Слайд 2
Слайд 3

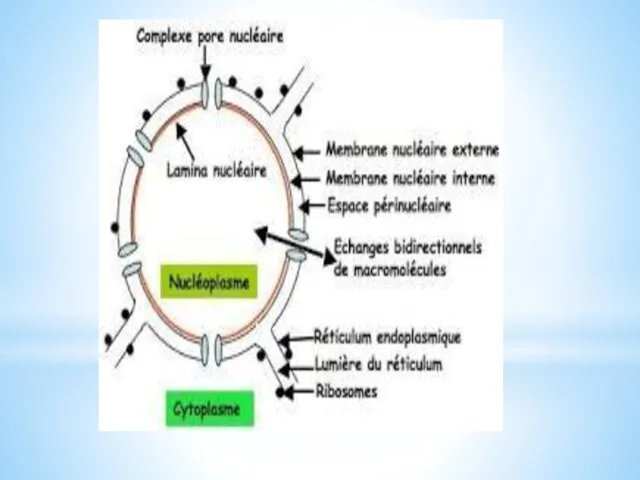
Localisation des lamines
(figure : vetopsy.fr d'après Dittmer)
Les filaments intermédiaires de
type V sont composés de protéines fibrillaires appelées lamines, retrouvées essentiellement dans le nucléoplasme. Elle forme :
la lamina nucléaire, armature protéique de 10 à 30 nm de large qui borde la membrane nucléaire sur sa face interne, formée par les différents types de lamines,
le voile nucléoplasmique diffus dans le nucléoplasme durant l'interphase, plutôt formé par les lamines B.
Les lamines sont aussi retrouvées dans le cytoplasme dans lequel elle sont synthétisées avant d'être exportées vers le noyau
Слайд 4
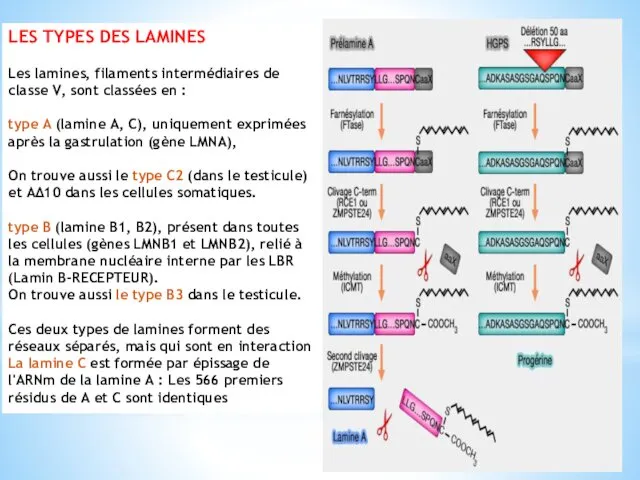
LES TYPES DES LAMINES
Les lamines, filaments intermédiaires de classe V, sont
classées en :
type A (lamine A, C), uniquement exprimées après la gastrulation (gène LMNA),
On trouve aussi le type C2 (dans le testicule) et AΔ10 dans les cellules somatiques.
type B (lamine B1, B2), présent dans toutes les cellules (gènes LMNB1 et LMNB2), relié à la membrane nucléaire interne par les LBR (Lamin B-RECEPTEUR).
On trouve aussi le type B3 dans le testicule.
Ces deux types de lamines forment des réseaux séparés, mais qui sont en interaction
La lamine C est formée par épissage de l'ARNm de la lamine A : Les 566 premiers résidus de A et C sont identiques
Слайд 5

Structure et polymérisation des lamines
Ces lamines diffèrent des protéines des
filaments intermédiaires cytoplasmiques par :
le domaine hélical en forme de tige (rod domain) plus long de 42 acides aminés, flanqué de part et d'autre d'un site de phosphorylation,
l'extrémité C-terminale pourvue :
d'un signal de localisation nucléaire (NLS : Nuclear Localization Signal), séquence d'acides aminés qui cible les protéines pour leur transport vers le noyau de la cellule,
d'un domaine IGF-like
Les lamines forment des homodimères in vitro même si elles peuvent s'hétérodiméres in vitro
Elles s'asocient par les deux extrémités de la tige - IFCM1 Elles se polymérisent tête à queue et peuvent former des associations de tétramères antiparallèles, dont on peut voir le domaine IGF-like globulaire se répéter tous les 48 nm.
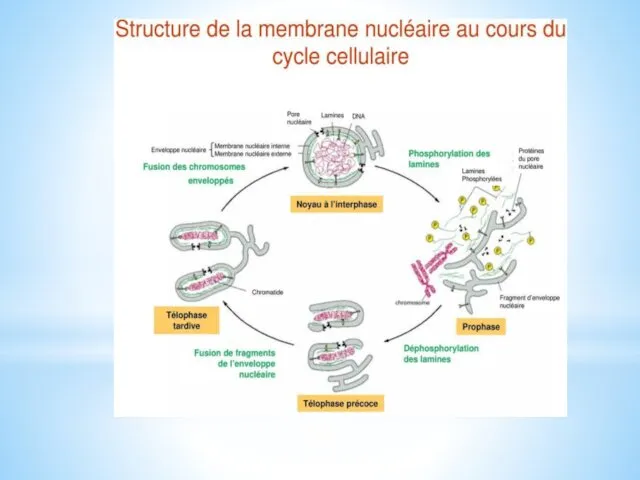
Les montages et démontages des filaments intermédiaires nucléaires sont beaucoup plus fréquents que les cytoplasmiques qui sont relativement stables, en particulier lors de la division cellulaire.
La dépolymérisation des lamines nucléaires, au début de la mitose, conduit à la désintégration de l'enveloppe nucléaire.
Слайд 6
Слайд 7
Слайд 8
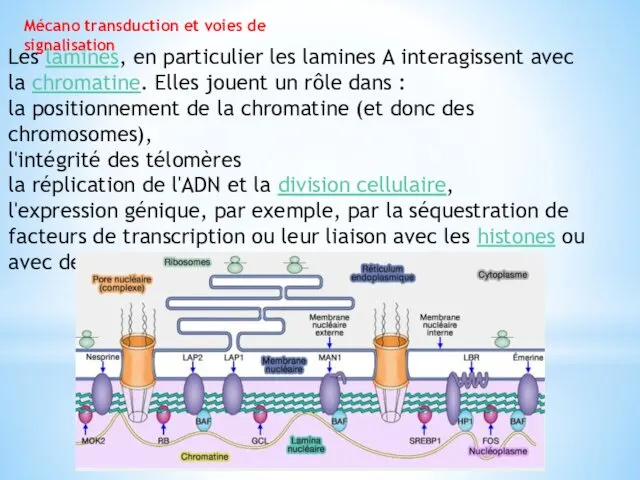
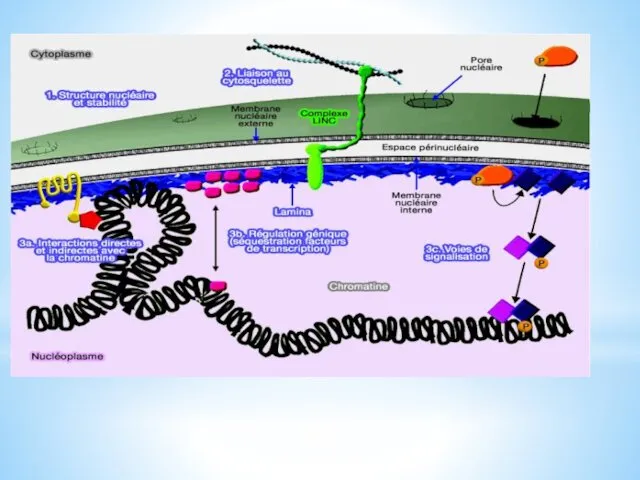
Mécano transduction et voies de signalisation
Les lamines, en particulier les lamines A
interagissent avec la chromatine. Elles jouent un rôle dans :
la positionnement de la chromatine (et donc des chromosomes),
l'intégrité des télomères
la réplication de l'ADN et la division cellulaire,
l'expression génique, par exemple, par la séquestration de facteurs de transcription ou leur liaison avec les histones ou avec des protéines qui se lient à la chromatine.
Слайд 9

l'émerine :
elle entre en contact avec de nombreuses protéines impliquées dans la
régulation de la transcription, des ARNm, et avec
les protéines BAF , également connues pour se lier directement au double brin de l'ADN, aux lamines et à plusieurs protéines membranaires nucléaires - LAP2, MAN1…
LBR (Lamin B Receptor) qui se lie aussi à de multiples cibles nucléaires, ADN, histones et diverses protéines associées à la chromatine

 Соединённые Штаты Америки (США)
Соединённые Штаты Америки (США) Коррекция общей моторики у детей со стертой формой дизартрии.
Коррекция общей моторики у детей со стертой формой дизартрии. Моє рідне місто Житомир
Моє рідне місто Житомир Классный час Ока Иванович Городовиков
Классный час Ока Иванович Городовиков ООО НПИ Биопрепараты. Почвоутомление – современная глобальная проблема
ООО НПИ Биопрепараты. Почвоутомление – современная глобальная проблема БИОЛОГИЯ палочники
БИОЛОГИЯ палочники Становление и развитие дошкольной педагогики
Становление и развитие дошкольной педагогики Rezultaty_priemnoy_komissii_2020
Rezultaty_priemnoy_komissii_2020 Социальная стратификация и социальная мобильность
Социальная стратификация и социальная мобильность Туризм как вид спорта
Туризм как вид спорта Химическая теория запаха
Химическая теория запаха Английский язык www.vk.com/egppt
Английский язык www.vk.com/egppt Опасные природные явления в литосфере
Опасные природные явления в литосфере Взаимодействие семьи с образовательными институтами
Взаимодействие семьи с образовательными институтами Велимир Хлебников
Велимир Хлебников От норм ГТО - к олимпийским медалям
От норм ГТО - к олимпийским медалям Внеурочное занятие. Праздничные блюда.
Внеурочное занятие. Праздничные блюда. Современные проблемы готовности детей 6-7 лет к школьному обучению.
Современные проблемы готовности детей 6-7 лет к школьному обучению. презентация Фронтовая тетрадь- песенник Трифонова С.И.
презентация Фронтовая тетрадь- песенник Трифонова С.И. Акустические свойства горных пород
Акустические свойства горных пород Маркетинговые стратегии территорий. (Лекция 3)
Маркетинговые стратегии территорий. (Лекция 3) Тағамды сақтау жолы консервілеу
Тағамды сақтау жолы консервілеу Игры на развитие мышления
Игры на развитие мышления 20231214_isakova_t.a._master-klass_izmerenie_pulsa_s_pomoshchyu_fotopletizmografii_v_ramkah_uchebnoy_laboratorii_po_neyrotehnologii_i_ispolzovanie_tsifrovogo_mikroskopa_na_urokah_biologii_0
20231214_isakova_t.a._master-klass_izmerenie_pulsa_s_pomoshchyu_fotopletizmografii_v_ramkah_uchebnoy_laboratorii_po_neyrotehnologii_i_ispolzovanie_tsifrovogo_mikroskopa_na_urokah_biologii_0 Проект Что в имени тебе моём
Проект Что в имени тебе моём Egozhev
Egozhev Разработка технической стратегии предприятия в области электроэнергетики
Разработка технической стратегии предприятия в области электроэнергетики примеры рекламы по потребителям
примеры рекламы по потребителям