- Липиды и везикулярный транспорт (лекция 6)

Содержание
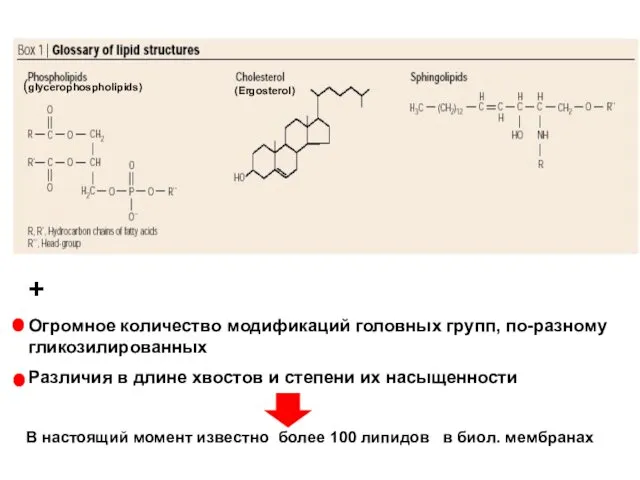
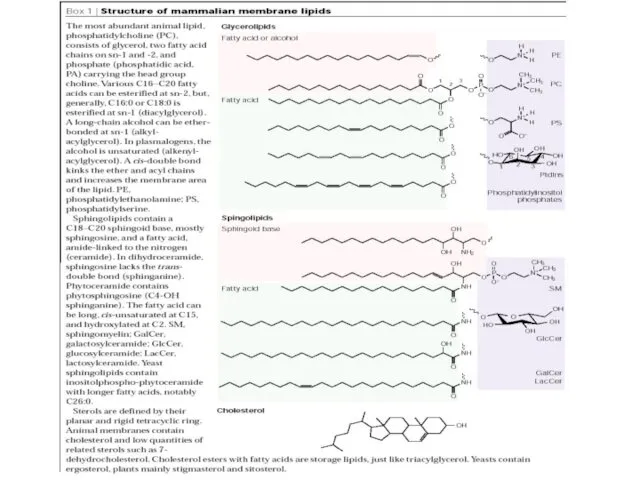
- 2. (glycerophospholipids) (Ergosterol) + Огромное количество модификаций головных групп, по-разному гликозилированных Различия в длине хвостов и степени
- 4. Исходно считалось, что липиды мембран играют роль инертной платформы, пассивного двумерного растворителя, в котором заякорены функционально
- 5. С развитием методических подходов, позволивших анализировать липидный состав отдельных органелл и маленьких участков мембран (флуоресцентно-меченые производные
- 6. ER: 60% PC 10% PE 10% PI Липидный состав мембран разных компартментов неоднороден
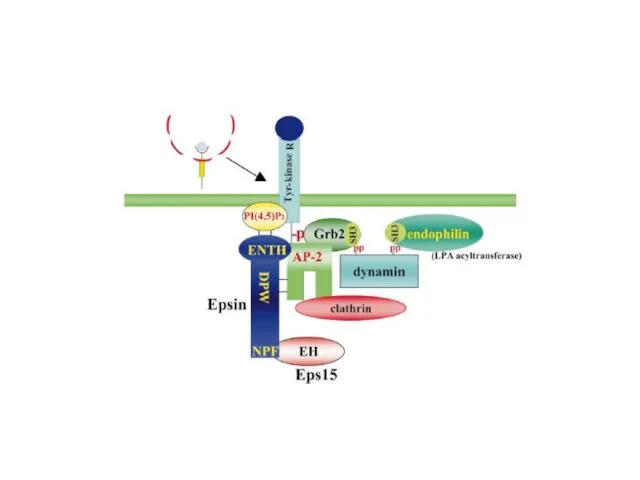
- 7. Липидные «территории» на эндоцитозном пути Различные фосфатидилинозитиды, продукты фосфатидилинозитолкиназ, работают на эндоцитозном пути LBPA (lysobiphosphatic acid)
- 8. Синтез липидов в ЭПР происходит в цитоплазматическом слое, а распределение между внутренним и внешним - одинаковое
- 9. Механизмы создания «трансмембранной» ассиметрии «мембранный» «цитоплазматический»
- 10. Латеральная ассиметрия и процесс сортировки липидов связаны между собой
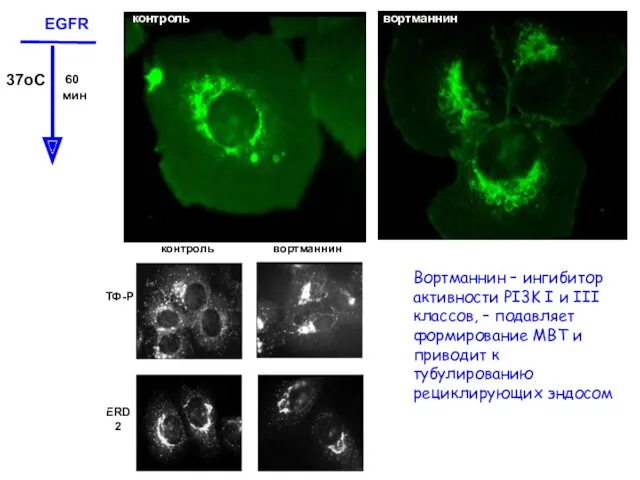
- 11. Нечувствительность к одному детергенту не означает устойчивость к детергентам вообще EGFR локализуется на ПМ в рафтах,
- 12. Рафты способны включать одни липиды и исключать другие, а также специфически ассоциироваться с определенными белками GPI-связанные
- 13. Липидная композиция рафта (минорные липиды) могут определять набор белков, включаемых в данный рафт и т.образом, способствовать
- 14. Сегрегация липидов может стимулироваться формообразованием Форма может определять сортировку липидов:
- 15. Липидный состав органелл в значительной степени определяется формой органеллы Структура конкретной липидной молекулы диктует ее предпочтительную
- 16. Сама форма липидных молекул (конус, цилиндр, инвертированный цилиндр) может определять кривизну мембраны, а перераспределение липидов вызывать
- 17. Активность липид-модифицирующих ферментов также может приводить к изменению формы липида, и следовательно, кривизны мембраны, как это
- 18. Как правило, искривление мембран происходит именно в области рафтов, что вовлекает их в процессы как слияния
- 19. В конечном итоге, форму мембраны или ее изменение определяют совместно как липиды, так и белки Механизм
- 20. Многочисленные липидозы (заболевания, вызываемые накоплением липидов в клетках) также связаны с нарушением их нормальной транспортировки При
- 21. Фосфатидилинозитиды (PtdIns) играют важную регуляторную роль в сигнальных и транспортных процессах
- 22. Может быть фосфорилирован по D3, 4 и 5-положениям инозитольного кольца PtIns
- 23. Пути взаимопревращений фосфорилированных форм PtdIns с участием фосфатидилинозитол-3-киназ
- 25. Ассиметричная локализация ключевых фосфоинозитидов в клеточных мембранах PI(4,5)P2 Синаптоджанин синаптотагмин PI(3,4)P2, PI(3,4,5)P3 PI3K (p85, ?) cинтез
- 27. Классификация PI3Ks Class Ia р110 –каталитическая субъединица р85α,β; р55α,γ; p50α-регуляторная субъединица Class Ib P110γ –каталититческая P101
- 28. PI3K I класса p85/p110
- 29. Сайты действия mVps34 1. Гомотипическое слияние ранних эндосом 2. Формирование внутренних пузырьков МВТ
- 31. ФИ-3-киназа р85 отвечает на действие ЭФР, но не ко-локализуется с рецептор-содержащими эндосомами. 0 мин 15 мин
- 33. Скачать презентацию
(glycerophospholipids)
(Ergosterol)
+
Огромное количество модификаций головных групп, по-разному гликозилированных
Различия в длине хвостов и
(glycerophospholipids)
(Ergosterol)
+
Огромное количество модификаций головных групп, по-разному гликозилированных
Различия в длине хвостов и
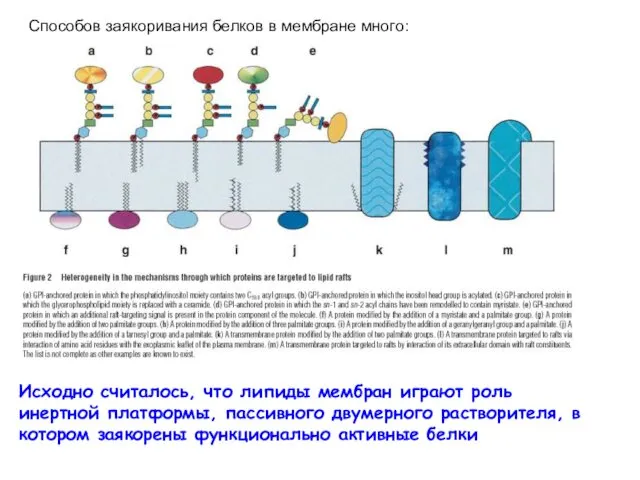
Исходно считалось, что липиды мембран играют роль инертной платформы, пассивного двумерного
Исходно считалось, что липиды мембран играют роль инертной платформы, пассивного двумерного

С развитием методических подходов, позволивших анализировать липидный состав отдельных органелл и
С развитием методических подходов, позволивших анализировать липидный состав отдельных органелл и
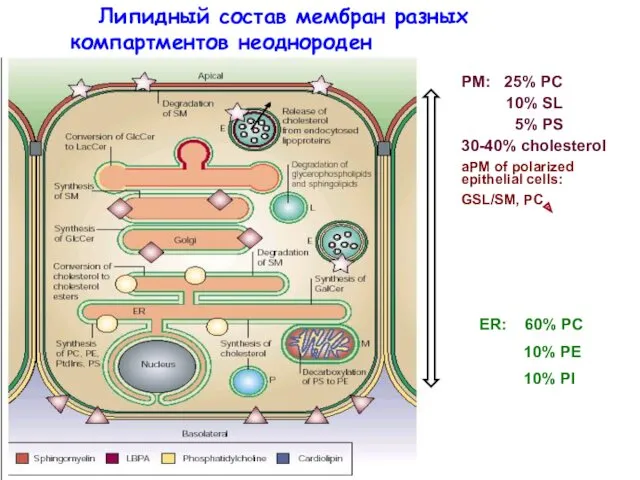
ER: 60% PC
10% PE
10% PI
Липидный состав мембран разных
ER: 60% PC
10% PE
10% PI
Липидный состав мембран разных
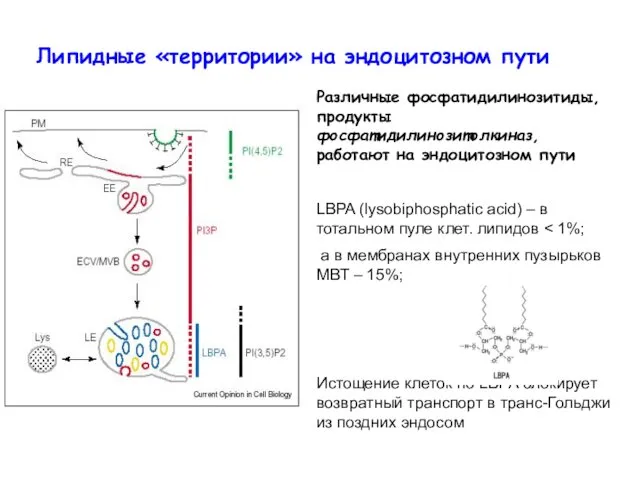
Липидные «территории» на эндоцитозном пути
Различные фосфатидилинозитиды, продукты фосфатидилинозитолкиназ, работают на эндоцитозном
Липидные «территории» на эндоцитозном пути
Различные фосфатидилинозитиды, продукты фосфатидилинозитолкиназ, работают на эндоцитозном
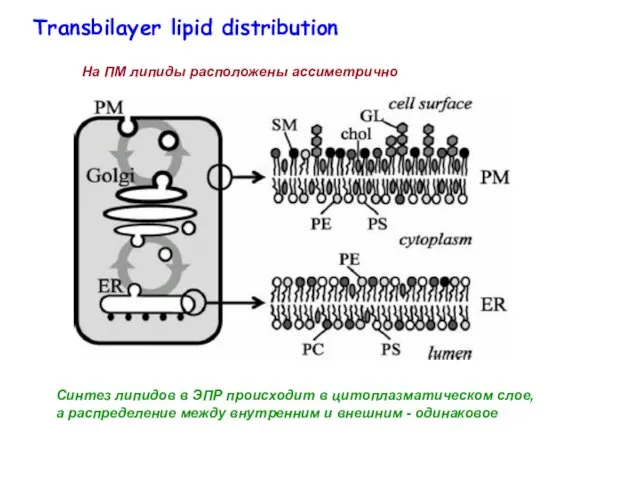
Синтез липидов в ЭПР происходит в цитоплазматическом слое, а распределение между
Синтез липидов в ЭПР происходит в цитоплазматическом слое, а распределение между
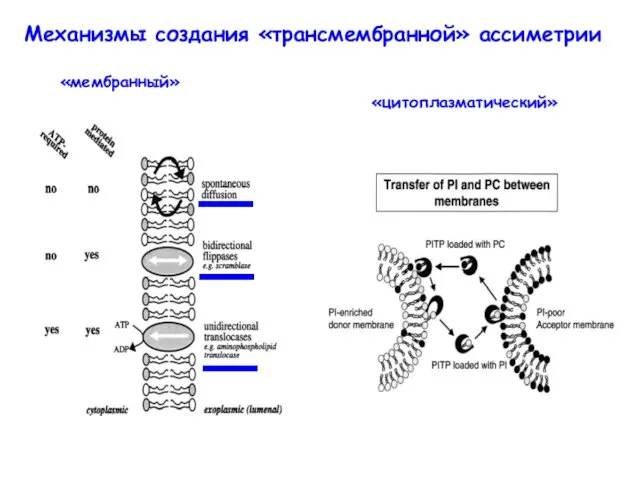
Механизмы создания «трансмембранной» ассиметрии
«мембранный»
«цитоплазматический»
Механизмы создания «трансмембранной» ассиметрии
«мембранный»
«цитоплазматический»

Латеральная ассиметрия и процесс сортировки липидов связаны между собой
Латеральная ассиметрия и процесс сортировки липидов связаны между собой
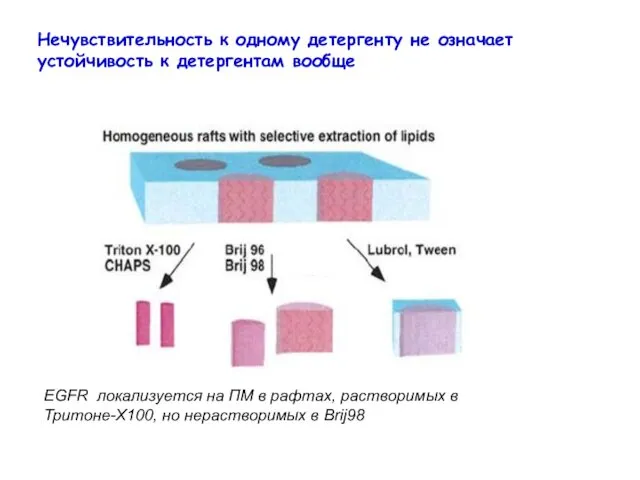
Нечувствительность к одному детергенту не означает устойчивость к детергентам вообще
EGFR локализуется
Нечувствительность к одному детергенту не означает устойчивость к детергентам вообще
EGFR локализуется
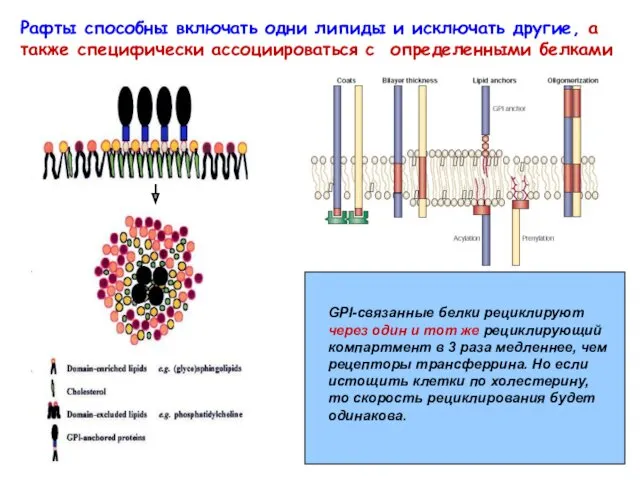
Рафты способны включать одни липиды и исключать другие, а также специфически
Рафты способны включать одни липиды и исключать другие, а также специфически
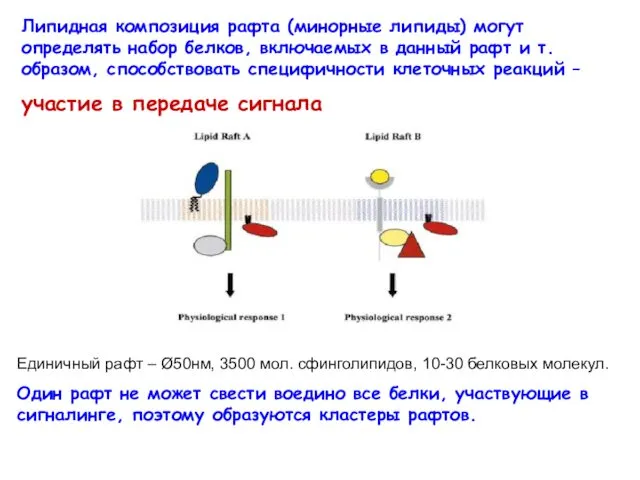
Липидная композиция рафта (минорные липиды) могут определять набор белков, включаемых в
Липидная композиция рафта (минорные липиды) могут определять набор белков, включаемых в
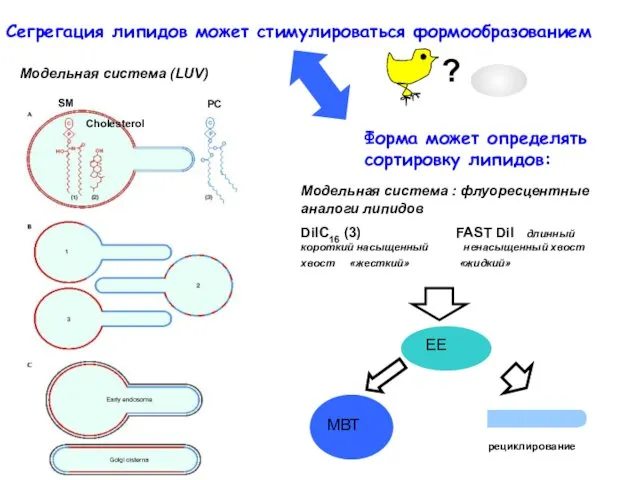
Сегрегация липидов может стимулироваться формообразованием
Форма может определять сортировку липидов:
Сегрегация липидов может стимулироваться формообразованием
Форма может определять сортировку липидов:
Липидный состав органелл в значительной степени определяется формой органеллы
Структура конкретной липидной
Липидный состав органелл в значительной степени определяется формой органеллы
Структура конкретной липидной
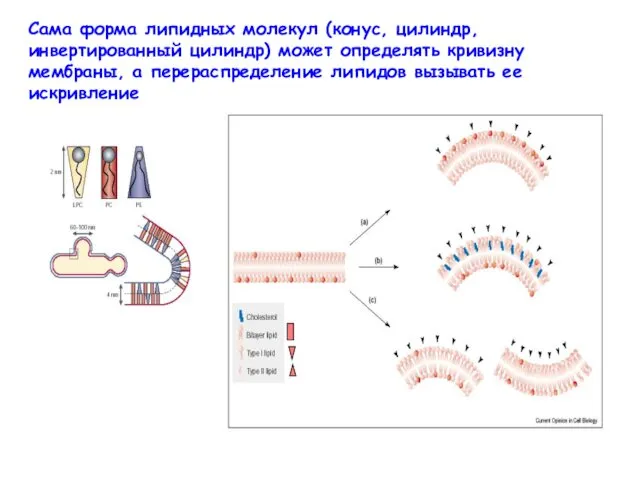
Сама форма липидных молекул (конус, цилиндр, инвертированный цилиндр) может определять кривизну
Сама форма липидных молекул (конус, цилиндр, инвертированный цилиндр) может определять кривизну
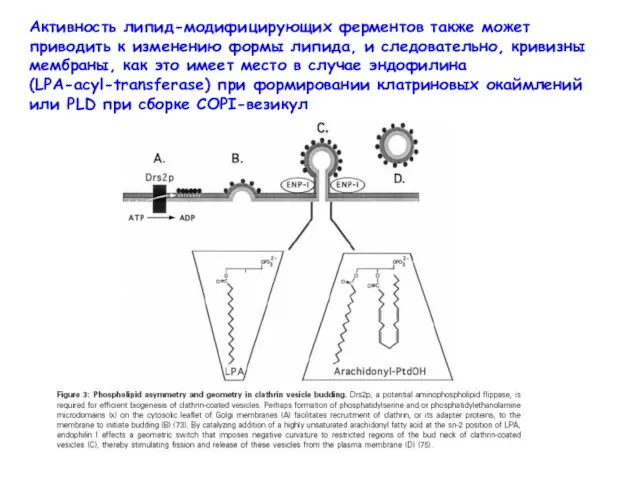
Активность липид-модифицирующих ферментов также может приводить к изменению формы липида, и
Активность липид-модифицирующих ферментов также может приводить к изменению формы липида, и

Как правило, искривление мембран происходит именно в области рафтов, что вовлекает
Как правило, искривление мембран происходит именно в области рафтов, что вовлекает
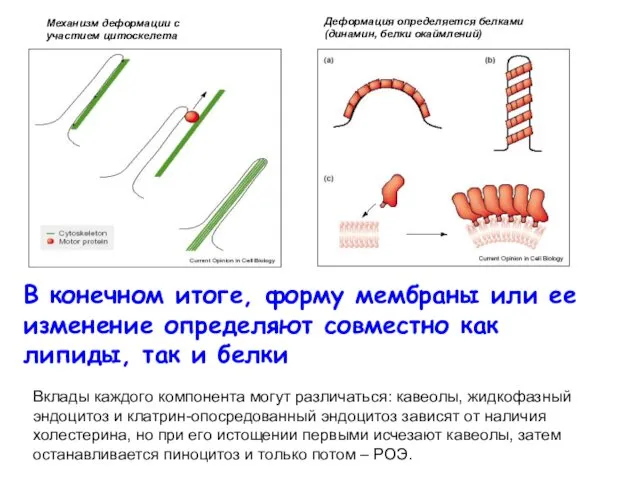
В конечном итоге, форму мембраны или ее изменение определяют совместно как
В конечном итоге, форму мембраны или ее изменение определяют совместно как
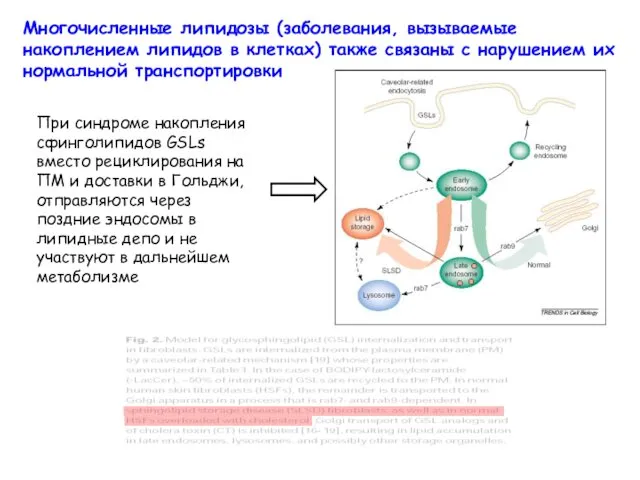
Многочисленные липидозы (заболевания, вызываемые накоплением липидов в клетках) также связаны с
Многочисленные липидозы (заболевания, вызываемые накоплением липидов в клетках) также связаны с

Фосфатидилинозитиды (PtdIns)
играют важную регуляторную роль
в сигнальных и транспортных
процессах
Фосфатидилинозитиды (PtdIns)
играют важную регуляторную роль
в сигнальных и транспортных
процессах
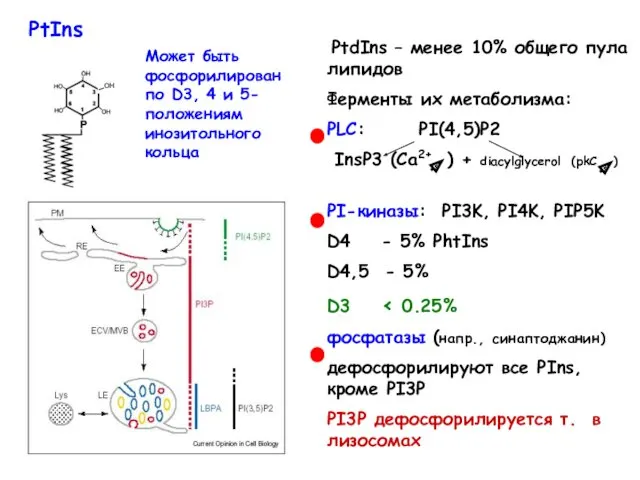
Может быть фосфорилирован по D3, 4 и 5-положениям инозитольного кольца
PtIns
Может быть фосфорилирован по D3, 4 и 5-положениям инозитольного кольца
PtIns
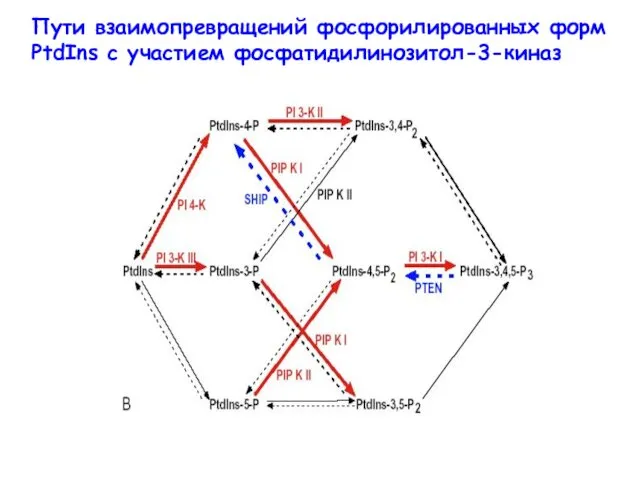
Пути взаимопревращений фосфорилированных форм PtdIns с участием фосфатидилинозитол-3-киназ
Пути взаимопревращений фосфорилированных форм PtdIns с участием фосфатидилинозитол-3-киназ
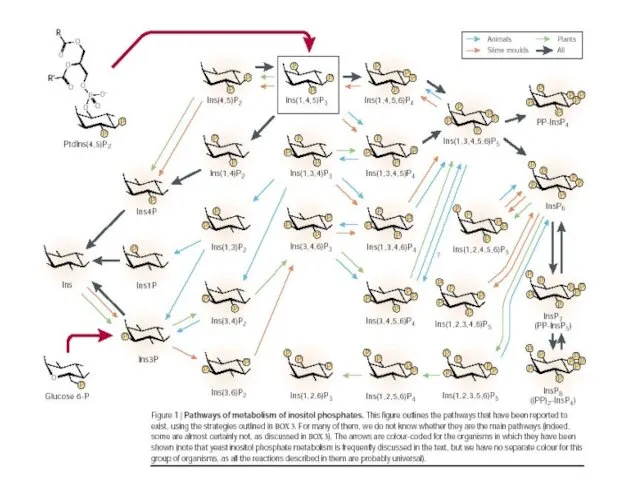
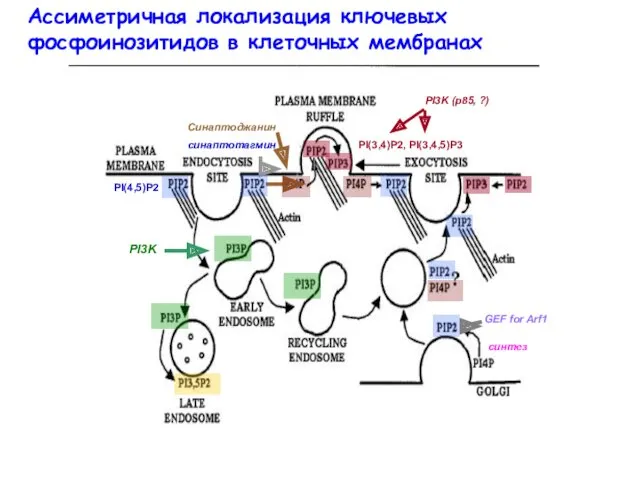
Ассиметричная локализация ключевых фосфоинозитидов в клеточных мембранах
PI(4,5)P2
Синаптоджанин
синаптотагмин
PI(3,4)P2, PI(3,4,5)P3
PI3K (p85, ?)
cинтез
GEF for
Ассиметричная локализация ключевых фосфоинозитидов в клеточных мембранах
PI(4,5)P2
Синаптоджанин
синаптотагмин
PI(3,4)P2, PI(3,4,5)P3
PI3K (p85, ?)
cинтез
GEF for
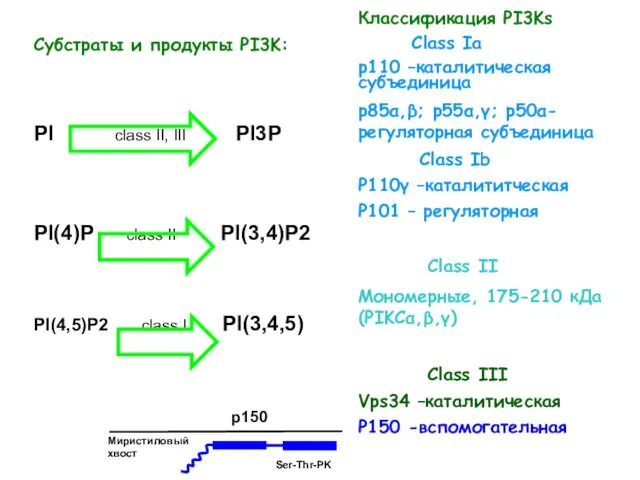
Классификация PI3Ks
Class Ia
р110 –каталитическая субъединица
р85α,β; р55α,γ; p50α-регуляторная субъединица
Class Ib
P110γ
Классификация PI3Ks
Class Ia
р110 –каталитическая субъединица
р85α,β; р55α,γ; p50α-регуляторная субъединица
Class Ib
P110γ
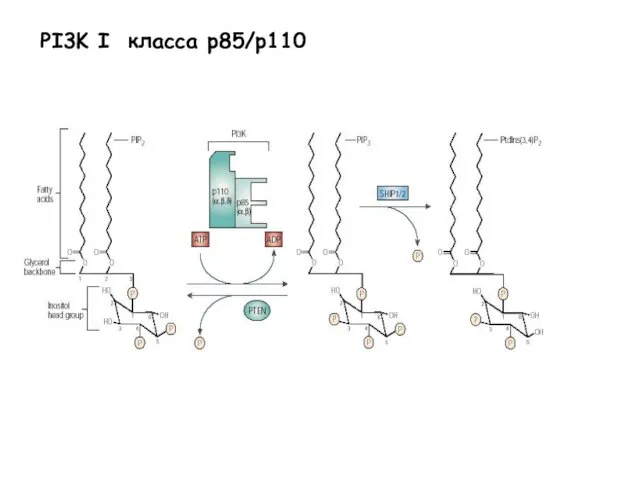
PI3K I класса p85/p110
PI3K I класса p85/p110
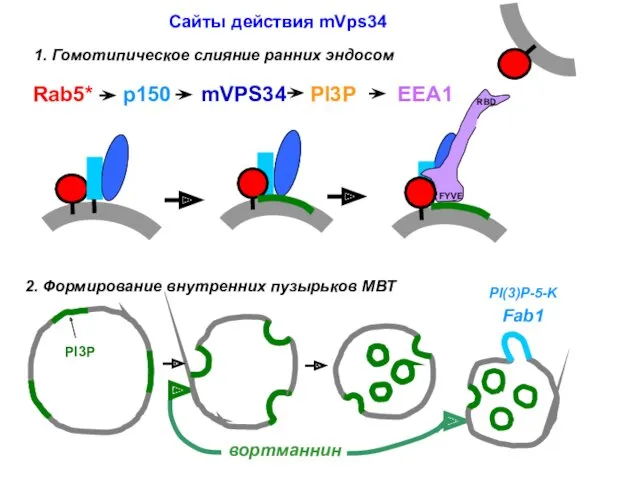
Сайты действия mVps34
1. Гомотипическое слияние ранних эндосом
2. Формирование внутренних пузырьков МВТ
Сайты действия mVps34
1. Гомотипическое слияние ранних эндосом
2. Формирование внутренних пузырьков МВТ
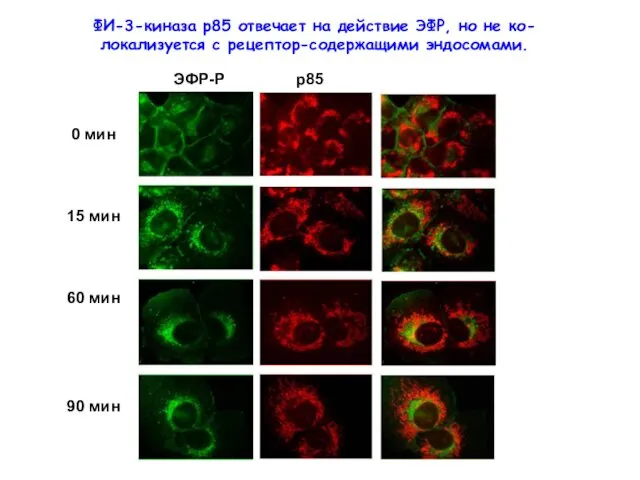
ФИ-3-киназа р85 отвечает на действие ЭФР, но не ко-локализуется с рецептор-содержащими
ФИ-3-киназа р85 отвечает на действие ЭФР, но не ко-локализуется с рецептор-содержащими
 Форматы команд и способы адресации ЭВМ
Форматы команд и способы адресации ЭВМ Краудсорсинг, как инновация для реализации проектов по развитию городской среды
Краудсорсинг, как инновация для реализации проектов по развитию городской среды 20240122_ponyatie_algoritma
20240122_ponyatie_algoritma Концепция структуры и содержания образования по физической культуре в школе
Концепция структуры и содержания образования по физической культуре в школе Значение витаминов в организме человека
Значение витаминов в организме человека Таблица сравнения. Тайный покупатель
Таблица сравнения. Тайный покупатель ЭКО МАРКЕТ. Сеть магазинов Компас Здоровья натуральных продуктов и косметики
ЭКО МАРКЕТ. Сеть магазинов Компас Здоровья натуральных продуктов и косметики Prezentatsia_Gauya
Prezentatsia_Gauya Сидоренко
Сидоренко Конфликты в обществе
Конфликты в обществе Конфликт. Источники конфликта. Разновидности конфликтов. Типы конфликтных личностей. Способы выхода из конфликта
Конфликт. Источники конфликта. Разновидности конфликтов. Типы конфликтных личностей. Способы выхода из конфликта Светский, рациональный характер культуры: литература и искусство
Светский, рациональный характер культуры: литература и искусство Деревья нашего края. Подготовительная группа
Деревья нашего края. Подготовительная группа Организация самостоятельной работы обучающихся на уроках химии в 9 классе
Организация самостоятельной работы обучающихся на уроках химии в 9 классе Реконструктивно-пластические операции при огнестрельно-взрывных ранениях
Реконструктивно-пластические операции при огнестрельно-взрывных ранениях Урок химии в 8 классе по теме Химические уравнения
Урок химии в 8 классе по теме Химические уравнения Іван Драч Крила
Іван Драч Крила Внеклассное мероприятие по географии: игра Юные знатоки географии
Внеклассное мероприятие по географии: игра Юные знатоки географии Презентация 8 марта
Презентация 8 марта Автоматизация и дифференциация звуков Л - ЛЬ
Автоматизация и дифференциация звуков Л - ЛЬ Загадки на логопедическом занятии (Полуэктова Е. Н. - учитель-логопед)
Загадки на логопедическом занятии (Полуэктова Е. Н. - учитель-логопед) Аксиально-поршневые гидромашины. (Лекция 5)
Аксиально-поршневые гидромашины. (Лекция 5) Интенсивная терапия после абдоминальных операций
Интенсивная терапия после абдоминальных операций Конституция Российской Федерации классный час
Конституция Российской Федерации классный час Славные люди нашего края. Колетвинов Николай Михайлович
Славные люди нашего края. Колетвинов Николай Михайлович KUMA_презентация
KUMA_презентация Первое родительское собрание
Первое родительское собрание Презентация к родительскому собранию по ОРКСЭ 4 класс
Презентация к родительскому собранию по ОРКСЭ 4 класс