаторы: В-лимфоциты и дендр. клетки)
Переход
Гуморальный тип иммунного ответа ( инициаторы: В-лимфоциты и дендр. клетки)
Переход

Основные «участники» синапсов в зависимости от типа и вида распознаваемого Аг
Основные «участники» синапсов в зависимости от типа и вида распознаваемого Аг

В зависимости от состава антигена выделяю ТИМУСзависимые АГ и Тимус не
В зависимости от состава антигена выделяю ТИМУСзависимые АГ и Тимус не

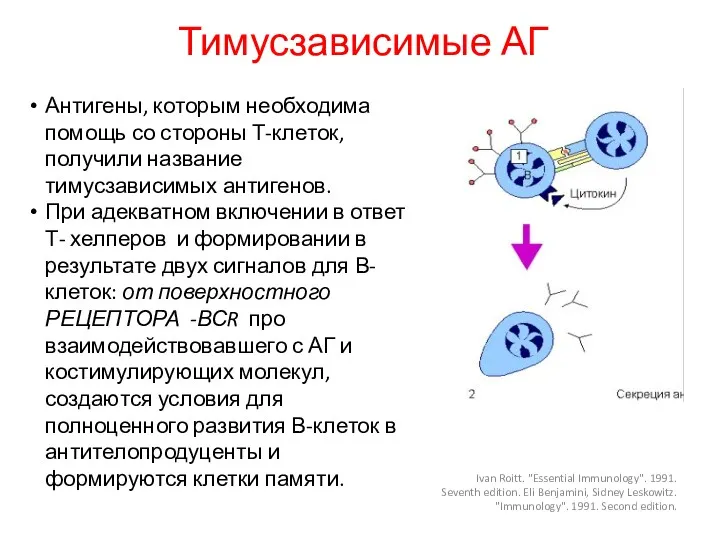
Тимусзависимые АГ
Нужна помощь хелперов для В-лимфоцитов для распознавания АГ
Тимусзависимые АГ
Нужна помощь хелперов для В-лимфоцитов для распознавания АГ
Тимусзависимые АГ
Антигены, которым необходима помощь со стороны Т-клеток, получили название тимусзависимых
Тимусзависимые АГ
Антигены, которым необходима помощь со стороны Т-клеток, получили название тимусзависимых
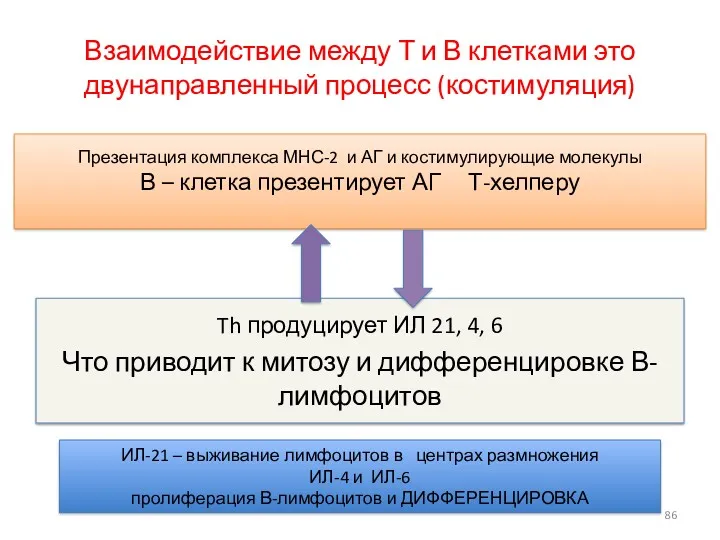
Взаимодействие между Т и В клетками это двунаправленный процесс (костимуляция)
Th продуцирует
Взаимодействие между Т и В клетками это двунаправленный процесс (костимуляция)
Th продуцирует
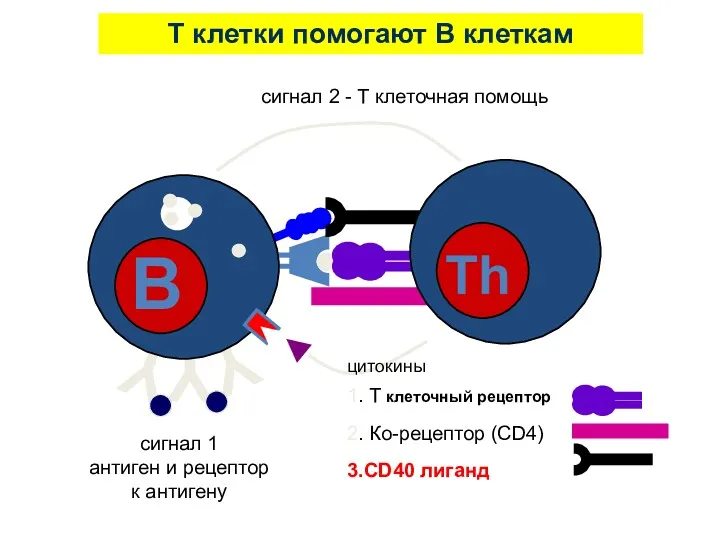
T клетки помогают B клеткам
сигнал 1
антиген и рецептор
к антигену
1. T
T клетки помогают B клеткам
сигнал 1
антиген и рецептор
к антигену
1. T

Тимусзависимые АГ
Зачем такие сложности?????
Если В-клетки взаимодействовали с антигеном (АГ), но
Тимусзависимые АГ
Зачем такие сложности?????
Если В-клетки взаимодействовали с антигеном (АГ), но

Тимусзависимые АГ
для распознавания АГ хелпер не нужен
В-лимфоцит справится САМ в
Тимусзависимые АГ
для распознавания АГ хелпер не нужен
В-лимфоцит справится САМ в
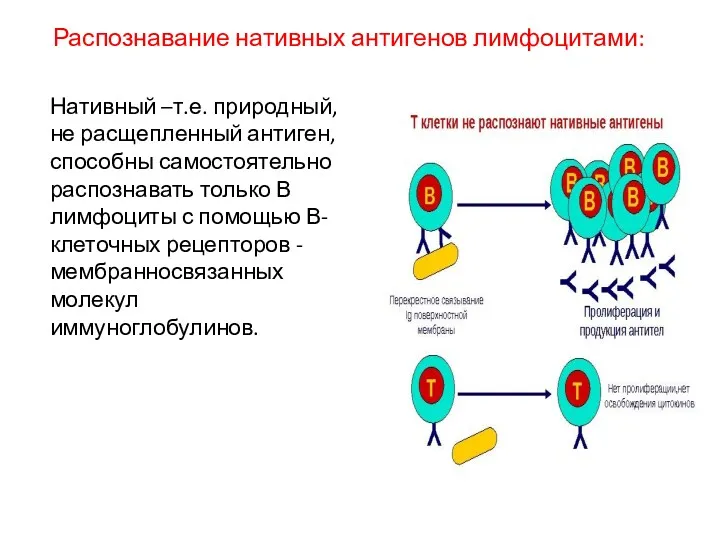
Распознавание нативных антигенов лимфоцитами:
Нативный –т.е. природный, не расщепленный антиген, способны
Распознавание нативных антигенов лимфоцитами:
Нативный –т.е. природный, не расщепленный антиген, способны
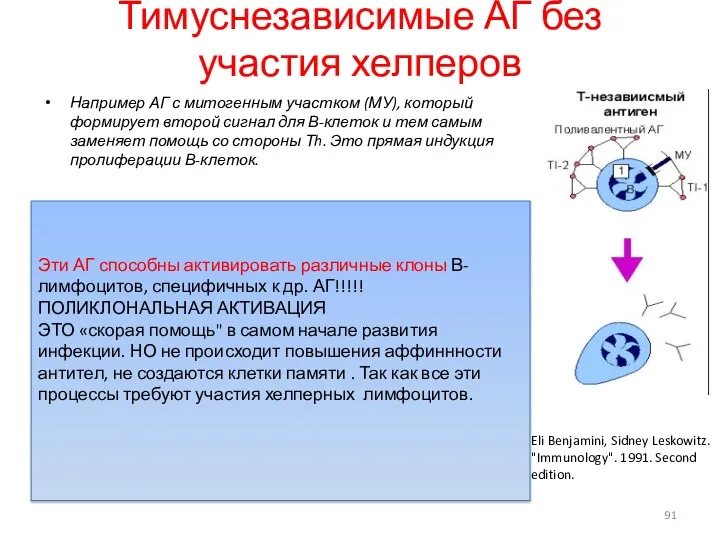
Тимуснезависимые АГ без участия хелперов
Например АГ с митогенным участком (МУ),
Тимуснезависимые АГ без участия хелперов
Например АГ с митогенным участком (МУ),

Первичный гуморальный ответ на тимуснезависимые АГ (Тi)немного слабее, чем на Т-
Первичный гуморальный ответ на тимуснезависимые АГ (Тi)немного слабее, чем на Т-
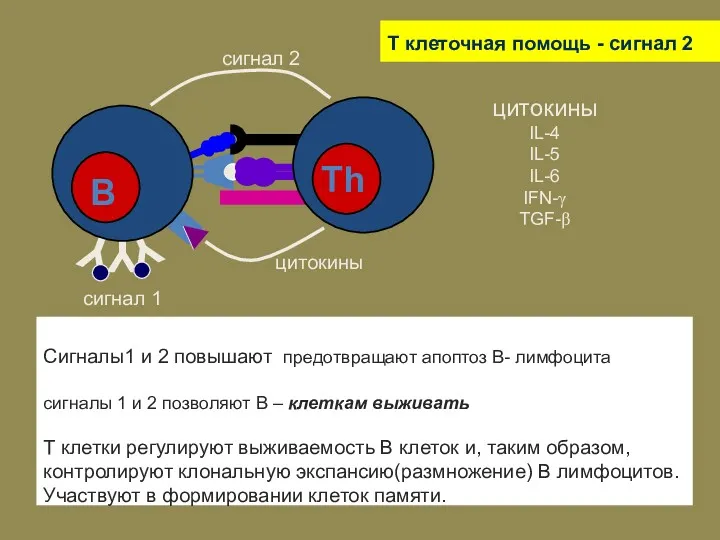
T клеточная помощь - сигнал 2
Сигналы1 и 2 повышают
T клеточная помощь - сигнал 2
Сигналы1 и 2 повышают
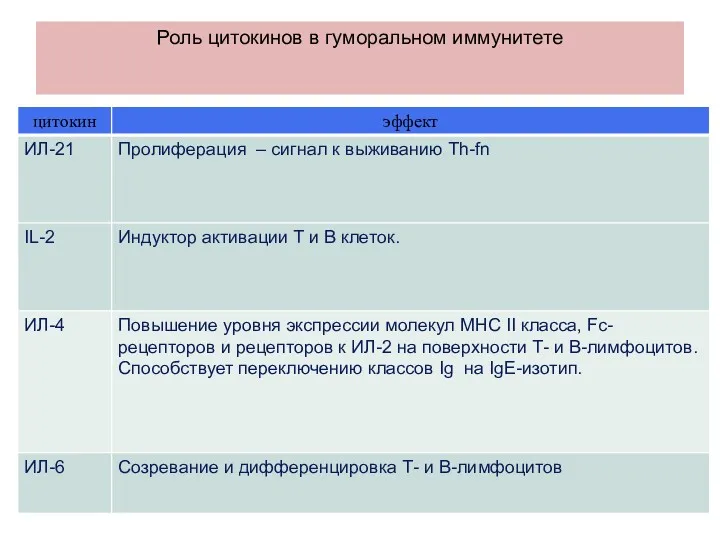
Роль цитокинов в гуморальном иммунитете
Роль цитокинов в гуморальном иммунитете

Этапы гуморального ИО
Наивные В- клетки сами распознают растворимый антиген (АГ) (без
Этапы гуморального ИО
Наивные В- клетки сами распознают растворимый антиген (АГ) (без
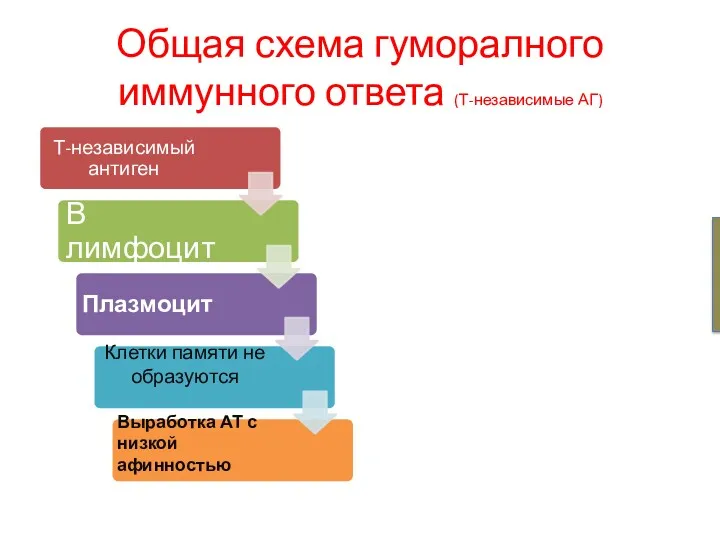
Общая схема гуморалного иммунного ответа (Т-независимые АГ)
Общая схема гуморалного иммунного ответа (Т-независимые АГ)

Общая схема гуморалного иммунного ответа (Т зависимые АГ)
Общая схема гуморалного иммунного ответа (Т зависимые АГ)
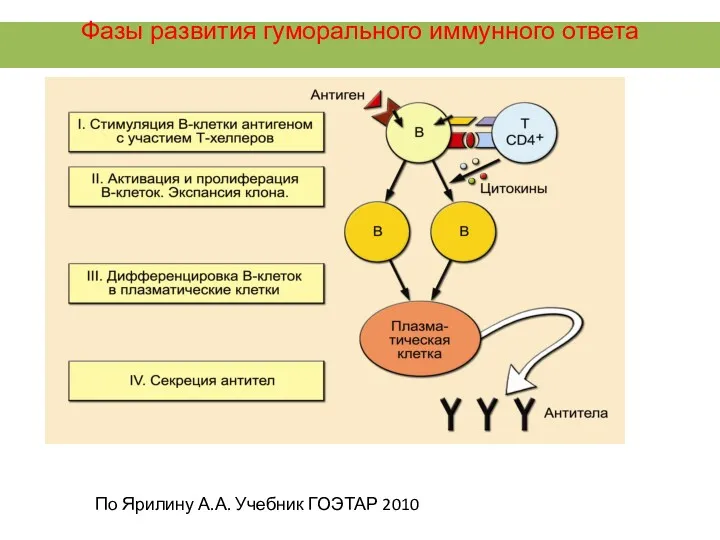
Фазы развития гуморального иммунного ответа
По Ярилину А.А. Учебник ГОЭТАР 2010
Фазы развития гуморального иммунного ответа
По Ярилину А.А. Учебник ГОЭТАР 2010
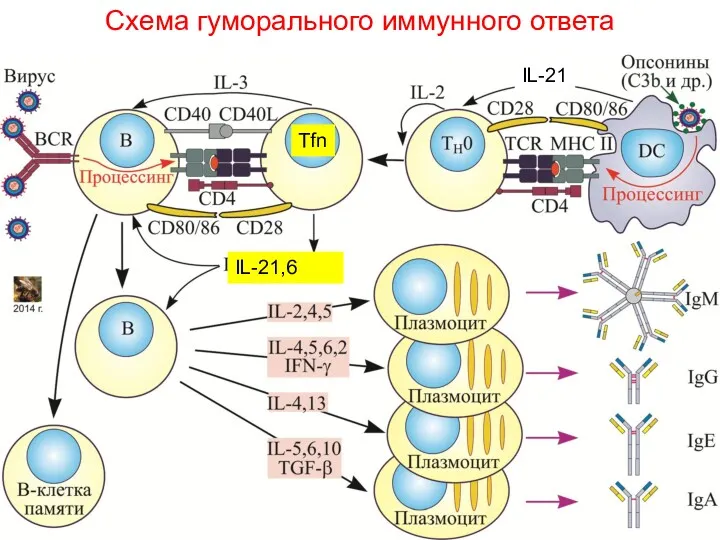
Схема гуморального иммунного ответа
Tfn
IL-21,6
IL-21
Схема гуморального иммунного ответа
Tfn
IL-21,6
IL-21

КООПЕРАЦИЯ КЛЕТОК В ИММУННОМ ОТВЕТЕ (общая схема)
КООПЕРАЦИЯ КЛЕТОК В ИММУННОМ ОТВЕТЕ (общая схема)
КООПЕРАЦИЯ КЛЕТОК ИММУННОЙ СИСТЕМЫ
1
2
3
4
5
6
7
8
КООПЕРАЦИЯ КЛЕТОК ИММУННОЙ СИСТЕМЫ
1
2
3
4
5
6
7
8
 Методики обследования памяти в дошкольный, школьный возраст, у взрослых людей
Методики обследования памяти в дошкольный, школьный возраст, у взрослых людей Сканери штрихових кодів
Сканери штрихових кодів презентация 75 лет Краснодарскому краю
презентация 75 лет Краснодарскому краю Простые слова
Простые слова Автомобиль будущего
Автомобиль будущего С Днём Рождения
С Днём Рождения Ивановский текстиль. Распродажа постельного белья
Ивановский текстиль. Распродажа постельного белья Morse Code Day
Morse Code Day Использование здоровье сберегающих технологий в логопедической практике
Использование здоровье сберегающих технологий в логопедической практике Презентация Воздушный океан 6 класс
Презентация Воздушный океан 6 класс Введение в гистологию. История науки
Введение в гистологию. История науки Архитектура с общей шиной
Архитектура с общей шиной Архитектор. Архитектура. Главные принципы архитектуры
Архитектор. Архитектура. Главные принципы архитектуры Статические тиристорные компенсаторы реактивной мощности (СТК)
Статические тиристорные компенсаторы реактивной мощности (СТК) Феодализм. Этапы феодальной экономики
Феодализм. Этапы феодальной экономики Производственный менеджмент
Производственный менеджмент prezentatsia
prezentatsia 70-летию со дня прорыва блокады Ленинграда посвязается....
70-летию со дня прорыва блокады Ленинграда посвязается.... Как общаться с наркоманом
Как общаться с наркоманом Система муниципальных правовых актов
Система муниципальных правовых актов Технические условия на переоборудование и перепланировку помещения
Технические условия на переоборудование и перепланировку помещения Современные энергосберегающие технологии
Современные энергосберегающие технологии Целиакия. Патофизиология целиакии
Целиакия. Патофизиология целиакии Подготовительная работа над научной статьей. Создание условий работы
Подготовительная работа над научной статьей. Создание условий работы Атомные электростанции, их виды
Атомные электростанции, их виды Подходы к иммунокорригирующей терапии у больных с респираторной аллергией
Подходы к иммунокорригирующей терапии у больных с респираторной аллергией Технолігїї підвищення продуктивності процесорів
Технолігїї підвищення продуктивності процесорів Разработка нефтяных месторождений. Часть 2
Разработка нефтяных месторождений. Часть 2