Механизмы окислительной модификации макромолекул. Материал для студентов ЛФ(стоматология) презентация
- Механизмы окислительной модификации макромолекул. Материал для студентов ЛФ(стоматология)

Содержание
- 2. Предмет изучения: - понятие об окислительной модификации мак- ромолекул (ОММ), локализации и механизмах, - основные субстраты
- 3. Место ОММ в разделах биохимии Статическая биохимия – раздел о химическом составе организма, структуре и свойствах
- 5. ОКИСЛИТЕЛЬНЫЙ СТРЕСС (ОС)
- 6. Ферментативные (дыхательная цепь МХ, микросомальное окисление, респираторный взрыв, ксантиноксидаза и др.) Неферментативные (свободные радикалы, АФК, ионы
- 8. I КЛАСС ферментов ОКСИДОРЕДУКТАЗЫ катализ окислительно-восстановительных реакций, лежащих в основе биологического окисления, микросомального окисления, перекисного окисления.
- 9. Роль кофакторов в ОВР 1) кофакторы хромопротеинов Сочетание белков с окрашенными веществами: флаво-, гемо-, ретинальпротеины и
- 10. ГЕМ цитохромов способен осуществлять перемещение 1ȇ путём изменения валентности железа: Fe2+→Fe3+, Fe3+ → Fe2+
- 11. Витамин B2 (рибофлавин) переносит 2Н
- 13. 2) витамин РР- никотинамидные кофакторы переносят 1Н и 1ê НАД+ → НАДН НАДФ+ → НАДФН
- 14. СУБСТРАТЫ процессов ОММ Субстраты-макромолекулы: - белки (АК, полипептидная цепь), - нуклеиновые кислоты (азотистые основания, D-рибоза цепи
- 15. Жизнь – водная форма существования белковых тел Информация об организме записана в генах (ДНК). Реализация информации
- 16. 1. Аминокислоты с неполярными или гидрофобными R-группами: Gly, Ala, Val, Leu, Ile, Met, Phe, Trp, Pro
- 17. 2. Аминокислоты с полярными незаряженными R-группами: Ser, Thr, Cys, Tyr, Asn, Gln
- 18. 3. Аминокислоты с полярными заряженными R-группами: (-) заряд Asp, Glu; (+) заряд Arg, Lys, His
- 19. Роль О-содержащих АК (норма) 1) фосфопротеины Белки фосфорилируются через боковые радикалы аминокислот, имеющих ОН-группу: СЕР, ТРЕ,
- 20. 2) гликопротеины, протеогликаны Содержат углеводную часть, соединённую с белком ковалентно через боковые радикалы СЕР, ТРЕ (атом
- 21. N- и O-гликозидные связи
- 22. Широко распространён узел белка и углеводной части ГП посредством О-связи СЕР с дисахаридом (N-ацетилгалактозамин-галактоза-…)
- 23. Гликозаминогликан - связующий трисахарид (КОР) - серин кóрового белка Связь ГАГ-полисахаридов с кóровым белком: 1. О-гликозидная
- 24. В норме в коллагене-I необходимо окисление ПРО, ЛИЗ : 1) ПРО→ОН-ПРО до ~1/4 АК, стабилизируют II-структуру;
- 25. Гидроксилирование остатков ПРО в α-цепи проколлагена с получением 4-ОН-ПРО (реакция для получения 5-ОН-ЛИЗ аналогична )
- 26. Сшивки коллагена, сформированные гидроксиЛИЗ и гидроксиаль-ЛИЗ Сшивки коллагена, сформированные гидроксиЛИЗ и гидроксиаль-ЛИЗ
- 27. Окисление ароматических аминокислот (норма)
- 28. Окисление остатков ЦИС → дисульфидные мосты
- 29. В ходе синтеза белка SS-связи образуются при спонтанном окислении SH-групп сближающихся остатков ЦИС первичной структуры. Особо
- 30. Фолдинг – спонтанное сворачивание синтезированной полипептидной цепи в уникальную пространственную структуру: на внешней поверхности глобулы белка
- 32. ДЕНАТУРАЦИЯ БЕЛКА при ОС окисление и осаждение белка с нарушением пространственной структуры и потерей биологических свойств.
- 33. Обратимое окисление и РЕНАТУРАЦИЯ При ренатурации белок вновь сворачивается в нативную конформацию и его биологическая активность
- 34. Обратимое окисление белков по остаткам МЕТ и ЦИС при ОС
- 36. Вариант окисления ЦИС как свободной АК
- 37. НЕОБРАТИМОЕ окисление в белках В отличие от дисульфидов и сульфенов невозможно вос-становить до SH-группы сульфиновые и
- 38. НЕобратимое окисление белков Получение свободнорадикальных продуктов при атаке по аромати-ческим боковым цепям АК, как в составе
- 39. Окисление белковой цепи, боковых радикалов АК ведёт к необратимой модификации – карбонилированию АРГ → алутаминовый полуальдегид
- 40. ЛИЗ АРГ ГЛУ
- 41. Пролин при окислительном стрессе КАРБОНИЛИРОВАНИЕ ЦЕПИ и ПРОЛИН При избыточном окислении пептидной цепи обычно происходит разрыв
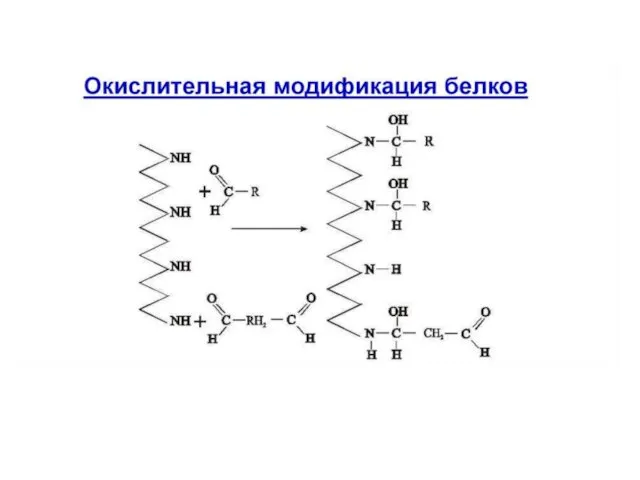
- 42. Окислительная модификация белковой цепи
- 44. Альфа-амидный и диамидный пути разрыва пептидной цепи
- 45. Металлопротеины Содержат ионы одного или нескольких металлов. Характерна связь ионов с Асп, Глу, Цис, Гис белка
- 46. Fe(II) - Fe(III) и другие металлы с переменной валентностью Белки-металлопротеины Связывание металлов предупреждает металл-катализируемое окисление (МКО)
- 47. металлы с переменной валентностью
- 48. Пространственное строение комплексов металл-трансферрин Строение церулоплазмина (содержит Cu, играет роль СОД плазмы крови) Белок лактоферрин из
- 49. строение гемоглобина и миоглобина гемоглобин (а), его субъединица (б), Расположение гема и белковой части в миоглобине
- 50. Связь гема, глобина, молекулы кислорода (без доставки О2 невозможно окисление)
- 51. Удаление окисленных белков Необратимо повреждённые белки могут разрушаться несколькими способами: 1) протеазы (с разной субстратной специфичностью)
- 52. Убиквитинирование (убиквитинилирование) — присоединение убиквитинлигазами 1-го или нескольких мономеров убиквитина ковалентной связью к боковым аминогруппам лизина
- 53. Убиквитин из 76 АК ubuque (лат.) вездесущий
- 54. Нуклеопротеины и окислительная модификация нуклеиновых кислот (ОМНК) Апобелки НП защищают нуклеиновые кислоты от окисления и иных
- 55. АПОБЕЛКИ НП – гистоны и протамины протамины - простые белки, не содержат серы, у некоторых видов
- 56. Редокс-чувствительная мишень для ОН-радикалов: -СН2- группы дезокси-рибозы ДНК
- 57. При ОС: критично повреждение азотистых оснований ДНК ОН-радикалами
- 58. Повреждения ДНК могут быть без окисления. Пример – депуринизация дезоксирибозы
- 59. Подробно механизмы повреждения – на практическом занятии
- 60. Сшивки ДНК
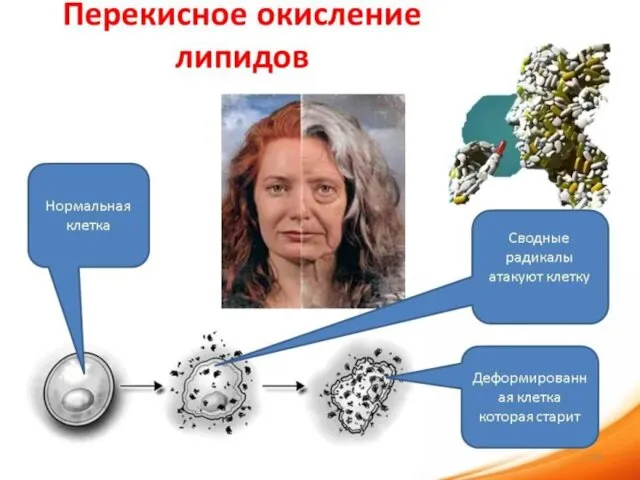
- 61. Окислительная модификация липидов – это перекисное окисление липидов (ПОЛ) Плазматическая мембрана клетки и транспортные липопротеины содержат
- 62. Фосфолипиды – основа мембраны (глицеро- и сфингофосфолипиды)
- 63. Фосфолипиды транспортных ЛП крови
- 64. Липопротеины (ЛП) 1) транспортные ЛП крови – надмолекулярные структуры, содержат все классы липидов и белки, контакт
- 65. Пример: наружная стенка бледной трепонемы состоит из липопротеинов и белков. Липопротеины поддер- живают механическую целостность мембраны,
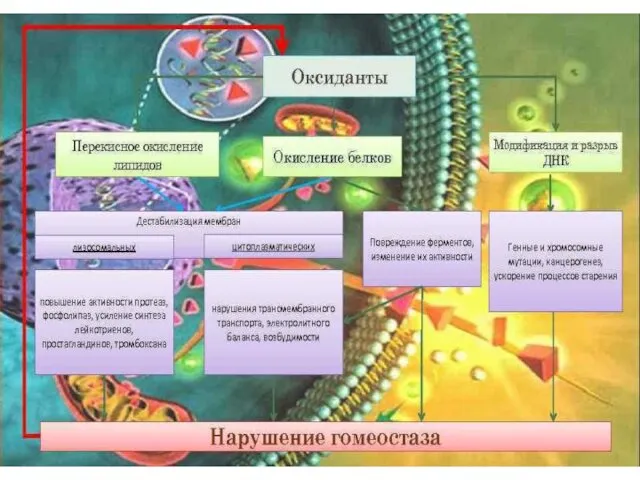
- 66. Окислительный стресс
- 67. Полиненасыщенные жирные кислоты (вит F)
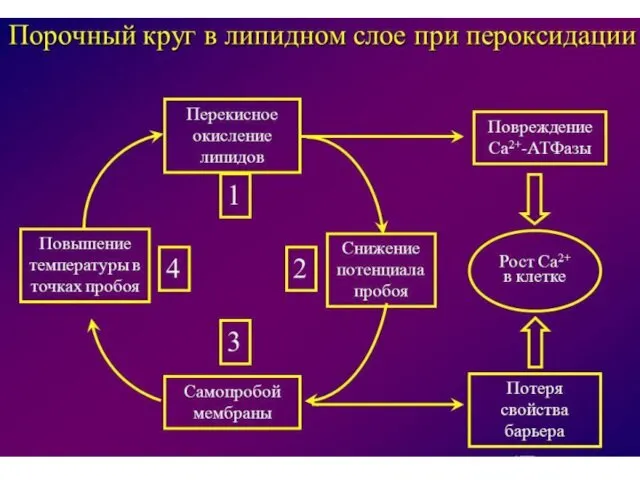
- 68. Механизм ПОЛ
- 71. Продукты ПОЛ образуют ковалентные сшивки с белками и липидами мембран
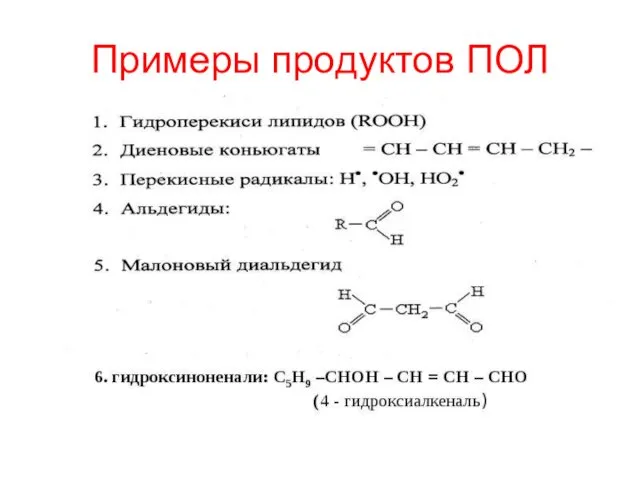
- 74. Примеры продуктов ПОЛ
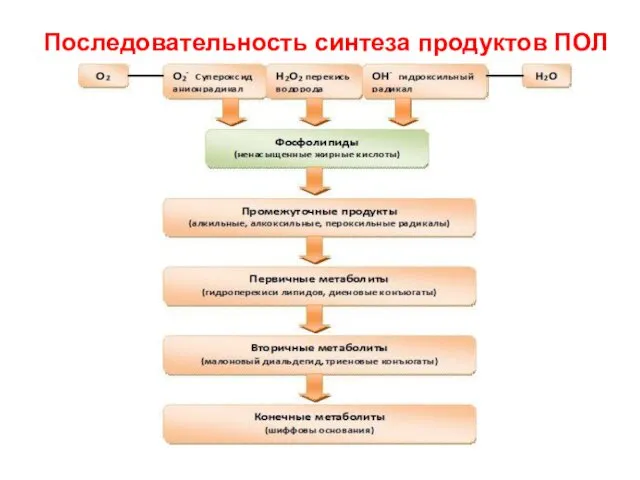
- 75. Последовательность синтеза продуктов ПОЛ
- 76. Влияние промежуточных продуктов ПОЛ на белок может быть обратимым (зависит от дозы и мишени)
- 81. Скачать презентацию

Предмет изучения:
- понятие об окислительной модификации мак-
ромолекул (ОММ), локализации
Предмет изучения: - понятие об окислительной модификации мак- ромолекул (ОММ), локализации

Место ОММ в разделах биохимии
Статическая биохимия – раздел о химическом составе
Место ОММ в разделах биохимии
Статическая биохимия – раздел о химическом составе


ОКИСЛИТЕЛЬНЫЙ СТРЕСС (ОС)
ОКИСЛИТЕЛЬНЫЙ СТРЕСС (ОС)

Ферментативные (дыхательная цепь МХ, микросомальное окисление, респираторный взрыв, ксантиноксидаза и др.)
Неферментативные
Ферментативные (дыхательная цепь МХ, микросомальное окисление, респираторный взрыв, ксантиноксидаза и др.)
Неферментативные


I КЛАСС ферментов ОКСИДОРЕДУКТАЗЫ
катализ окислительно-восстановительных реакций, лежащих в основе биологического окисления,
I КЛАСС ферментов ОКСИДОРЕДУКТАЗЫ
катализ окислительно-восстановительных реакций, лежащих в основе биологического окисления,

Роль кофакторов в ОВР
1) кофакторы хромопротеинов
Сочетание белков с окрашенными
Роль кофакторов в ОВР
1) кофакторы хромопротеинов
Сочетание белков с окрашенными
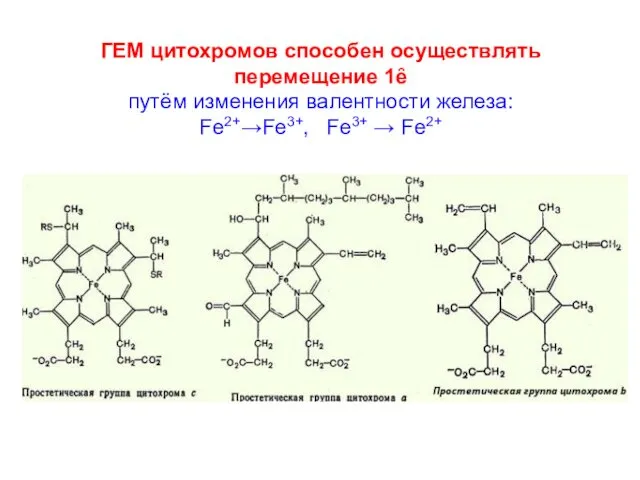
ГЕМ цитохромов способен осуществлять перемещение 1ȇ
путём изменения валентности железа:
Fe2+→Fe3+,
ГЕМ цитохромов способен осуществлять перемещение 1ȇ путём изменения валентности железа: Fe2+→Fe3+,
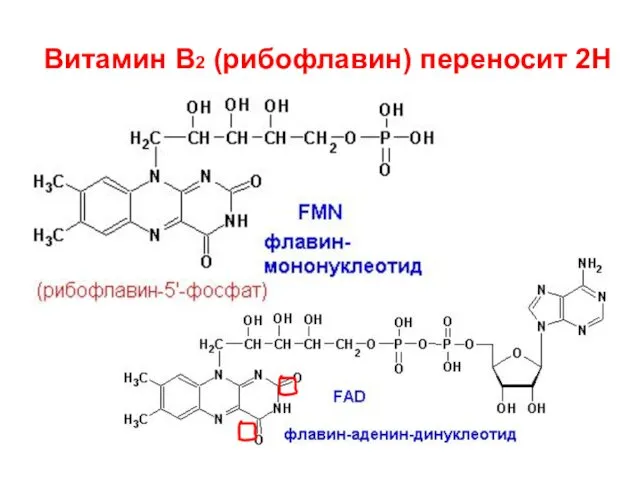
Витамин B2 (рибофлавин) переносит 2Н
Витамин B2 (рибофлавин) переносит 2Н
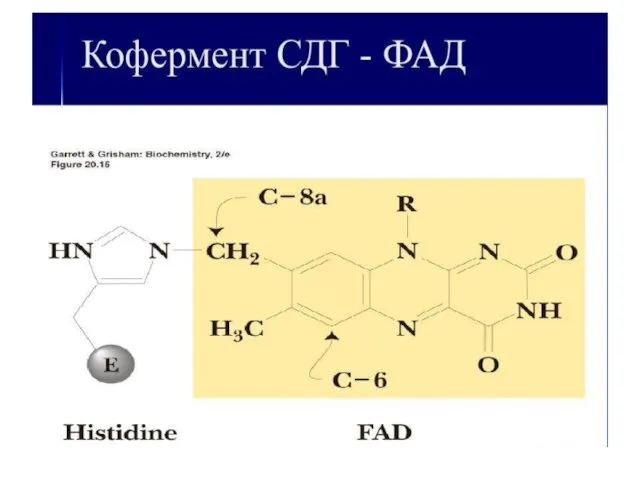
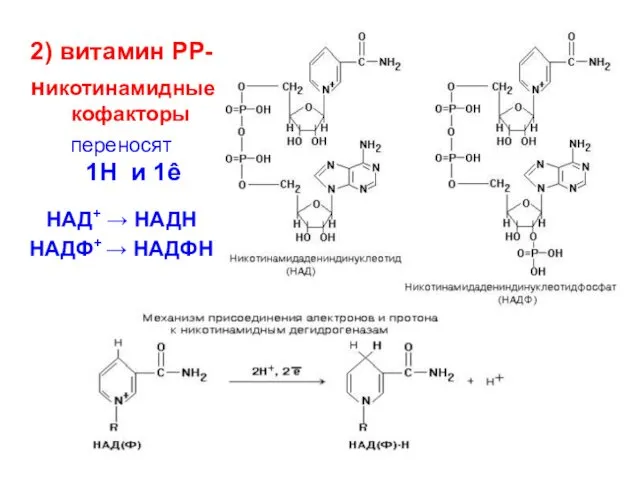
2) витамин РР-
никотинамидные
кофакторы
переносят
1Н и 1ê
НАД+ → НАДН
НАДФ+
2) витамин РР-
никотинамидные
кофакторы
переносят
1Н и 1ê
НАД+ → НАДН
НАДФ+

СУБСТРАТЫ процессов ОММ
Субстраты-макромолекулы:
- белки
(АК, полипептидная цепь),
- нуклеиновые кислоты
(азотистые основания,
СУБСТРАТЫ процессов ОММ
Субстраты-макромолекулы:
- белки
(АК, полипептидная цепь),
- нуклеиновые кислоты
(азотистые основания,

Жизнь – водная форма существования
белковых тел
Информация об организме записана
в
Жизнь – водная форма существования
белковых тел
Информация об организме записана в
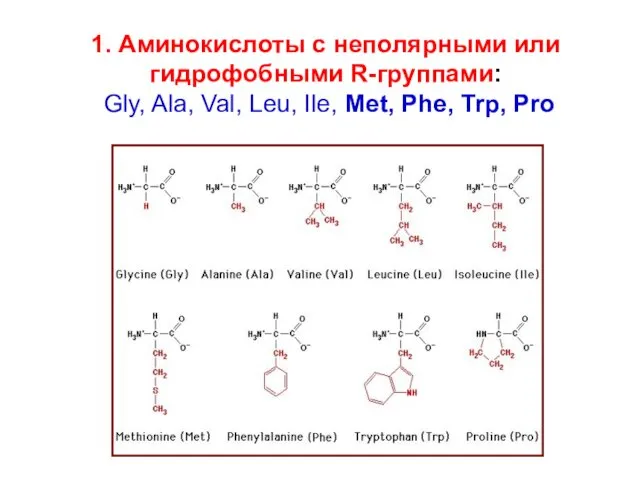
1. Аминокислоты с неполярными или гидрофобными R-группами: Gly, Ala, Val, Leu,
1. Аминокислоты с неполярными или гидрофобными R-группами: Gly, Ala, Val, Leu,
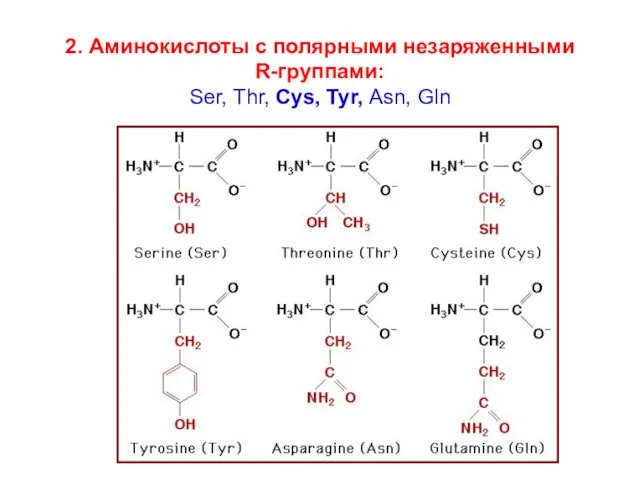
2. Аминокислоты с полярными незаряженными
R-группами:
Ser, Thr, Cys, Tyr, Asn, Gln
2. Аминокислоты с полярными незаряженными
R-группами:
Ser, Thr, Cys, Tyr, Asn, Gln
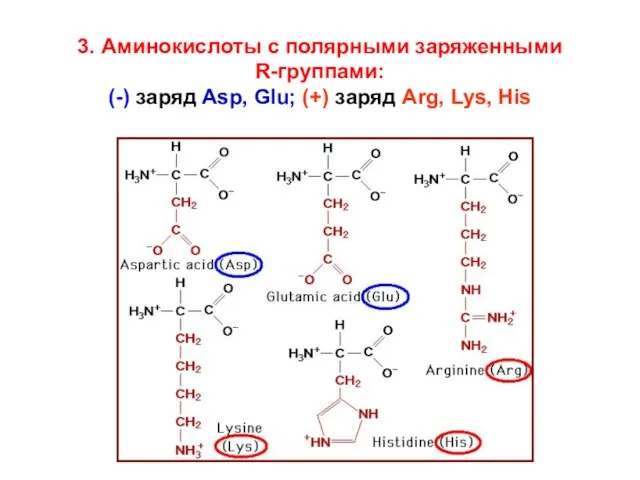
3. Аминокислоты с полярными заряженными
R-группами:
(-) заряд Asp, Glu; (+)
3. Аминокислоты с полярными заряженными R-группами: (-) заряд Asp, Glu; (+)
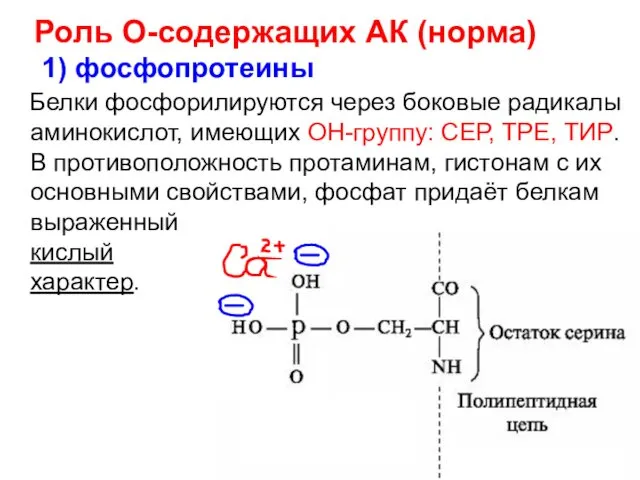
Роль О-содержащих АК (норма)
1) фосфопротеины
Белки фосфорилируются через боковые
Роль О-содержащих АК (норма)
1) фосфопротеины
Белки фосфорилируются через боковые

2) гликопротеины, протеогликаны
Содержат углеводную часть, соединённую с белком ковалентно через боковые
2) гликопротеины, протеогликаны
Содержат углеводную часть, соединённую с белком ковалентно через боковые
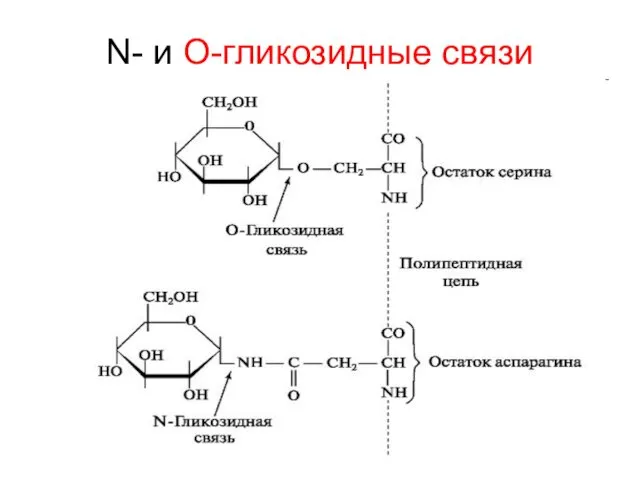
N- и O-гликозидные связи
N- и O-гликозидные связи
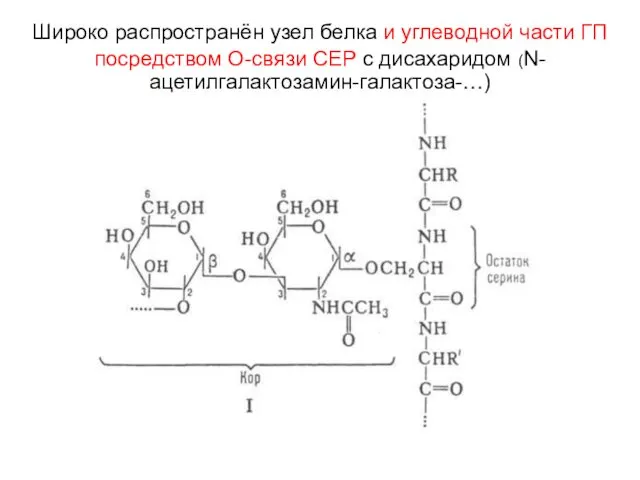
Широко распространён узел белка и углеводной части ГП посредством О-связи СЕР
Широко распространён узел белка и углеводной части ГП посредством О-связи СЕР
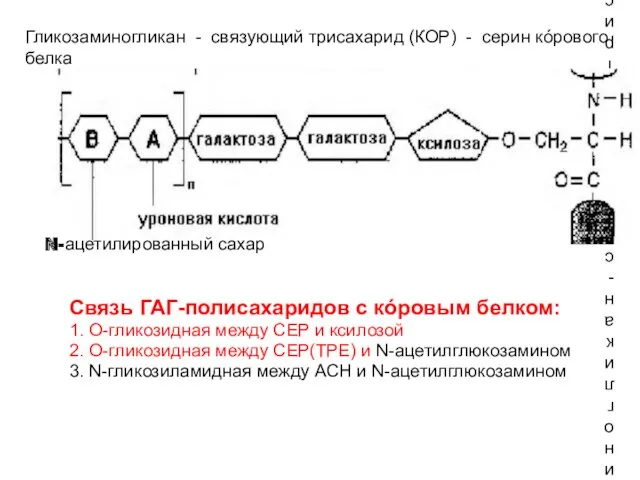
Гликозаминогликан - связующий трисахарид (КОР) - серин кóрового белка
Связь ГАГ-полисахаридов с
Гликозаминогликан - связующий трисахарид (КОР) - серин кóрового белка
Связь ГАГ-полисахаридов с
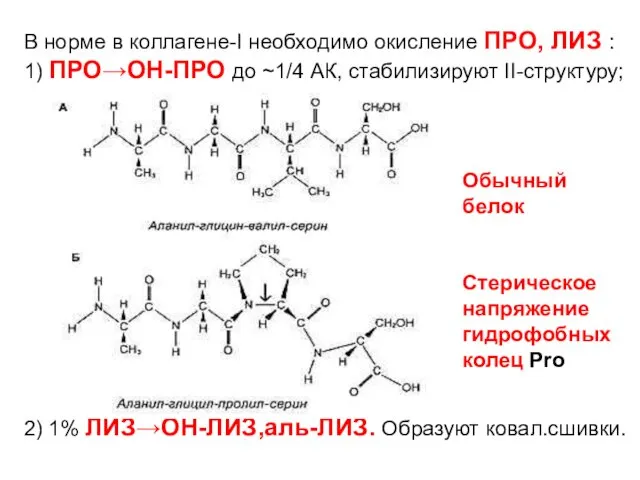
В норме в коллагене-I необходимо окисление ПРО, ЛИЗ :
1) ПРО→ОН-ПРО до
В норме в коллагене-I необходимо окисление ПРО, ЛИЗ :
1) ПРО→ОН-ПРО до
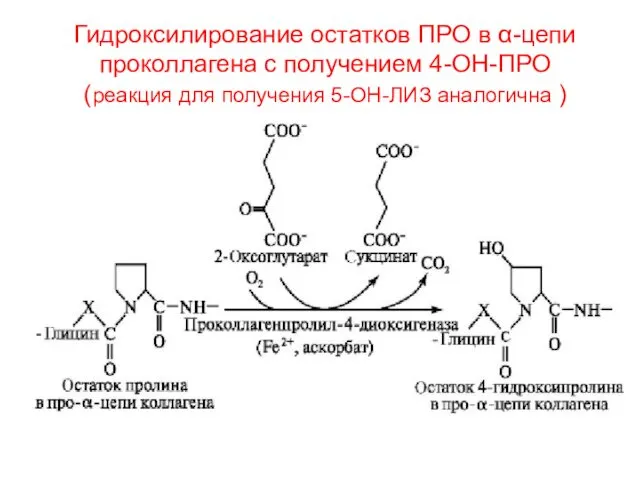
Гидроксилирование остатков ПРО в α-цепи проколлагена с получением 4-ОН-ПРО
(реакция для получения
Гидроксилирование остатков ПРО в α-цепи проколлагена с получением 4-ОН-ПРО (реакция для получения
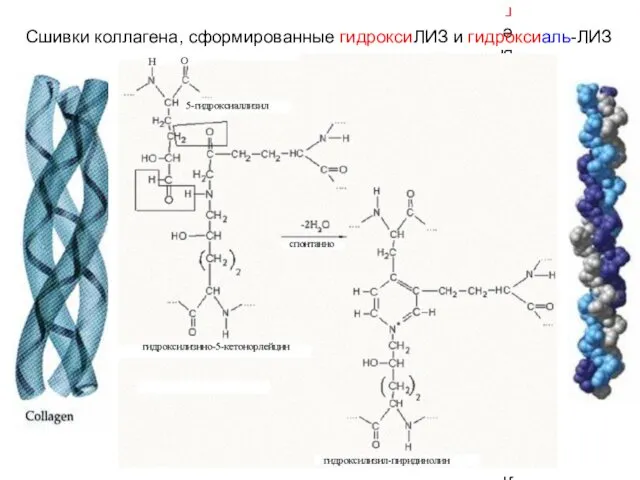
Сшивки коллагена, сформированные гидроксиЛИЗ и гидроксиаль-ЛИЗ
Сшивки коллагена, сформированные гидроксиЛИЗ и гидроксиаль-ЛИЗ
Сшивки коллагена, сформированные гидроксиЛИЗ и гидроксиаль-ЛИЗ
Сшивки коллагена, сформированные гидроксиЛИЗ и гидроксиаль-ЛИЗ
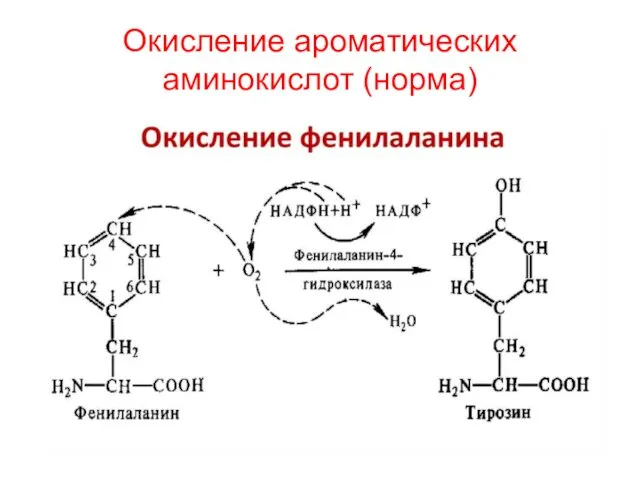
Окисление ароматических аминокислот (норма)
Окисление ароматических аминокислот (норма)
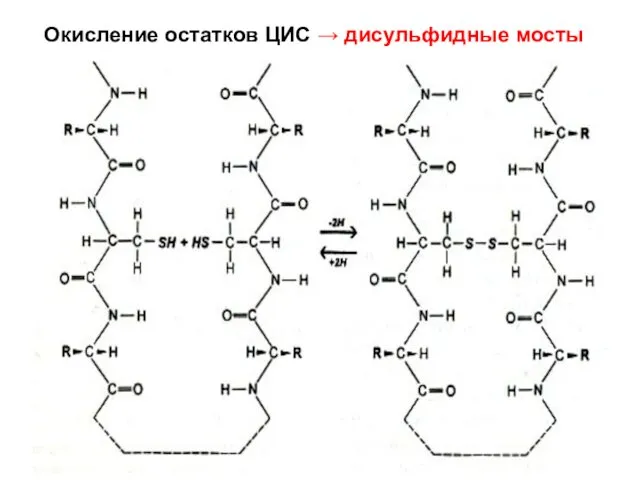
Окисление остатков ЦИС → дисульфидные мосты
Окисление остатков ЦИС → дисульфидные мосты

В ходе синтеза белка SS-связи образуются при спонтанном окислении SH-групп сближающихся

Фолдинг – спонтанное сворачивание синтезированной полипептидной цепи
в уникальную пространственную структуру:
Фолдинг – спонтанное сворачивание синтезированной полипептидной цепи в уникальную пространственную структуру:
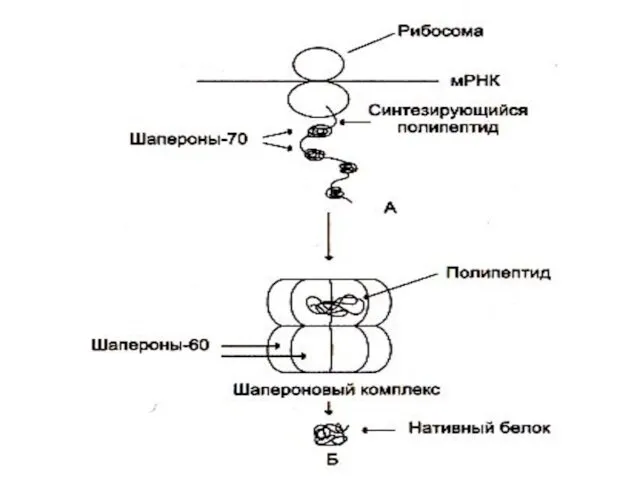

ДЕНАТУРАЦИЯ БЕЛКА при ОС
окисление и осаждение белка с
ДЕНАТУРАЦИЯ БЕЛКА при ОС
окисление и осаждение белка с

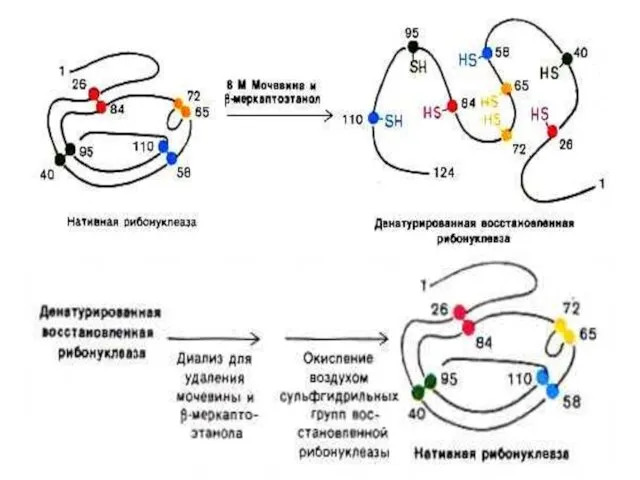
Обратимое окисление и РЕНАТУРАЦИЯ
При ренатурации белок вновь сворачивается в нативную
Обратимое окисление и РЕНАТУРАЦИЯ
При ренатурации белок вновь сворачивается в нативную
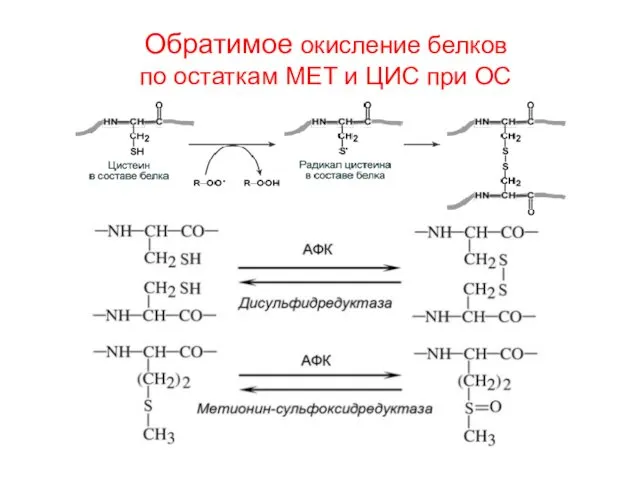
Обратимое окисление белков
по остаткам МЕТ и ЦИС при ОС
Обратимое окисление белков
по остаткам МЕТ и ЦИС при ОС
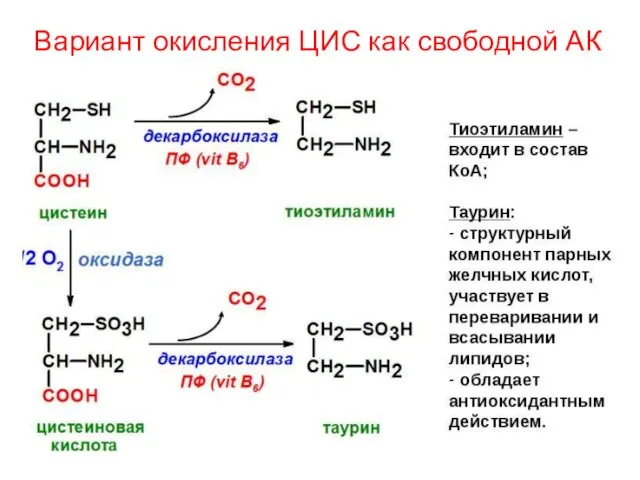
Вариант окисления ЦИС как свободной АК
Вариант окисления ЦИС как свободной АК
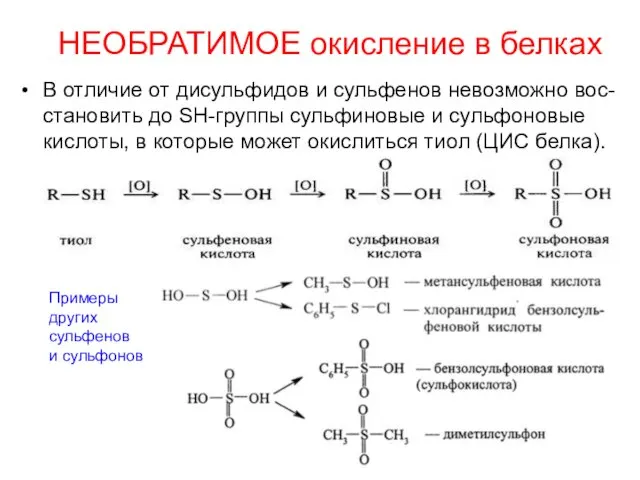
НЕОБРАТИМОЕ окисление в белках
В отличие от дисульфидов и сульфенов невозможно вос-становить
НЕОБРАТИМОЕ окисление в белках
В отличие от дисульфидов и сульфенов невозможно вос-становить
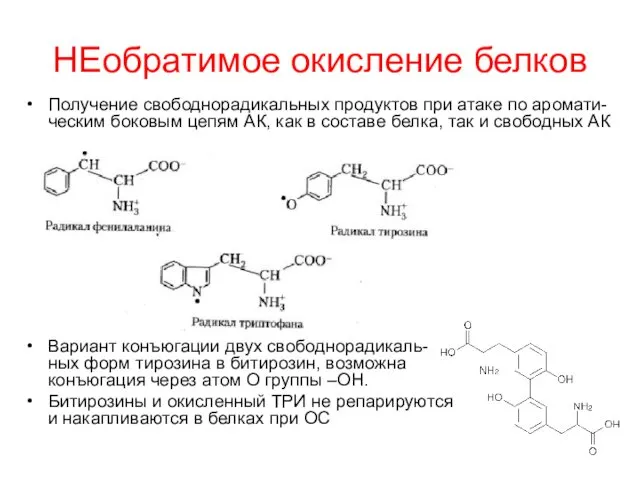
НЕобратимое окисление белков
Получение свободнорадикальных продуктов при атаке по аромати-ческим боковым цепям
НЕобратимое окисление белков
Получение свободнорадикальных продуктов при атаке по аромати-ческим боковым цепям
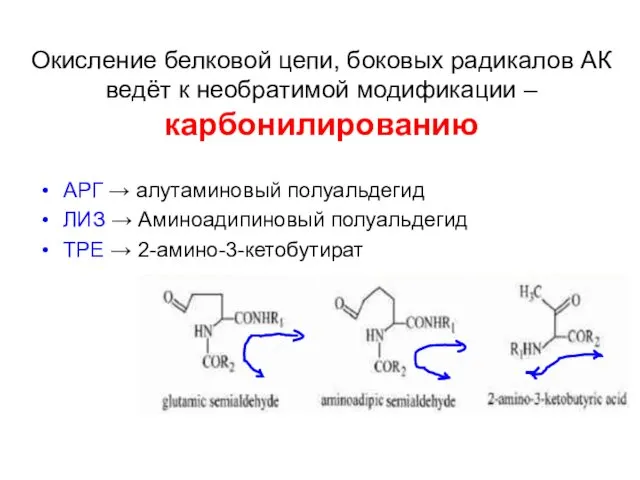
Окисление белковой цепи, боковых радикалов АК
ведёт к необратимой модификации –
Окисление белковой цепи, боковых радикалов АК ведёт к необратимой модификации –
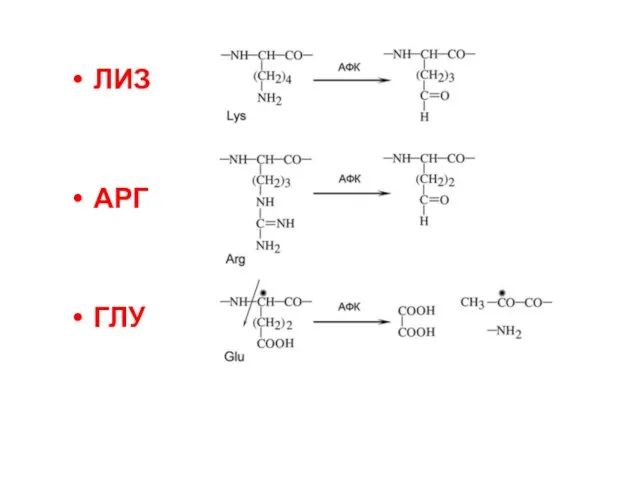
ЛИЗ
АРГ
ГЛУ
ЛИЗ
АРГ
ГЛУ
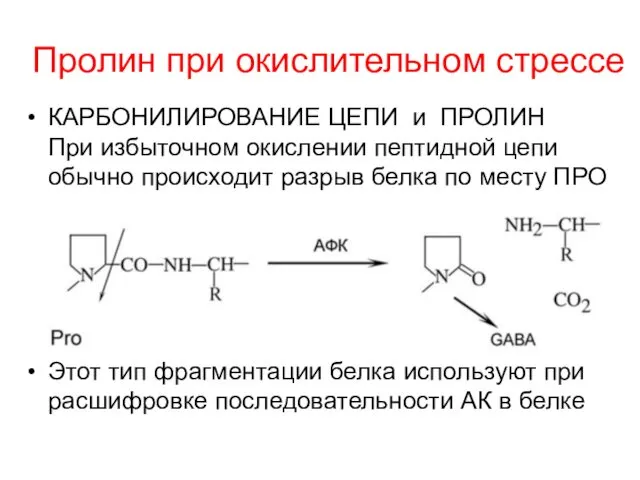
Пролин при окислительном стрессе
КАРБОНИЛИРОВАНИЕ ЦЕПИ и ПРОЛИН
При избыточном окислении пептидной цепи
Пролин при окислительном стрессе
КАРБОНИЛИРОВАНИЕ ЦЕПИ и ПРОЛИН При избыточном окислении пептидной цепи
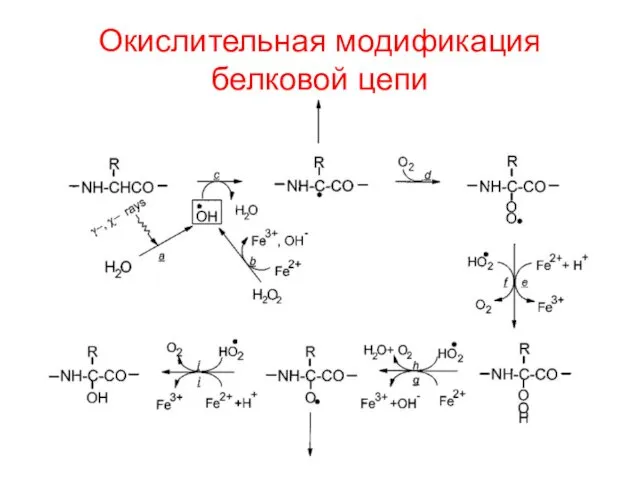
Окислительная модификация белковой цепи
Окислительная модификация белковой цепи
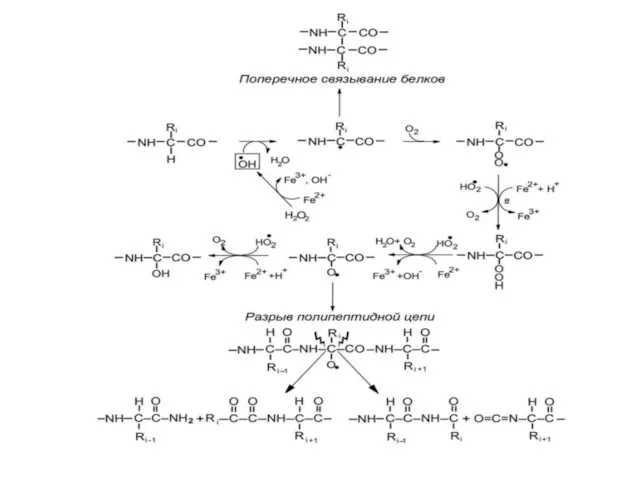
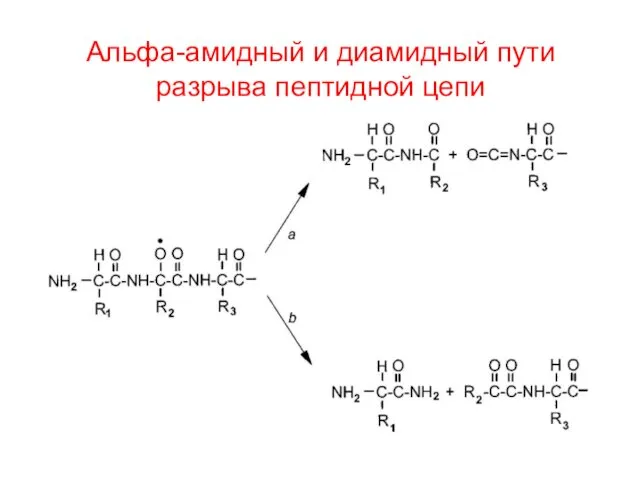
Альфа-амидный и диамидный пути разрыва пептидной цепи
Альфа-амидный и диамидный пути разрыва пептидной цепи

Металлопротеины
Содержат ионы одного или нескольких металлов. Характерна связь ионов с Асп,
Металлопротеины
Содержат ионы одного или нескольких металлов. Характерна связь ионов с Асп,

Fe(II) - Fe(III) и другие металлы
с переменной валентностью
Белки-металлопротеины
Связывание металлов предупреждает
Fe(II) - Fe(III) и другие металлы
с переменной валентностью
Белки-металлопротеины
Связывание металлов предупреждает

металлы с переменной валентностью
металлы с переменной валентностью
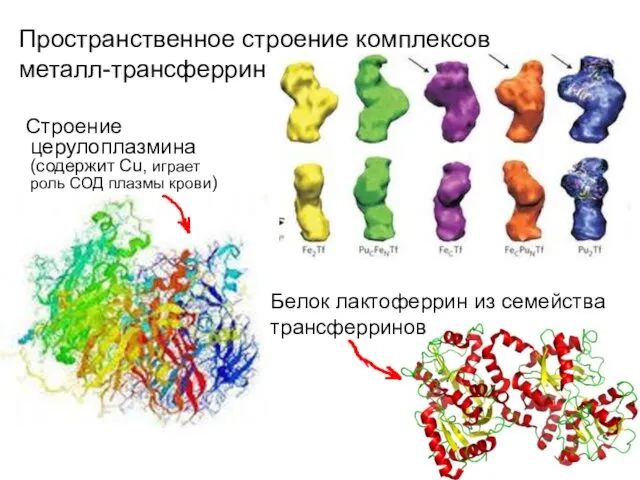
Пространственное строение комплексов
металл-трансферрин
Строение церулоплазмина (содержит Cu, играет роль СОД
Пространственное строение комплексов
металл-трансферрин
Строение церулоплазмина (содержит Cu, играет роль СОД
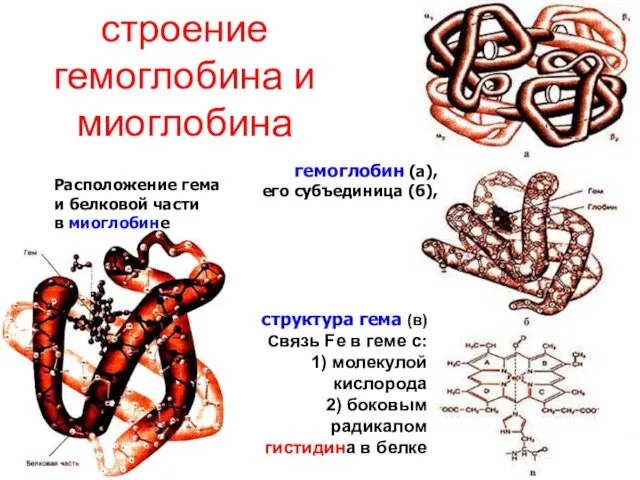
строение гемоглобина и миоглобина
гемоглобин (а),
его субъединица (б),
Расположение гема
и
строение гемоглобина и миоглобина
гемоглобин (а),
его субъединица (б),
Расположение гема и
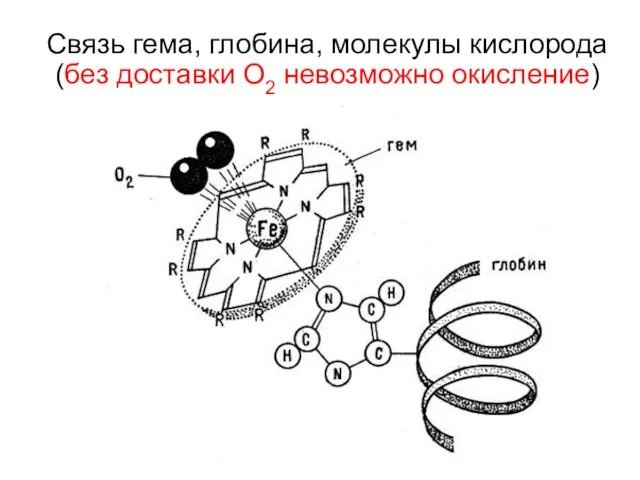
Связь гема, глобина, молекулы кислорода
(без доставки О2 невозможно окисление)
Связь гема, глобина, молекулы кислорода
(без доставки О2 невозможно окисление)

Удаление окисленных белков
Необратимо повреждённые белки могут разрушаться несколькими способами:
1) протеазы (с
Удаление окисленных белков
Необратимо повреждённые белки могут разрушаться несколькими способами: 1) протеазы (с
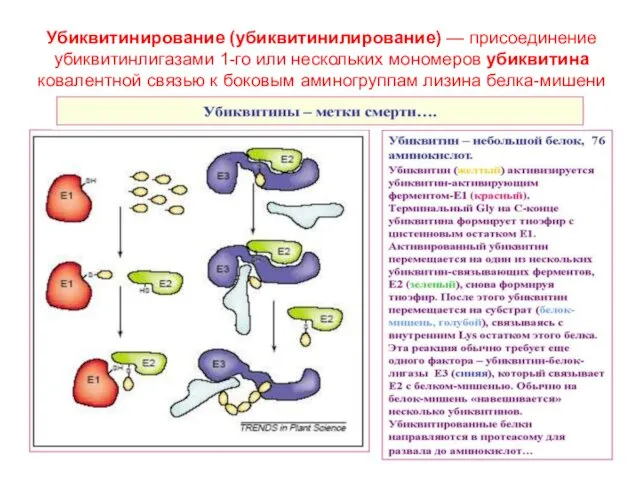
Убиквитинирование (убиквитинилирование) — присоединение убиквитинлигазами 1-го или нескольких мономеров убиквитина ковалентной связью
Убиквитинирование (убиквитинилирование) — присоединение убиквитинлигазами 1-го или нескольких мономеров убиквитина ковалентной связью
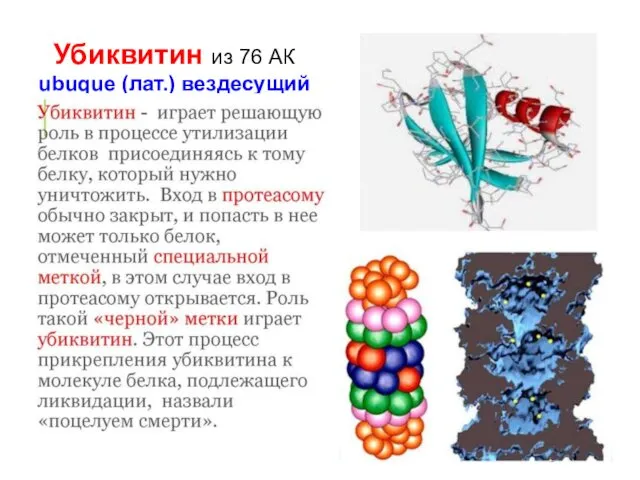
Убиквитин из 76 АК ubuque (лат.) вездесущий
Убиквитин из 76 АК ubuque (лат.) вездесущий

Нуклеопротеины и окислительная модификация нуклеиновых кислот (ОМНК)
Апобелки НП защищают нуклеиновые кислоты
Нуклеопротеины и окислительная модификация нуклеиновых кислот (ОМНК)
Апобелки НП защищают нуклеиновые кислоты
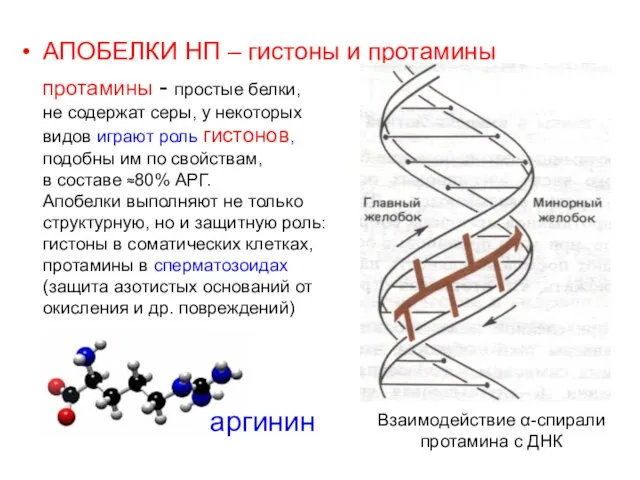
АПОБЕЛКИ НП – гистоны и протамины
протамины - простые белки,
не
АПОБЕЛКИ НП – гистоны и протамины
протамины - простые белки,
не
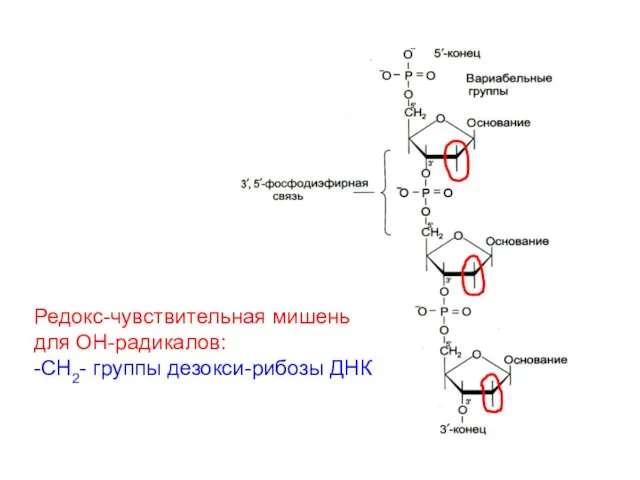
Редокс-чувствительная мишень
для ОН-радикалов:
-СН2- группы дезокси-рибозы ДНК
Редокс-чувствительная мишень
для ОН-радикалов:
-СН2- группы дезокси-рибозы ДНК
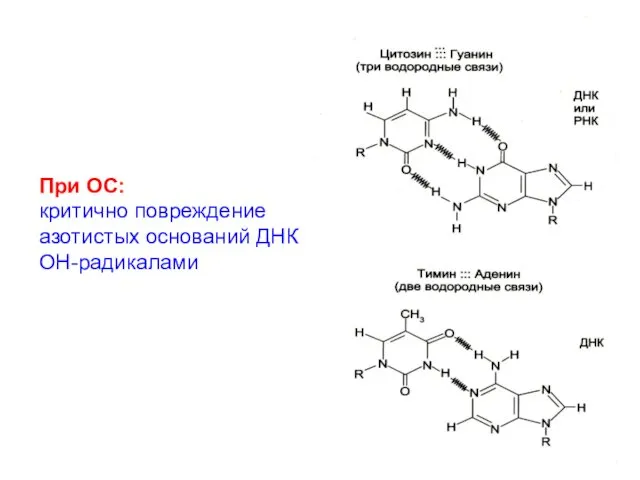
При ОС:
критично повреждение
азотистых оснований ДНК
ОН-радикалами
При ОС:
критично повреждение
азотистых оснований ДНК
ОН-радикалами
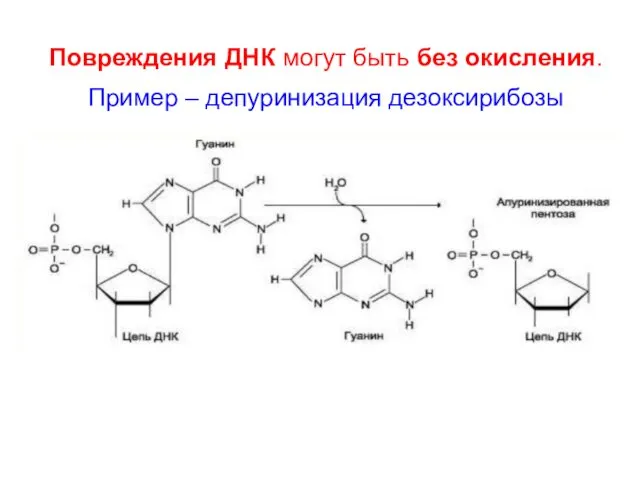
Повреждения ДНК могут быть без окисления. Пример – депуринизация дезоксирибозы
Повреждения ДНК могут быть без окисления. Пример – депуринизация дезоксирибозы
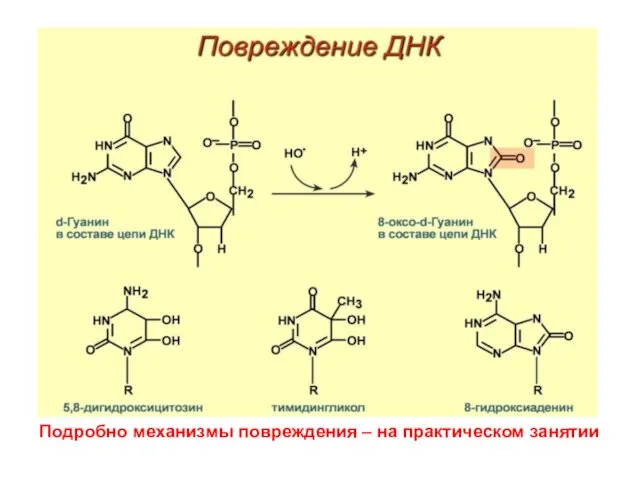
Подробно механизмы повреждения – на практическом занятии
Подробно механизмы повреждения – на практическом занятии
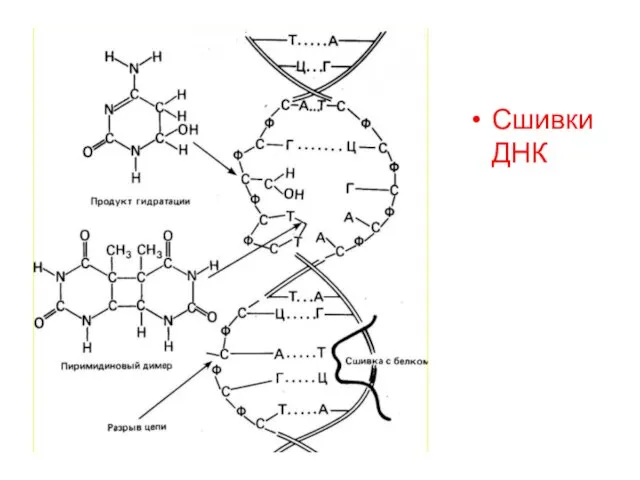
Сшивки ДНК
Сшивки ДНК

Окислительная модификация липидов – это перекисное
окисление липидов (ПОЛ)
Плазматическая мембрана клетки
Окислительная модификация липидов – это перекисное
окисление липидов (ПОЛ)
Плазматическая мембрана клетки
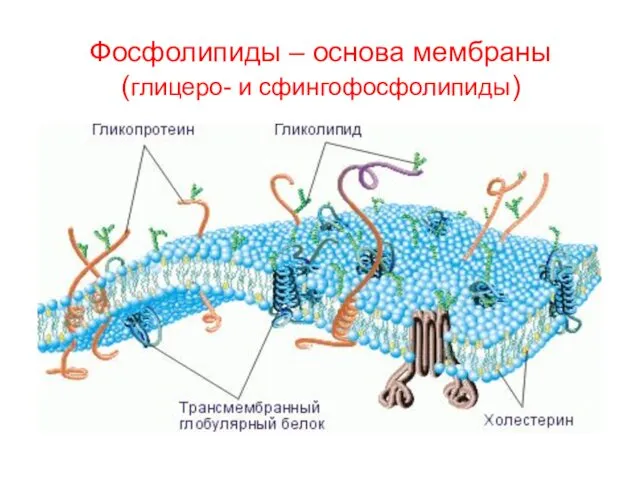
Фосфолипиды – основа мембраны (глицеро- и сфингофосфолипиды)
Фосфолипиды – основа мембраны (глицеро- и сфингофосфолипиды)
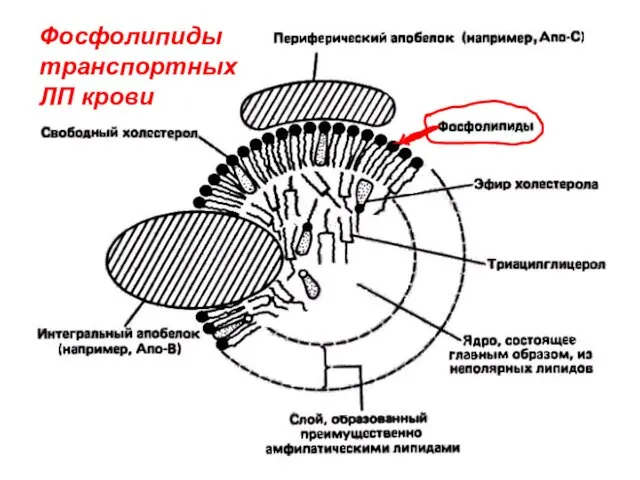
Фосфолипиды
транспортных
ЛП крови
Фосфолипиды
транспортных
ЛП крови

Липопротеины (ЛП)
1) транспортные ЛП крови – надмолекулярные структуры, содержат все классы
Липопротеины (ЛП)
1) транспортные ЛП крови – надмолекулярные структуры, содержат все классы
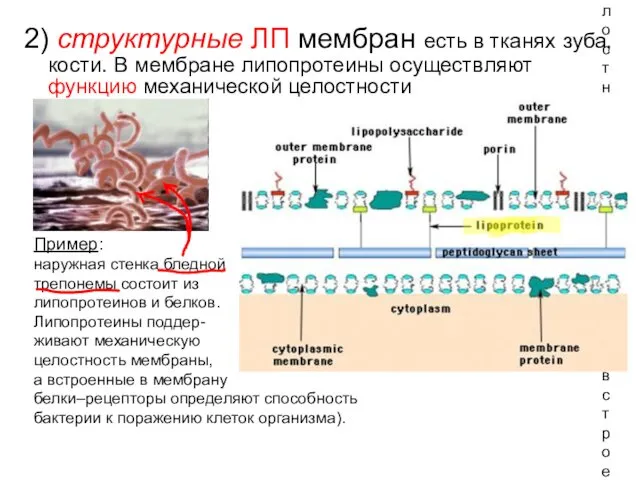
Пример:
наружная стенка бледной
трепонемы состоит из
липопротеинов и белков.
Липопротеины поддер-
живают механическую
целостность мембраны,
а
Пример: наружная стенка бледной трепонемы состоит из липопротеинов и белков. Липопротеины поддер- живают механическую целостность мембраны, а

Окислительный стресс
Окислительный стресс
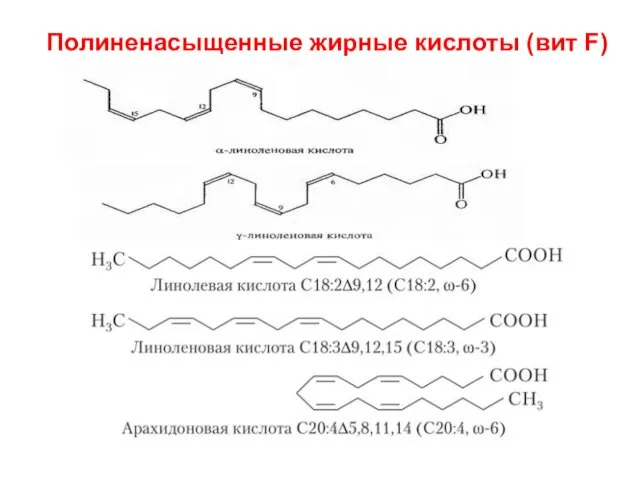
Полиненасыщенные жирные кислоты (вит F)
Полиненасыщенные жирные кислоты (вит F)
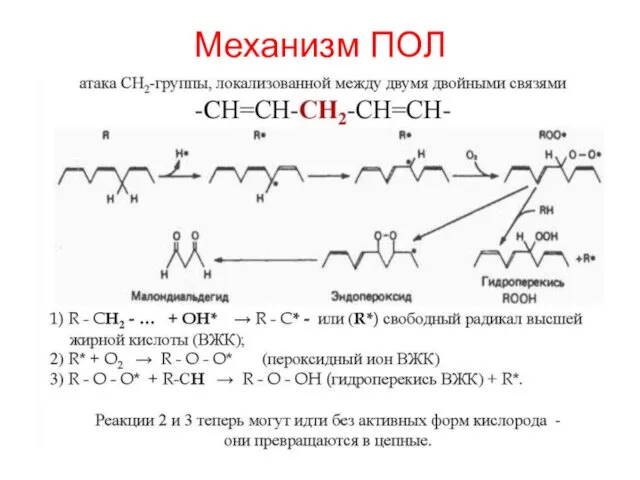
Механизм ПОЛ
Механизм ПОЛ
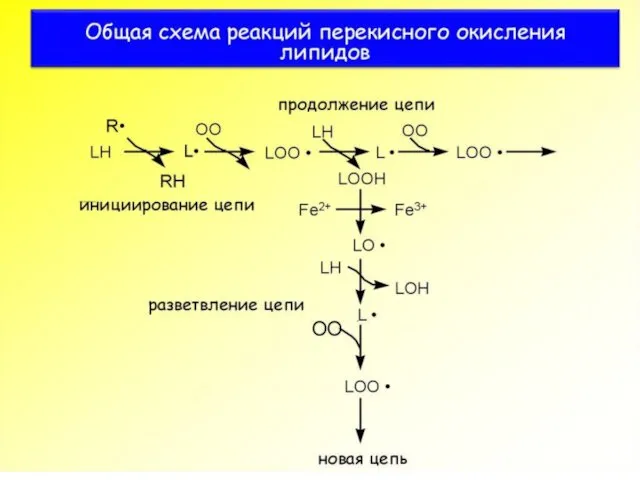
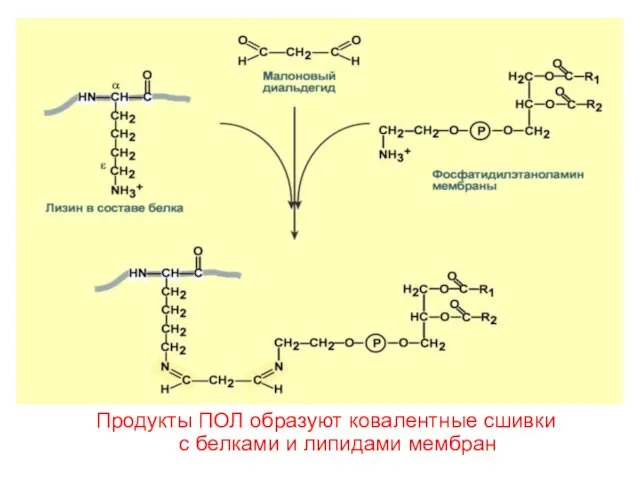
Продукты ПОЛ образуют ковалентные сшивки
с белками и липидами мембран
Продукты ПОЛ образуют ковалентные сшивки
с белками и липидами мембран
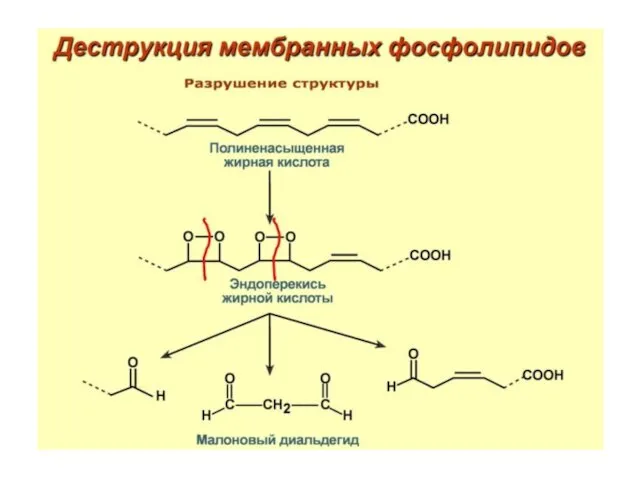
Примеры продуктов ПОЛ
Примеры продуктов ПОЛ
Последовательность синтеза продуктов ПОЛ
Последовательность синтеза продуктов ПОЛ
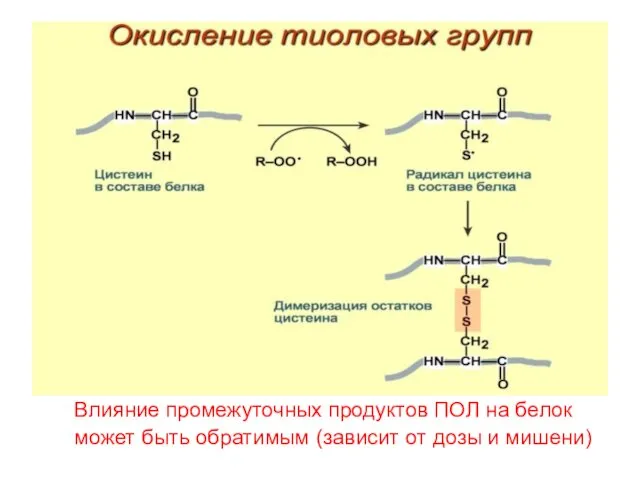
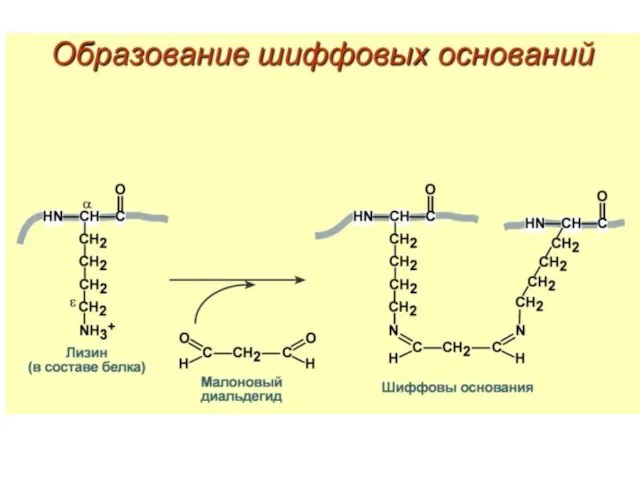
Влияние промежуточных продуктов ПОЛ на белок
может быть обратимым (зависит от дозы
Влияние промежуточных продуктов ПОЛ на белок
может быть обратимым (зависит от дозы
 Laser technologies of Triniti JSC
Laser technologies of Triniti JSC Основные понятия и процессы, определяющие бурение скважины в осложненных и аварийных условиях. Лекция 1
Основные понятия и процессы, определяющие бурение скважины в осложненных и аварийных условиях. Лекция 1 Свет души Презентация для осеннего бала.
Свет души Презентация для осеннего бала. Люди, животные, растения и минералы
Люди, животные, растения и минералы Площадь поверхности
Площадь поверхности Проект Гольф в школу
Проект Гольф в школу Правила дорожного движения
Правила дорожного движения Радіоприймальні пристрої ЗРЛ. Підсилювачі проміжної частоти і детектори. (Тема 4.3)
Радіоприймальні пристрої ЗРЛ. Підсилювачі проміжної частоти і детектори. (Тема 4.3) Глобальные проблемы человечества
Глобальные проблемы человечества Презентация. Поэты и Менделеев.
Презентация. Поэты и Менделеев. Маленький Мук, по сказке В. Гауфа
Маленький Мук, по сказке В. Гауфа Презентация Маленькая планета
Презентация Маленькая планета Рынок труда. Занятость и безработица
Рынок труда. Занятость и безработица Измерительные трансформаторы
Измерительные трансформаторы Подготовка спортсмена в Пауэрлифтинге
Подготовка спортсмена в Пауэрлифтинге Электронное пособие по развитию звукового анализа Мультяшки
Электронное пособие по развитию звукового анализа Мультяшки Хрящевая ткань
Хрящевая ткань Проект Молоко и его польза
Проект Молоко и его польза Лекция+5+Состав+преступления
Лекция+5+Состав+преступления Архитектура с общей шиной
Архитектура с общей шиной Аргументация и доказательство
Аргументация и доказательство Порядок составления заявки на ракетное топливо, горючее и смазочные материалы на год. Расчеты, расшифровки
Порядок составления заявки на ракетное топливо, горючее и смазочные материалы на год. Расчеты, расшифровки Профилактика гриппа и ОРВИ
Профилактика гриппа и ОРВИ Armelle - парфюмерная компания
Armelle - парфюмерная компания Участие во Всероссийской ярмарке и конкурсе событийного туризма
Участие во Всероссийской ярмарке и конкурсе событийного туризма Игра Самый умный
Игра Самый умный Northern Ireland
Northern Ireland Ақсу аудандандық ветеринариялық зертханасы
Ақсу аудандандық ветеринариялық зертханасы