- Межклеточные сигнальные молекулы (первичные посредники)

Содержание
- 2. 1. НЕБОЛЬШИЕ ЛИПОФИЛЬНЫЕ МОЛЕКУЛЫ Первичные посредники
- 3. СХЕМА ДЕЙСТВИЯ СТЕРОИДНЫХ И ТИРЕОИДНЫХ ГОРМОНОВ внутриклеточные рецепторы имеют гормон-связывающий домен ДНК-связывающий домен
- 5. 2. ЛИПОФИЛЬНЫЕ МОЛЕКУЛЫ Производные арахидоновой кислоты – ЭЙКОЗАНОИДЫ: ПРОСТАГЛАНДИНЫ ЛЕЙКОТРИЕНЫ ПРОСТАЦИКЛИНЫ ТРОМБОКСАНЫ Первичные посредники
- 6. ЭЙКОЗАНОИДЫ ОБРАЗУЮТСЯ ПРАКТИЧЕСКИ ВО ВСЕХ КЛЕТКАХ, ИХ БИОСИНТЕЗ ИНИЦИИРУЕТСЯ ФОСФОЛИПАЗОЙ А2 ДЕЙСТВУЮТ ЧЕРЕЗ МЕМБРАННЫЕ РЕЦЕПТОРЫ, ОСУЩЕСТВЛЯЮТ
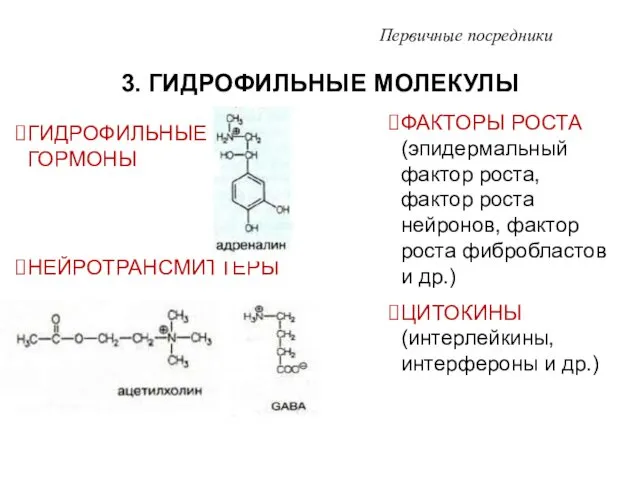
- 7. 3. ГИДРОФИЛЬНЫЕ МОЛЕКУЛЫ Первичные посредники ГИДРОФИЛЬНЫЕ ГОРМОНЫ НЕЙРОТРАНСМИТТЕРЫ ФАКТОРЫ РОСТА (эпидермальный фактор роста, фактор роста нейронов,
- 8. СХЕМА ДЕЙСТВИЯ НЕЙРОМЕДИАТОРОВ Первичные посредники
- 9. СХЕМА ДЕЙСТВИЯ ГИДРОФИЛЬНЫХ АГОНИСТОВ Первичные посредники Гидрофильные гормоны Инсулин, факторы роста
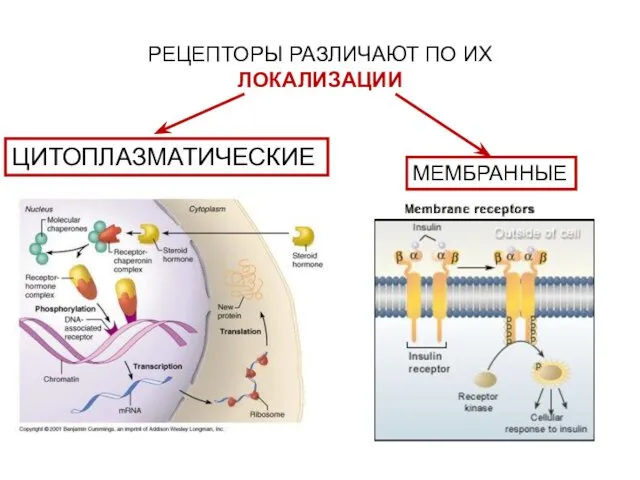
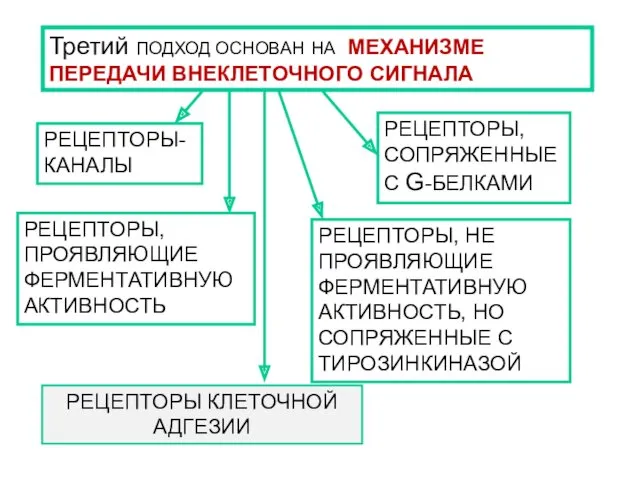
- 10. РЕЦЕПТОРЫ:ОБЩИЕ СВОЙСТВА И КЛАССИФИКАЦИЯ
- 11. РЕЦЕПТОР –СПЕЦИФИЧЕСКИЙ БЕЛОК, КОТОРЫЙ СВЯЗЫВАЕТ СИГНАЛЬНОЕ ВЕЩЕСТВО, ЧТО ПРИВОДИТ В КОНЕЧНОМ ИТОГЕ К РАЗВИТИЮ КЛЕТОЧНОЙ РЕАКЦИИ.
- 12. ХАРАКТЕРИСТИКИ РЕЦЕПТОРОВ
- 13. СЕЛЕКТИВНОСТЬ СВОЙСТВО, ОСНОВАННОЕ НА СТРОГОЙ СТРУКТУРНОЙ СПЕЦИФИЧНОСТИ. ДАННЫЙ ЛИГАНД ДОЛЖЕН БЫТЬ ЛИБО ЕДИНСТВЕННЫМ ВЕЩЕСТВОМ, СВЯЗЫВАЮЩИМСЯ С
- 15. НАСЫЩАЕМОСТЬ ЧИСЛО МЕСТ СВЯЗЫВАНИЯ С ЛИГАНДОМ ДОЛЖНО БЫТЬ КОНЕЧНЫМ.
- 16. СРОДСТВО К ЛИГАНДУ НАСЫЩЕНИЕ РЕЦЕПТОРА ПРОИСХОДИТ ПРИ ФИЗИОЛОГИЧЕСКИХ КОНЦЕНТРАЦИЯХ ЛИГАНДА АКТГ — 0-50 пг/мл Тироксин общий
- 17. ТКАНЕВАЯ СПЕЦИФИЧНОСТЬ СВЯЗЫВАНИЕ ЛИГАНДА С РЕЦЕПТОРОМ ПРОИСХОДИТ В ТОЙ ТКАНИ, ГДЕ НАБЛЮДАЕТСЯ ЕГО БИОЛОГИЧЕСКОЕ ДЕЙСТВИЕ.
- 18. КЛАССИФИКАЦИЯ РЕЦЕПТОРОВ
- 19. Классификация, основанная на видах агонистов, с которым взаимодействует рецептор (применяют, как правило, для мембранных рецепторов). НАПРИМЕР
- 23. ЦИТОПЛАЗМАТИЧЕСКИЕ РЕЦЕПТОРЫ ПРЕДСТАВЛЕНЫ ГЛОБУЛЯРНЫМИ БЕЛКАМИ
- 24. МЕМБРАННЫЕ РЕЦЕПТОРЫ ЯВЛЯЮТСЯ ИНТЕГРАЛЬНЫМИ БЕЛКАМИ ОБЩИЙ ПЛАН СТРОЕНИЯ: ВНЕКЛЕТОЧНЫЙ ДОМЕН СПЕЦИФИЧЕН К ВНЕШНЕМУ СИГНАЛУ ТРАНСМЕМБРАННЫЙ УЧАСТОК
- 25. РЕЦЕПТОРЫ, ПРОЯВЛЯЮЩИЕ ФЕРМЕНТАТИВНУЮ АКТИВНОСТЬ
- 27. РЕЦЕПТОРЫ-КАНАЛЫ
- 28. СВЯЗЫВАНИЕ С ЛИГАНДОМ ПРИВОДИТ К КОНФОРМАЦИОННЫМ ПЕРЕСТРОЙКАМ РЕЦЕПТОРА, ЧТО ВЫЗЫВАЕТ ОТКРЫВАНИЕ ИОННОГО КАНАЛА МЕХАНИЗМ РАБОТЫ РЕЦЕПТОРА-КАНАЛА
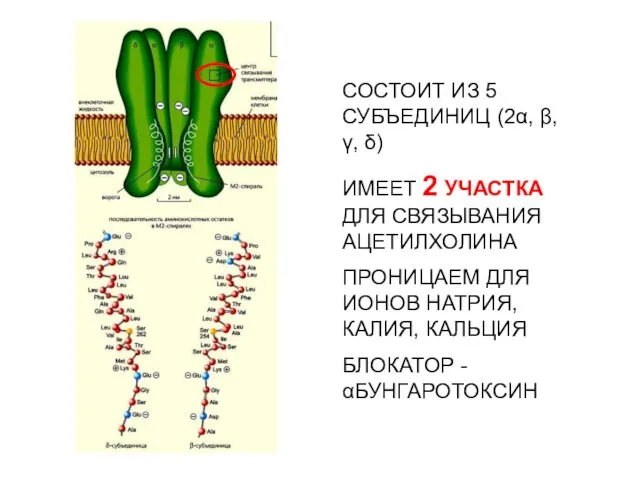
- 29. Электрический скат, из электрического органа которого были выделены и очищены никотиновые холинэргические рецепторы, расшифрована их аминокислотная
- 31. ДРУГИЕ ПРИМЕРЫ: РЕЦЕПТОРЫ К ГЛУТАМАТУ РЕЦЕПТОРЫ К АСПАРТАТУ ГЛИЦИНОВЫЕ РЕЦЕПТОРЫ ГАМК-рецепторы 1,2 проницаемы для катионов 3,4
- 32. РЕЦЕПТОРЫ, СОПРЯЖЕННЫЕ С G-БЕЛКАМИ
- 33. СТРУКТУРА РЕЦЕПТОРОВ,СОПРЯЖЕННЫХ С G-БЕЛКОМ N-концевой домен Участки гликозилирования С-концевой домен
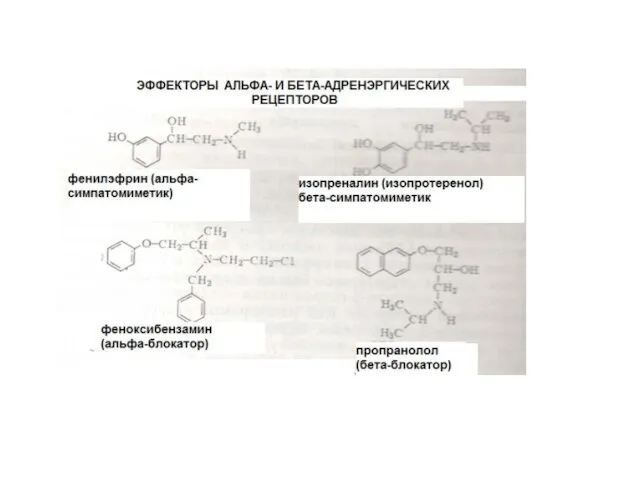
- 34. ПРИМЕРЫ АГОНИСТОВ, ДЕЙСТВУЮЩИХ НА РЕЦЕПТОРЫ, СОПРЯЖЕННЫЕ С G-БЕЛКАМИ АДРЕНАЛИН, НОРАДРЕНАЛИН АЦЕТИЛХОЛИН СЕРОТОНИН ГИСТАМИН ОПИОИДЫ, КАННАБИНОИДЫ ГОРМОНЫ
- 35. ПРИМЕРЫ РЕЦЕПТОРОВ, СОПРЯЖЕННЫХ С G-БЕЛКОМ α1АДРЕНОРЕЦЕПТОР β2 АДРЕНОРЕЦЕПТОР
- 36. Кроме ферментов, G-белки могут взаимодействовать с ион-транспортными системами, например, каналами, ионообменниками.
- 37. Калиевый канал
- 38. СТРУКТУРА G-БЕЛКА
- 39. МЕХАНИЗМ ФУНКЦИОНИРОВАНИЯ РЕЦЕПТОРОВ, СОПРЯЖЕННЫХ С G-БЕЛКОМ
- 41. ВИДЫ G-БЕЛКОВ ИДЕНТИФИЦИРОВАНО ОКОЛО 20 ВИДОВ G-БЕЛКОВ GS И GI БЕЛКИ ДЕЙСТВУЮТ НА АДЕНИЛАТЦИКЛАЗУ GO ИНГИБИРУЕТ
- 42. ИСПОЛЬЗОВАНИЕ БАКТЕРИАЛЬНЫХ ТОКСИНОВ ПОЗВОЛИЛО ОБНАРУЖИТЬ ДВА ВИДА G-БЕЛКОВ ПО ОТНОШЕНИЮ К АДЕНИЛАТЦИКЛАЗЕ стимулирующий ингибирующий Холерный токсин
- 44. Холерный токсин, также как и коклюшный, содержит фермент АДФ-рибозилазу. АДФрибозилирование α-субъединицы GS белка приводит к ее
- 45. ЦИКЛИЧЕСКИЕ НУКЛЕОТИДЫ В РОЛИ ВТОРИЧНЫХ ПОСРЕДНИКОВ
- 46. цАМФ открыт при изучении стимуляции гликогенолиза в печени адреналином. Циклический аденозинмонофосфат (цАМФ) – первое соединение, которое
- 47. Фермент, который образует цАМФ, - аденилатциклаза Фермент, который разрушает цАМФ (переводит в нециклическую форму) – фосфодиэстераза
- 48. ОБРАЗОВАНИЕ И РАЗРУШЕНИЕ цАМФ Внутриклеточная концентрация цАМФ определяется скоростью синтеза и распада этого соединения
- 49. АТФ АМФ цАМФ АЦ ФДЭ РЕСИНТЕЗ АТФ
- 50. МЕХАНИЗМ РЕГУЛЯЦИИ АДЕНИЛАТЦИКЛАЗЫ β-адренорецепторы α2 адренорецептор
- 51. ХАРАКТЕРИСТИКИ цАМФ КАК ВТОРИЧНОГО ПОСРЕДНИКА Лиганд должен стимулировать аденилатциклазу в препаратах чувствительной ткани В ответ на
- 52. ПРИМЕРЫ цАМФ-ЗАВИСИМЫХ ПРОЦЕССОВ
- 53. ИЗМЕНЕНИЕ ПРОНИЦАЕМОСТИ КЛЕТОЧНЫХ МЕМБРАН СИНТЕЗ СТЕРОИДНЫХ ГОРМОНОВ СЕКРЕЦИЯ ЭКЗО- И ЭНДОКРИННЫХ ЖЕЛЕЗ ПЕРЕМЕЩЕНИЯ ВНУТРИКЛЕТОЧНЫХ СТРУКТУР цАМФ-зависимые
- 54. МЕХАНИЗМ КАСКАДНОГО УСИЛЕНИЯ РЕГУЛЯТОРНОГО СИГНАЛА С УЧАСТИЕМ цАМФ
- 55. ПРОТЕИНКИНАЗА А Протеинкиназа А в неактивном состоянии состоит из 2 регуляторных и 2 каталитических субъединиц. После
- 57. БЕЛКИ-СУБСТРАТЫ ДЛЯ ПРОТЕИНКИНАЗЫ А БЕЛКИ ИОННЫХ КАНАЛОВ ИОНООБМЕННИКИ ТРАНСПОРТНЫЕ АТФазы БЕЛКИ ЦИТОСКЕЛЕТА ФЕРМЕНТЫ, РЕГУЛИРУЮЩМЕ МЕТАБОЛИЧЕСКИЕ ПРОЦЕССЫ
- 58. цГМФ ОКАЗАЛОСЬ, ЧТО И цГМФ ЯВЛЯЕТСЯ ВТОРИЧНЫМ ПОСРЕДНИКОМ
- 59. ОБРАЗОВАНИЕ И РАСПАД цГМФ ФЕРМЕНТ, КОТОРЫЙ ОБРАЗУЕТ цГМФ, ЭТО ГУАНИЛАТЦИКЛАЗА ФЕРМЕНТ, КОТОРЫЙ ЕГО РАЗРУШАЕТ - ФОСФОДИЭСТЕРАЗА
- 60. цГМФ играет важную роль в регуляции Са2+-гомеостаза в различных типах клеток. Повышение концентрации цГМФ приводит к
- 61. ПК G фосфорилирует Na,K-АТФазу эпителиоцитов почечных канальцев, что приводит к торможению насоса. Результат: ослабление реабсорбции ионов
- 62. ПУТИ ОБРАЗОВАНИЯ цГМФ ОПОСРЕДОВАННЫЙ БАВ Например, натрийуретическим пептидом ОПОСРЕДОВАННЫЙ ОКСИДОМ АЗОТА (NO)
- 63. ДВЕ ФОРМЫ ГУАНИЛАТЦИКЛАЗЫ Мембранная гуанилатциклаза Растворимая форма гуанилатциклазы активируется оксидом азота ПРОТЕИНКИНАЗА G
- 64. МЕХАНИЗМ ДЕЙСТВИЯ НАТРИЙУРЕТИЧЕСКОГО ПЕПТИДА NaУП + R → ↑ГЦ → ↑цГМФ → ↑ПК G
- 65. ОСОБЕННОСТИ цГМФ-ЗАВИСИМОЙ РЕГУЛЯЦИИ ОТСУТСТВУЮТ G-БЕЛКИ СВЯЗЫВАНИЕ АГОНИСТА С РЕЦЕПТОРОМ ВСЕГДА ПРИВОДИТ К АКТИВАЦИИ ГУАНИЛАТЦИКЛАЗЫ
- 66. РАСТВОРИМАЯ ФОРМА ГЦ ГЕТЕРОДИМЕР СОДЕРЖИТ SH-группы СОДЕРЖИТ ГЕМ АКТИВИРУЕТСЯ ОКСИДОМ АЗОТА ШИРОКО РАСПРОСТРАНЕНА
- 68. Скачать презентацию
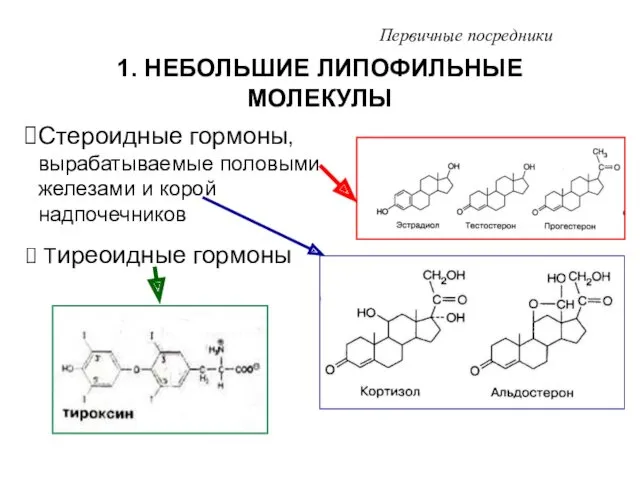
1. НЕБОЛЬШИЕ ЛИПОФИЛЬНЫЕ МОЛЕКУЛЫ
Первичные посредники
1. НЕБОЛЬШИЕ ЛИПОФИЛЬНЫЕ МОЛЕКУЛЫ
Первичные посредники
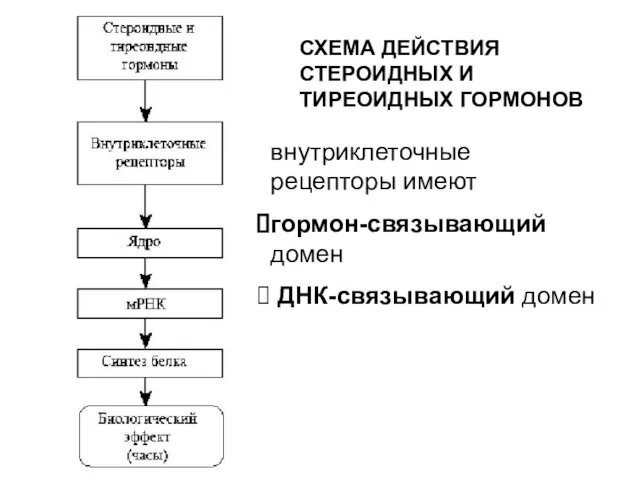
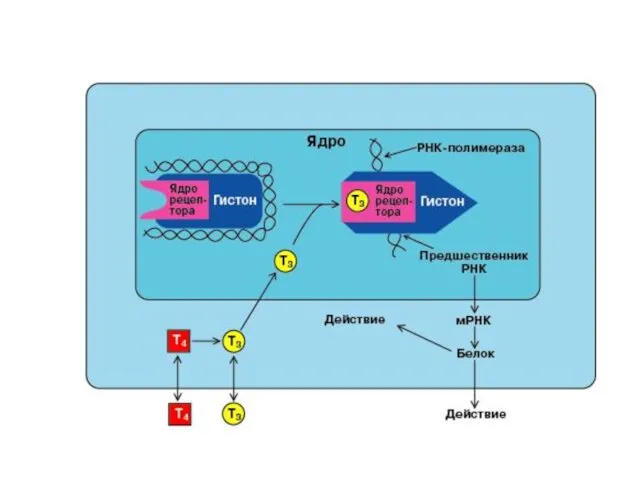
СХЕМА ДЕЙСТВИЯ СТЕРОИДНЫХ И ТИРЕОИДНЫХ ГОРМОНОВ
внутриклеточные рецепторы имеют
гормон-связывающий домен
СХЕМА ДЕЙСТВИЯ СТЕРОИДНЫХ И ТИРЕОИДНЫХ ГОРМОНОВ
внутриклеточные рецепторы имеют
гормон-связывающий домен
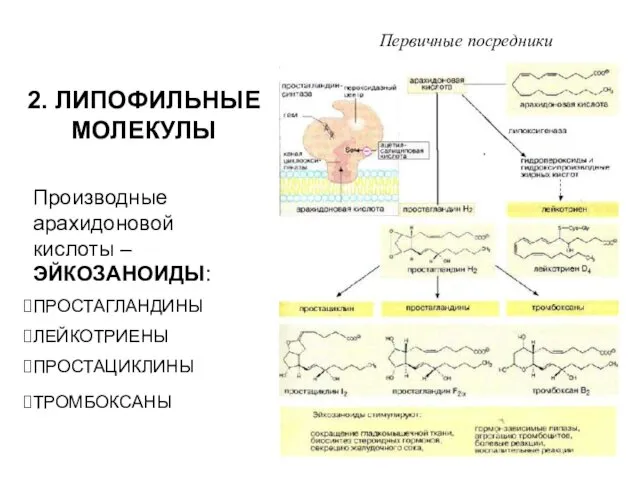
2. ЛИПОФИЛЬНЫЕ МОЛЕКУЛЫ
Производные арахидоновой кислоты – ЭЙКОЗАНОИДЫ:
ПРОСТАГЛАНДИНЫ
ЛЕЙКОТРИЕНЫ
ПРОСТАЦИКЛИНЫ
ТРОМБОКСАНЫ
Первичные посредники
2. ЛИПОФИЛЬНЫЕ МОЛЕКУЛЫ
Производные арахидоновой кислоты – ЭЙКОЗАНОИДЫ:
ПРОСТАГЛАНДИНЫ
ЛЕЙКОТРИЕНЫ
ПРОСТАЦИКЛИНЫ
ТРОМБОКСАНЫ
Первичные посредники

ЭЙКОЗАНОИДЫ ОБРАЗУЮТСЯ ПРАКТИЧЕСКИ ВО ВСЕХ КЛЕТКАХ, ИХ БИОСИНТЕЗ ИНИЦИИРУЕТСЯ ФОСФОЛИПАЗОЙ А2
ЭЙКОЗАНОИДЫ ОБРАЗУЮТСЯ ПРАКТИЧЕСКИ ВО ВСЕХ КЛЕТКАХ, ИХ БИОСИНТЕЗ ИНИЦИИРУЕТСЯ ФОСФОЛИПАЗОЙ А2
3. ГИДРОФИЛЬНЫЕ МОЛЕКУЛЫ
Первичные посредники
ГИДРОФИЛЬНЫЕ ГОРМОНЫ
НЕЙРОТРАНСМИТТЕРЫ
ФАКТОРЫ РОСТА (эпидермальный фактор роста, фактор роста
3. ГИДРОФИЛЬНЫЕ МОЛЕКУЛЫ
Первичные посредники
ГИДРОФИЛЬНЫЕ ГОРМОНЫ
НЕЙРОТРАНСМИТТЕРЫ
ФАКТОРЫ РОСТА (эпидермальный фактор роста, фактор роста
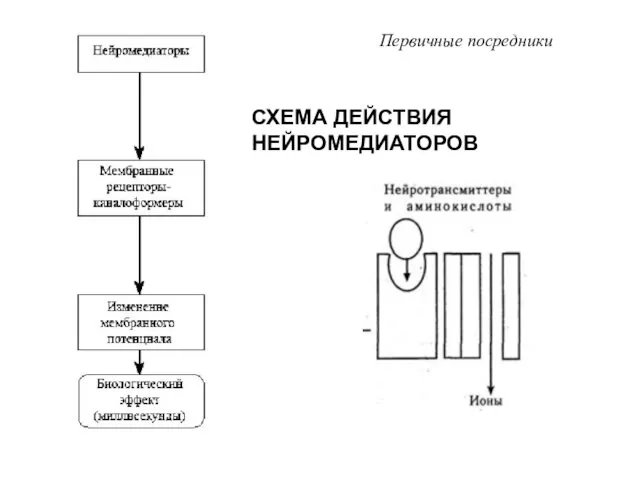
СХЕМА ДЕЙСТВИЯ НЕЙРОМЕДИАТОРОВ
Первичные посредники
СХЕМА ДЕЙСТВИЯ НЕЙРОМЕДИАТОРОВ
Первичные посредники
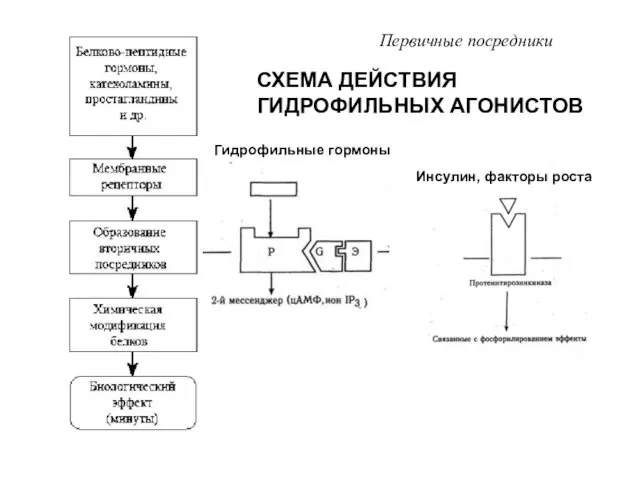
СХЕМА ДЕЙСТВИЯ ГИДРОФИЛЬНЫХ АГОНИСТОВ
Первичные посредники
Гидрофильные гормоны
Инсулин, факторы роста
СХЕМА ДЕЙСТВИЯ ГИДРОФИЛЬНЫХ АГОНИСТОВ
Первичные посредники
Гидрофильные гормоны
Инсулин, факторы роста

РЕЦЕПТОРЫ:ОБЩИЕ СВОЙСТВА И КЛАССИФИКАЦИЯ
РЕЦЕПТОРЫ:ОБЩИЕ СВОЙСТВА И КЛАССИФИКАЦИЯ

РЕЦЕПТОР –СПЕЦИФИЧЕСКИЙ БЕЛОК, КОТОРЫЙ СВЯЗЫВАЕТ СИГНАЛЬНОЕ ВЕЩЕСТВО, ЧТО ПРИВОДИТ В КОНЕЧНОМ
РЕЦЕПТОР –СПЕЦИФИЧЕСКИЙ БЕЛОК, КОТОРЫЙ СВЯЗЫВАЕТ СИГНАЛЬНОЕ ВЕЩЕСТВО, ЧТО ПРИВОДИТ В КОНЕЧНОМ

ХАРАКТЕРИСТИКИ РЕЦЕПТОРОВ
ХАРАКТЕРИСТИКИ РЕЦЕПТОРОВ
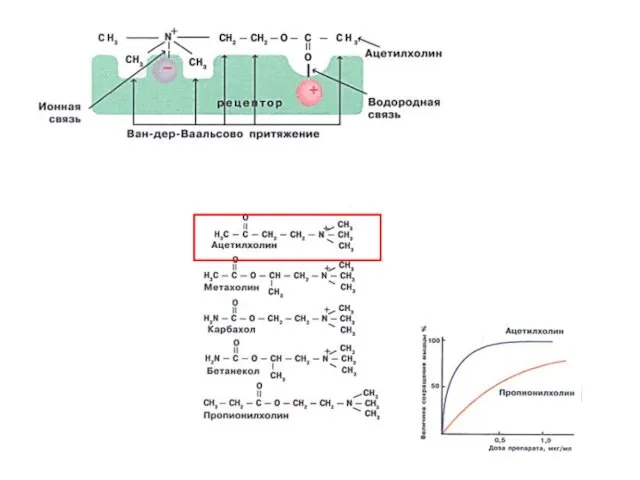
СЕЛЕКТИВНОСТЬ
СВОЙСТВО, ОСНОВАННОЕ НА СТРОГОЙ СТРУКТУРНОЙ СПЕЦИФИЧНОСТИ. ДАННЫЙ ЛИГАНД ДОЛЖЕН БЫТЬ
СЕЛЕКТИВНОСТЬ
СВОЙСТВО, ОСНОВАННОЕ НА СТРОГОЙ СТРУКТУРНОЙ СПЕЦИФИЧНОСТИ. ДАННЫЙ ЛИГАНД ДОЛЖЕН БЫТЬ
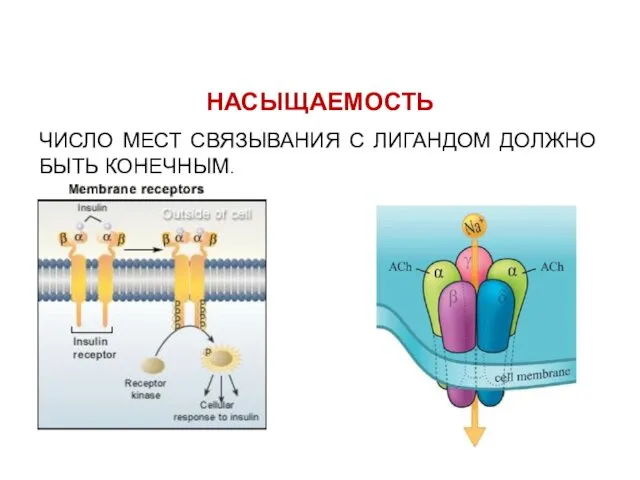
НАСЫЩАЕМОСТЬ
ЧИСЛО МЕСТ СВЯЗЫВАНИЯ С ЛИГАНДОМ ДОЛЖНО БЫТЬ КОНЕЧНЫМ.
НАСЫЩАЕМОСТЬ
ЧИСЛО МЕСТ СВЯЗЫВАНИЯ С ЛИГАНДОМ ДОЛЖНО БЫТЬ КОНЕЧНЫМ.

СРОДСТВО К ЛИГАНДУ
НАСЫЩЕНИЕ РЕЦЕПТОРА ПРОИСХОДИТ ПРИ ФИЗИОЛОГИЧЕСКИХ КОНЦЕНТРАЦИЯХ ЛИГАНДА
АКТГ —
СРОДСТВО К ЛИГАНДУ
НАСЫЩЕНИЕ РЕЦЕПТОРА ПРОИСХОДИТ ПРИ ФИЗИОЛОГИЧЕСКИХ КОНЦЕНТРАЦИЯХ ЛИГАНДА
АКТГ —
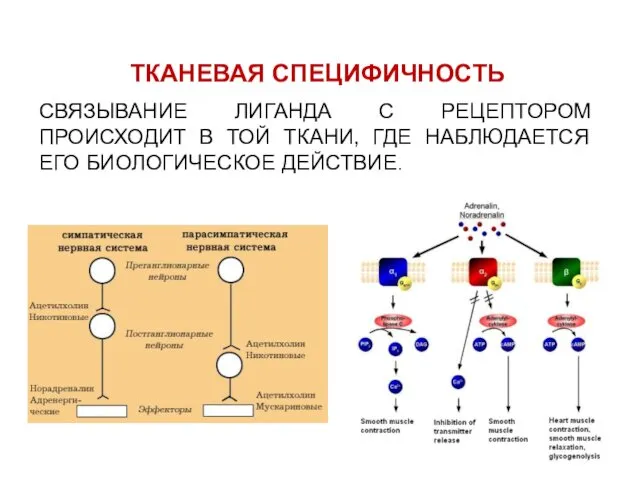
ТКАНЕВАЯ СПЕЦИФИЧНОСТЬ
СВЯЗЫВАНИЕ ЛИГАНДА С РЕЦЕПТОРОМ ПРОИСХОДИТ В ТОЙ ТКАНИ, ГДЕ НАБЛЮДАЕТСЯ
ТКАНЕВАЯ СПЕЦИФИЧНОСТЬ
СВЯЗЫВАНИЕ ЛИГАНДА С РЕЦЕПТОРОМ ПРОИСХОДИТ В ТОЙ ТКАНИ, ГДЕ НАБЛЮДАЕТСЯ

КЛАССИФИКАЦИЯ РЕЦЕПТОРОВ
КЛАССИФИКАЦИЯ РЕЦЕПТОРОВ

Классификация, основанная на видах агонистов, с которым взаимодействует рецептор (применяют, как
Классификация, основанная на видах агонистов, с которым взаимодействует рецептор (применяют, как
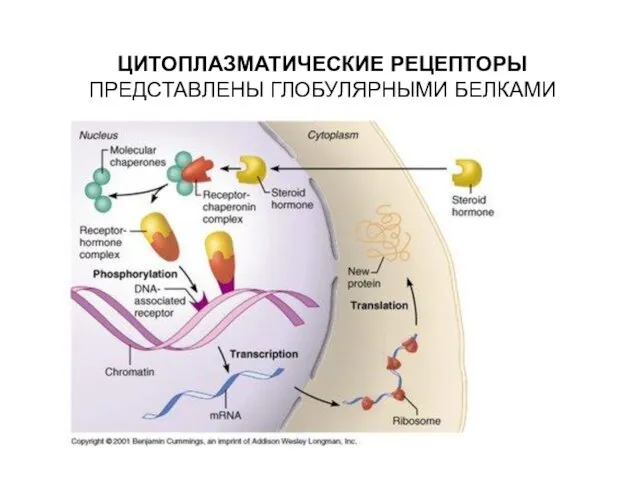
ЦИТОПЛАЗМАТИЧЕСКИЕ РЕЦЕПТОРЫ ПРЕДСТАВЛЕНЫ ГЛОБУЛЯРНЫМИ БЕЛКАМИ
ЦИТОПЛАЗМАТИЧЕСКИЕ РЕЦЕПТОРЫ ПРЕДСТАВЛЕНЫ ГЛОБУЛЯРНЫМИ БЕЛКАМИ
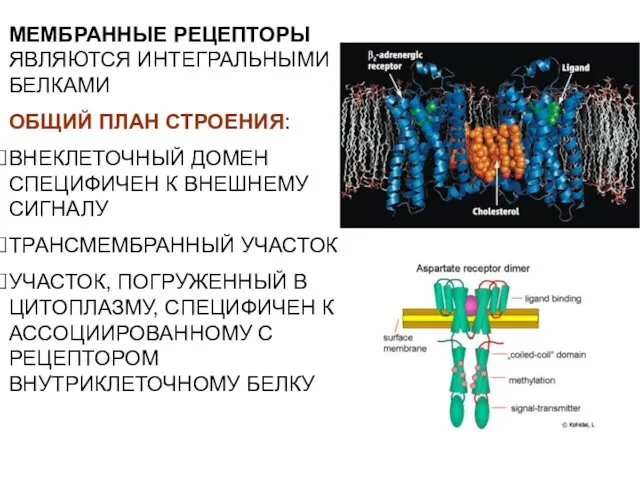
МЕМБРАННЫЕ РЕЦЕПТОРЫ ЯВЛЯЮТСЯ ИНТЕГРАЛЬНЫМИ БЕЛКАМИ
ОБЩИЙ ПЛАН СТРОЕНИЯ:
ВНЕКЛЕТОЧНЫЙ ДОМЕН СПЕЦИФИЧЕН К
МЕМБРАННЫЕ РЕЦЕПТОРЫ ЯВЛЯЮТСЯ ИНТЕГРАЛЬНЫМИ БЕЛКАМИ
ОБЩИЙ ПЛАН СТРОЕНИЯ:
ВНЕКЛЕТОЧНЫЙ ДОМЕН СПЕЦИФИЧЕН К
РЕЦЕПТОРЫ, ПРОЯВЛЯЮЩИЕ ФЕРМЕНТАТИВНУЮ АКТИВНОСТЬ
РЕЦЕПТОРЫ, ПРОЯВЛЯЮЩИЕ ФЕРМЕНТАТИВНУЮ АКТИВНОСТЬ
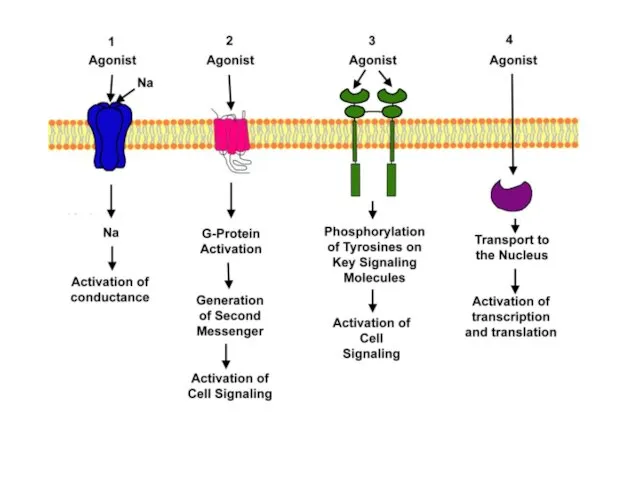

РЕЦЕПТОРЫ-КАНАЛЫ
РЕЦЕПТОРЫ-КАНАЛЫ
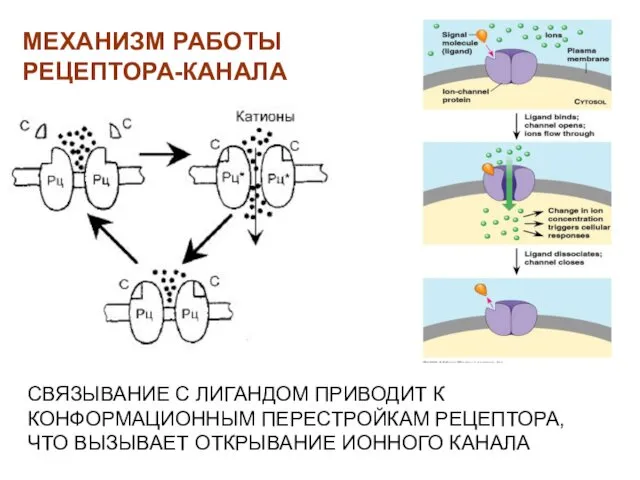
СВЯЗЫВАНИЕ С ЛИГАНДОМ ПРИВОДИТ К КОНФОРМАЦИОННЫМ ПЕРЕСТРОЙКАМ РЕЦЕПТОРА, ЧТО ВЫЗЫВАЕТ ОТКРЫВАНИЕ
СВЯЗЫВАНИЕ С ЛИГАНДОМ ПРИВОДИТ К КОНФОРМАЦИОННЫМ ПЕРЕСТРОЙКАМ РЕЦЕПТОРА, ЧТО ВЫЗЫВАЕТ ОТКРЫВАНИЕ

Электрический скат, из электрического органа которого были выделены и очищены никотиновые
Электрический скат, из электрического органа которого были выделены и очищены никотиновые
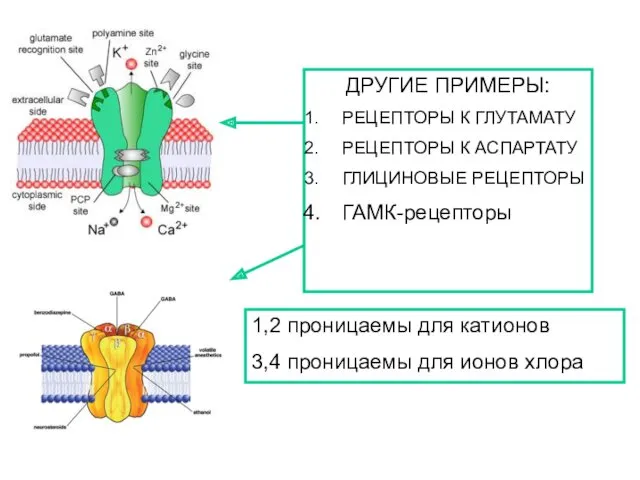
ДРУГИЕ ПРИМЕРЫ:
РЕЦЕПТОРЫ К ГЛУТАМАТУ
РЕЦЕПТОРЫ К АСПАРТАТУ
ГЛИЦИНОВЫЕ РЕЦЕПТОРЫ
ГАМК-рецепторы
1,2 проницаемы для катионов
3,4 проницаемы
ДРУГИЕ ПРИМЕРЫ:
РЕЦЕПТОРЫ К ГЛУТАМАТУ
РЕЦЕПТОРЫ К АСПАРТАТУ
ГЛИЦИНОВЫЕ РЕЦЕПТОРЫ
ГАМК-рецепторы
1,2 проницаемы для катионов
3,4 проницаемы

РЕЦЕПТОРЫ, СОПРЯЖЕННЫЕ С G-БЕЛКАМИ
РЕЦЕПТОРЫ, СОПРЯЖЕННЫЕ С G-БЕЛКАМИ
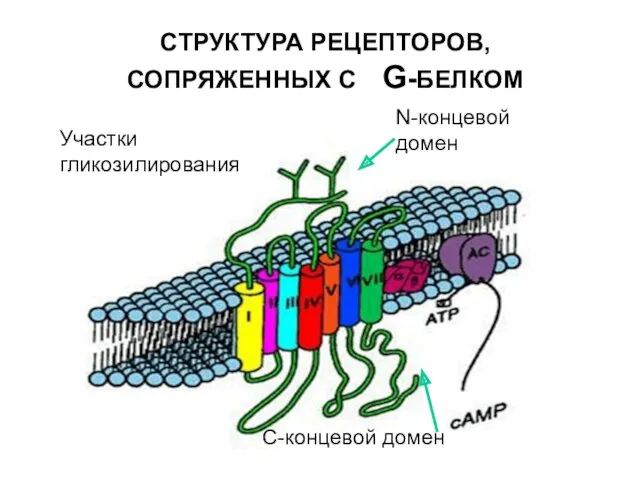
СТРУКТУРА РЕЦЕПТОРОВ,СОПРЯЖЕННЫХ С G-БЕЛКОМ
N-концевой домен
Участки гликозилирования
С-концевой домен
СТРУКТУРА РЕЦЕПТОРОВ,СОПРЯЖЕННЫХ С G-БЕЛКОМ
N-концевой домен
Участки гликозилирования
С-концевой домен

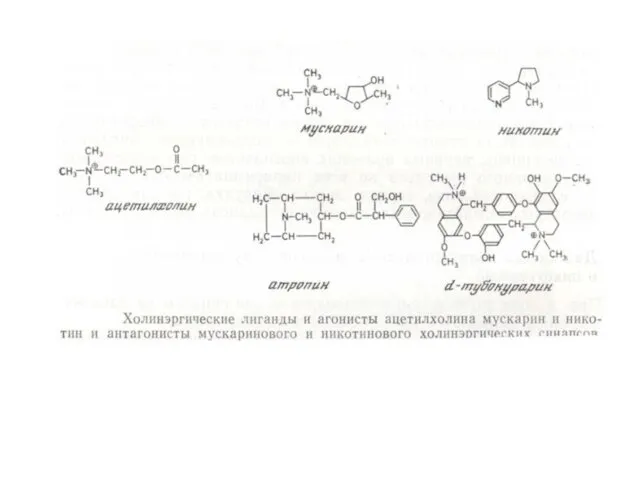
ПРИМЕРЫ АГОНИСТОВ, ДЕЙСТВУЮЩИХ НА РЕЦЕПТОРЫ, СОПРЯЖЕННЫЕ С G-БЕЛКАМИ
АДРЕНАЛИН, НОРАДРЕНАЛИН
АЦЕТИЛХОЛИН
СЕРОТОНИН
ГИСТАМИН
ОПИОИДЫ, КАННАБИНОИДЫ
ГОРМОНЫ ПЕПТИДНОЙ
ПРИМЕРЫ АГОНИСТОВ, ДЕЙСТВУЮЩИХ НА РЕЦЕПТОРЫ, СОПРЯЖЕННЫЕ С G-БЕЛКАМИ
АДРЕНАЛИН, НОРАДРЕНАЛИН
АЦЕТИЛХОЛИН
СЕРОТОНИН
ГИСТАМИН
ОПИОИДЫ, КАННАБИНОИДЫ
ГОРМОНЫ ПЕПТИДНОЙ
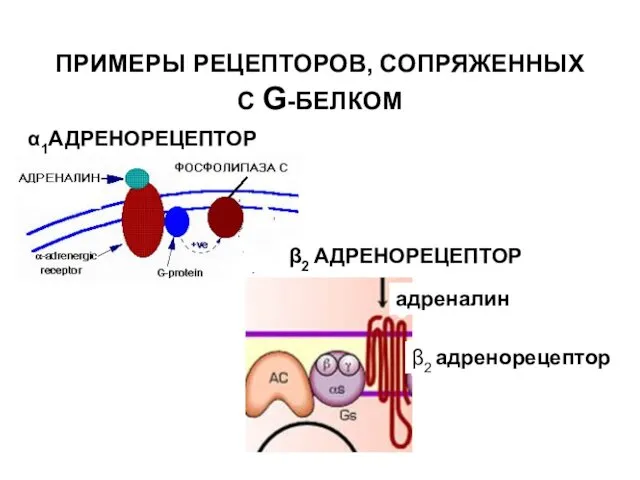
ПРИМЕРЫ РЕЦЕПТОРОВ, СОПРЯЖЕННЫХ С G-БЕЛКОМ
α1АДРЕНОРЕЦЕПТОР
β2 АДРЕНОРЕЦЕПТОР
ПРИМЕРЫ РЕЦЕПТОРОВ, СОПРЯЖЕННЫХ С G-БЕЛКОМ
α1АДРЕНОРЕЦЕПТОР
β2 АДРЕНОРЕЦЕПТОР
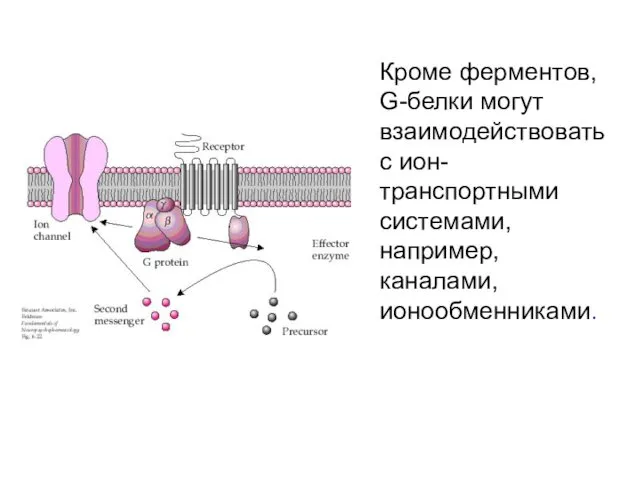
Кроме ферментов, G-белки могут взаимодействовать с ион-транспортными системами, например, каналами, ионообменниками.
Кроме ферментов, G-белки могут взаимодействовать с ион-транспортными системами, например, каналами, ионообменниками.
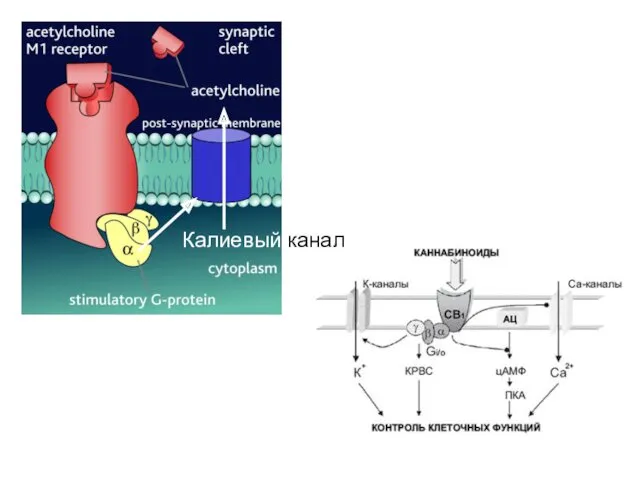
Калиевый канал
Калиевый канал
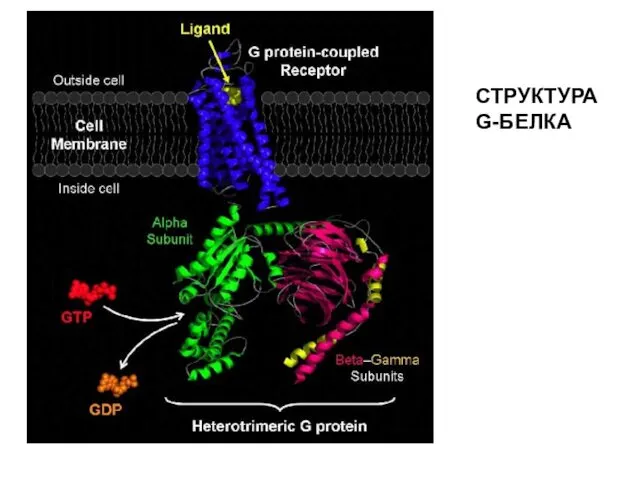
СТРУКТУРА G-БЕЛКА
СТРУКТУРА G-БЕЛКА
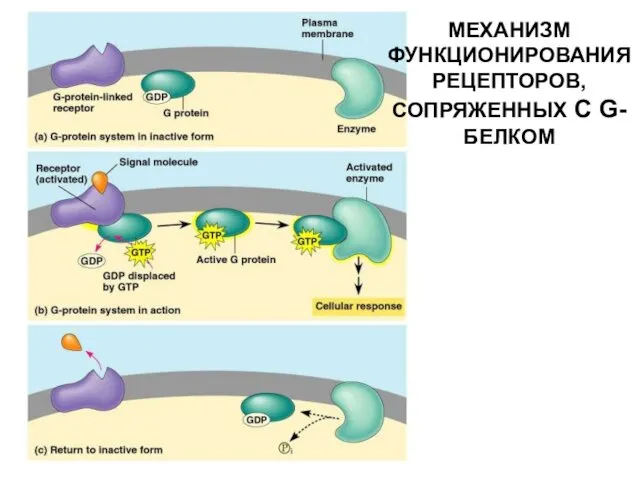
МЕХАНИЗМ ФУНКЦИОНИРОВАНИЯ РЕЦЕПТОРОВ, СОПРЯЖЕННЫХ С G-БЕЛКОМ
МЕХАНИЗМ ФУНКЦИОНИРОВАНИЯ РЕЦЕПТОРОВ, СОПРЯЖЕННЫХ С G-БЕЛКОМ

ВИДЫ G-БЕЛКОВ
ИДЕНТИФИЦИРОВАНО ОКОЛО 20 ВИДОВ G-БЕЛКОВ
GS И GI БЕЛКИ ДЕЙСТВУЮТ НА
ВИДЫ G-БЕЛКОВ
ИДЕНТИФИЦИРОВАНО ОКОЛО 20 ВИДОВ G-БЕЛКОВ
GS И GI БЕЛКИ ДЕЙСТВУЮТ НА
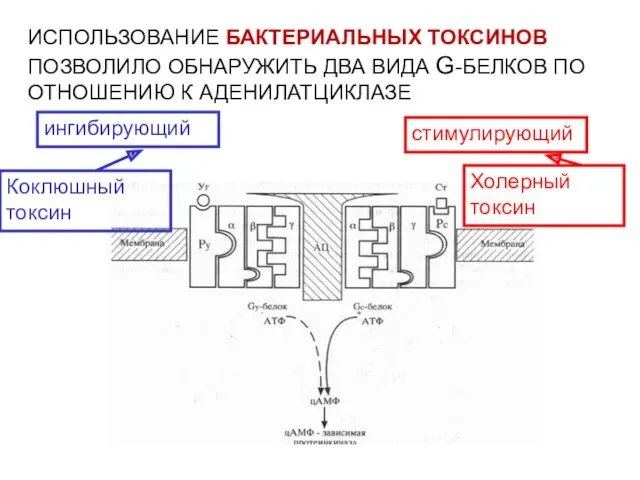
ИСПОЛЬЗОВАНИЕ БАКТЕРИАЛЬНЫХ ТОКСИНОВ ПОЗВОЛИЛО ОБНАРУЖИТЬ ДВА ВИДА G-БЕЛКОВ ПО ОТНОШЕНИЮ К
ИСПОЛЬЗОВАНИЕ БАКТЕРИАЛЬНЫХ ТОКСИНОВ ПОЗВОЛИЛО ОБНАРУЖИТЬ ДВА ВИДА G-БЕЛКОВ ПО ОТНОШЕНИЮ К
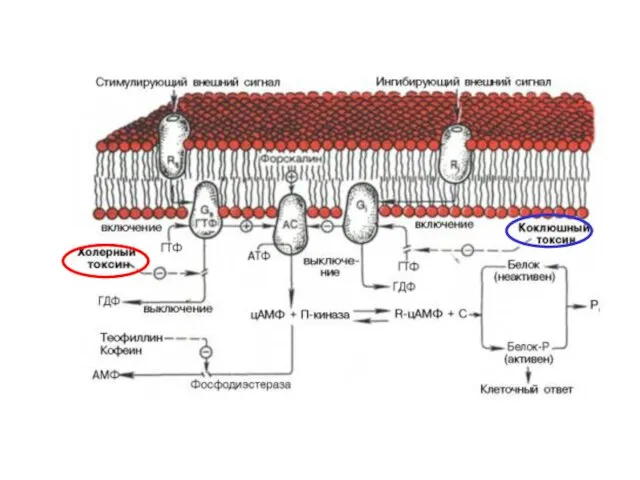

Холерный токсин, также как и коклюшный, содержит фермент АДФ-рибозилазу.
АДФрибозилирование α-субъединицы
Холерный токсин, также как и коклюшный, содержит фермент АДФ-рибозилазу.
АДФрибозилирование α-субъединицы

ЦИКЛИЧЕСКИЕ НУКЛЕОТИДЫ В РОЛИ ВТОРИЧНЫХ ПОСРЕДНИКОВ
ЦИКЛИЧЕСКИЕ НУКЛЕОТИДЫ В РОЛИ ВТОРИЧНЫХ ПОСРЕДНИКОВ
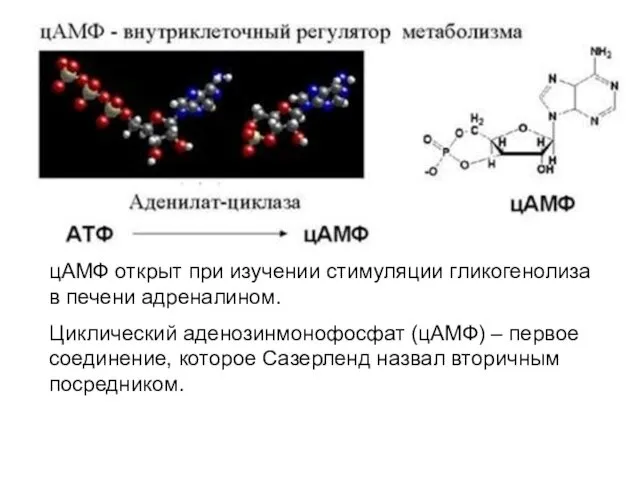
цАМФ открыт при изучении стимуляции гликогенолиза в печени адреналином.
Циклический аденозинмонофосфат
цАМФ открыт при изучении стимуляции гликогенолиза в печени адреналином.
Циклический аденозинмонофосфат
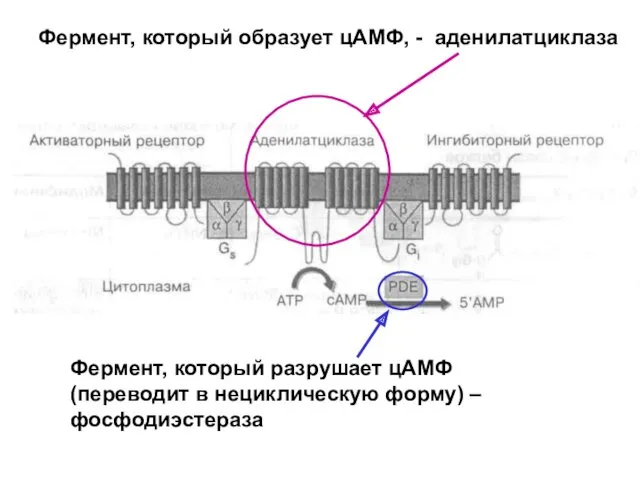
Фермент, который образует цАМФ, - аденилатциклаза
Фермент, который разрушает цАМФ (переводит
Фермент, который образует цАМФ, - аденилатциклаза
Фермент, который разрушает цАМФ (переводит
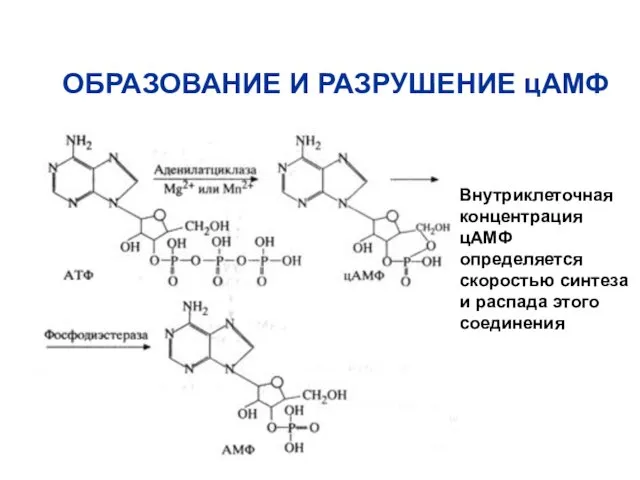
ОБРАЗОВАНИЕ И РАЗРУШЕНИЕ цАМФ
Внутриклеточная концентрация цАМФ определяется скоростью синтеза и распада
ОБРАЗОВАНИЕ И РАЗРУШЕНИЕ цАМФ
Внутриклеточная концентрация цАМФ определяется скоростью синтеза и распада
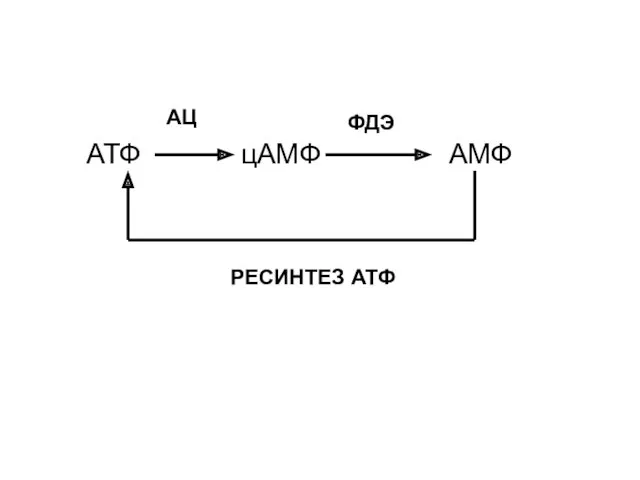
АТФ
АМФ
цАМФ
АЦ
ФДЭ
РЕСИНТЕЗ АТФ
АТФ
АМФ
цАМФ
АЦ
ФДЭ
РЕСИНТЕЗ АТФ
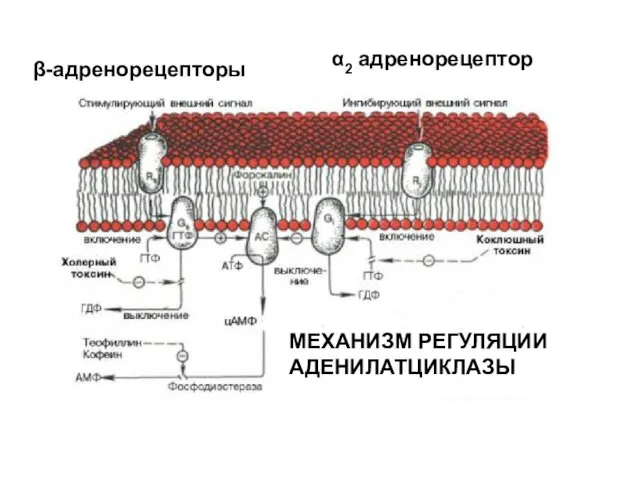
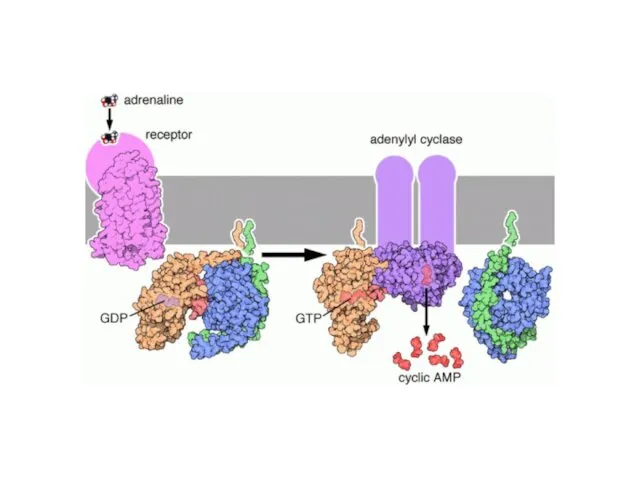
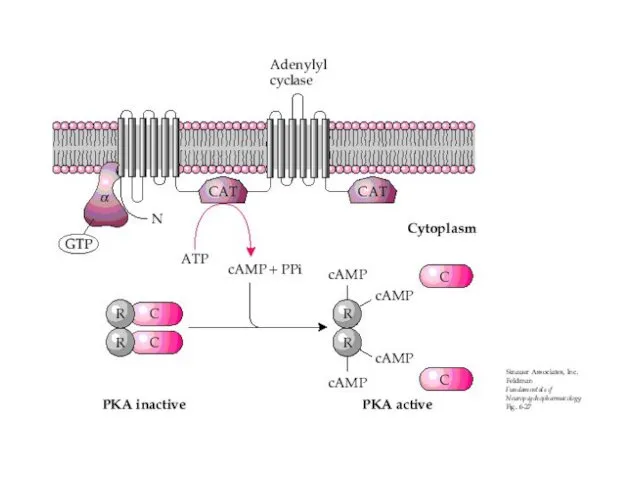
МЕХАНИЗМ РЕГУЛЯЦИИ АДЕНИЛАТЦИКЛАЗЫ
β-адренорецепторы
α2 адренорецептор
МЕХАНИЗМ РЕГУЛЯЦИИ АДЕНИЛАТЦИКЛАЗЫ
β-адренорецепторы
α2 адренорецептор
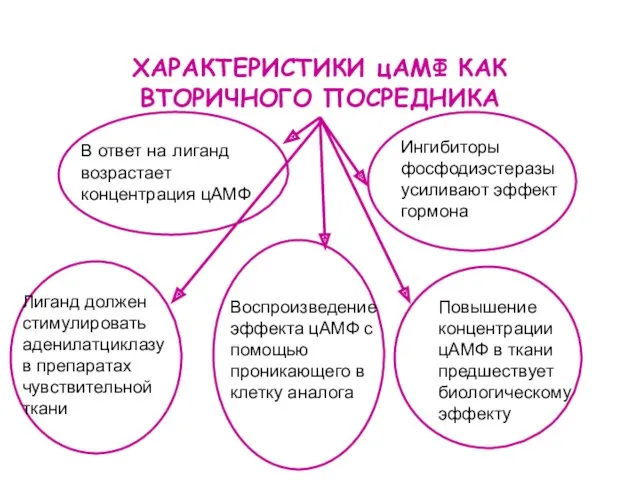
ХАРАКТЕРИСТИКИ цАМФ КАК ВТОРИЧНОГО ПОСРЕДНИКА
Лиганд должен стимулировать аденилатциклазу в препаратах чувствительной
ХАРАКТЕРИСТИКИ цАМФ КАК ВТОРИЧНОГО ПОСРЕДНИКА
Лиганд должен стимулировать аденилатциклазу в препаратах чувствительной

ПРИМЕРЫ цАМФ-ЗАВИСИМЫХ ПРОЦЕССОВ
ПРИМЕРЫ цАМФ-ЗАВИСИМЫХ ПРОЦЕССОВ
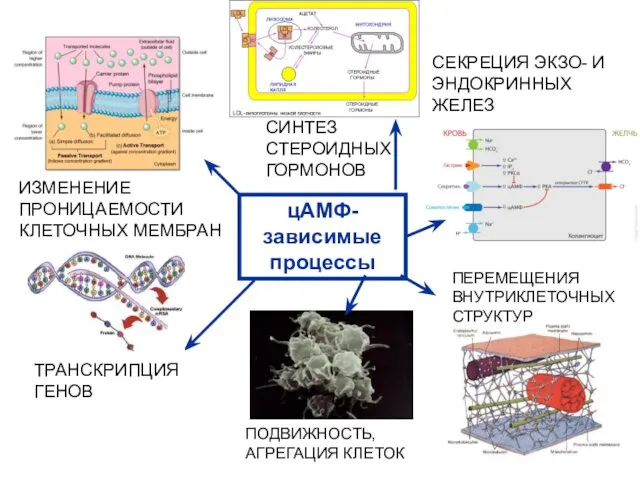
ИЗМЕНЕНИЕ ПРОНИЦАЕМОСТИ КЛЕТОЧНЫХ МЕМБРАН
СИНТЕЗ СТЕРОИДНЫХ ГОРМОНОВ
СЕКРЕЦИЯ ЭКЗО- И ЭНДОКРИННЫХ ЖЕЛЕЗ
ПЕРЕМЕЩЕНИЯ ВНУТРИКЛЕТОЧНЫХ
ИЗМЕНЕНИЕ ПРОНИЦАЕМОСТИ КЛЕТОЧНЫХ МЕМБРАН
СИНТЕЗ СТЕРОИДНЫХ ГОРМОНОВ
СЕКРЕЦИЯ ЭКЗО- И ЭНДОКРИННЫХ ЖЕЛЕЗ
ПЕРЕМЕЩЕНИЯ ВНУТРИКЛЕТОЧНЫХ
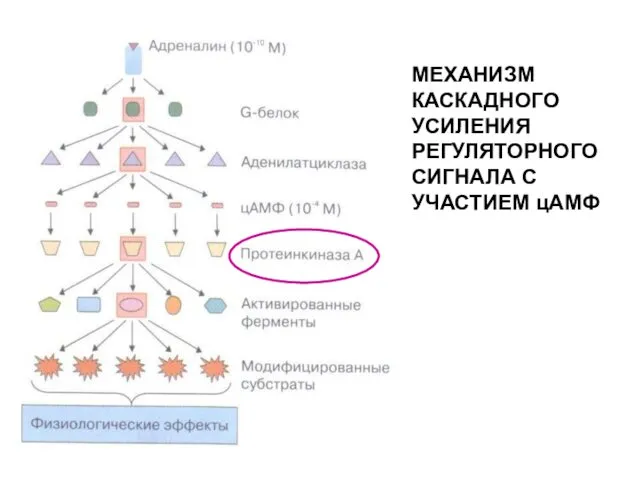
МЕХАНИЗМ КАСКАДНОГО УСИЛЕНИЯ РЕГУЛЯТОРНОГО СИГНАЛА С УЧАСТИЕМ цАМФ
МЕХАНИЗМ КАСКАДНОГО УСИЛЕНИЯ РЕГУЛЯТОРНОГО СИГНАЛА С УЧАСТИЕМ цАМФ
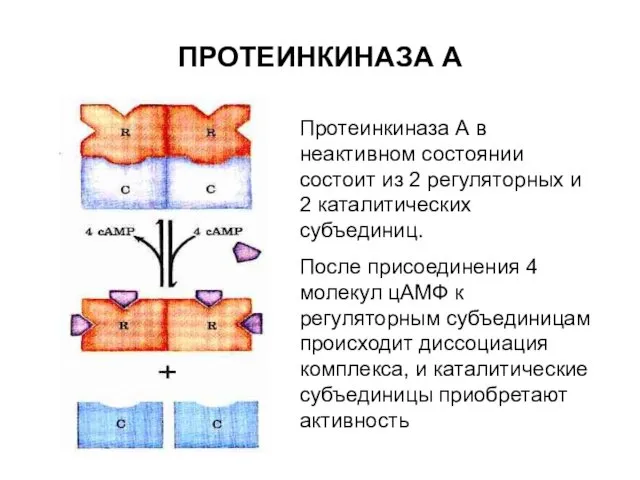
ПРОТЕИНКИНАЗА А
Протеинкиназа А в неактивном состоянии состоит из 2 регуляторных и
ПРОТЕИНКИНАЗА А
Протеинкиназа А в неактивном состоянии состоит из 2 регуляторных и
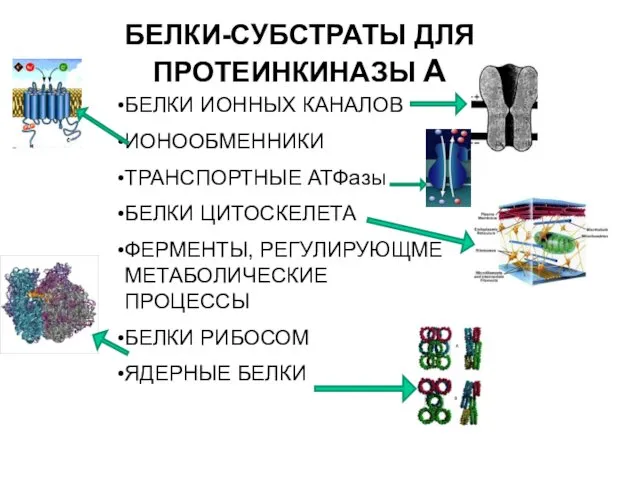
БЕЛКИ-СУБСТРАТЫ ДЛЯ ПРОТЕИНКИНАЗЫ А
БЕЛКИ ИОННЫХ КАНАЛОВ
ИОНООБМЕННИКИ
ТРАНСПОРТНЫЕ АТФазы
БЕЛКИ ЦИТОСКЕЛЕТА
ФЕРМЕНТЫ, РЕГУЛИРУЮЩМЕ МЕТАБОЛИЧЕСКИЕ ПРОЦЕССЫ
БЕЛКИ
БЕЛКИ-СУБСТРАТЫ ДЛЯ ПРОТЕИНКИНАЗЫ А
БЕЛКИ ИОННЫХ КАНАЛОВ
ИОНООБМЕННИКИ
ТРАНСПОРТНЫЕ АТФазы
БЕЛКИ ЦИТОСКЕЛЕТА
ФЕРМЕНТЫ, РЕГУЛИРУЮЩМЕ МЕТАБОЛИЧЕСКИЕ ПРОЦЕССЫ
БЕЛКИ
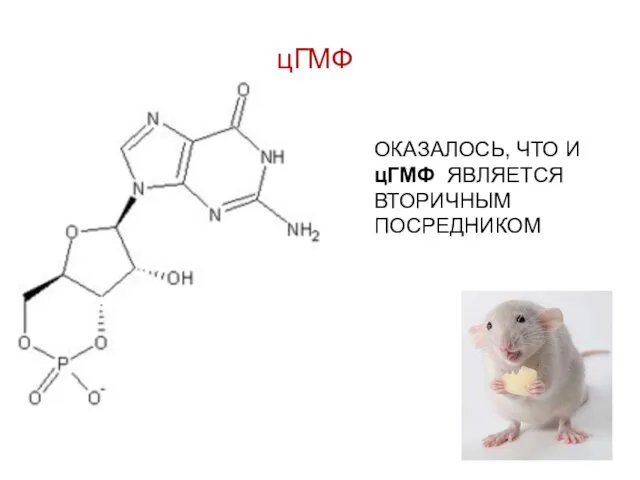
цГМФ
ОКАЗАЛОСЬ, ЧТО И цГМФ ЯВЛЯЕТСЯ ВТОРИЧНЫМ ПОСРЕДНИКОМ
цГМФ
ОКАЗАЛОСЬ, ЧТО И цГМФ ЯВЛЯЕТСЯ ВТОРИЧНЫМ ПОСРЕДНИКОМ
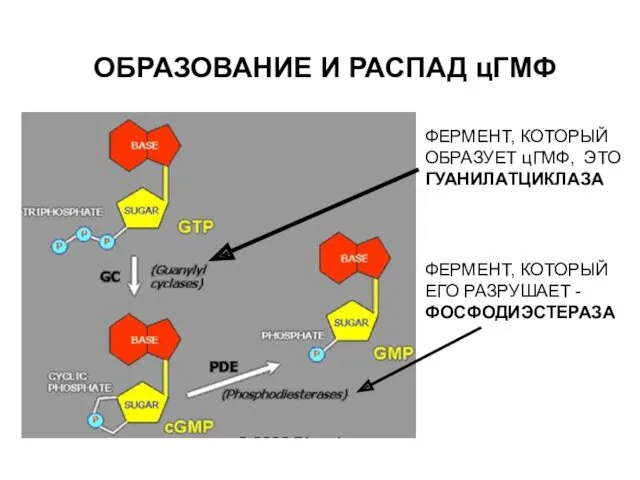
ОБРАЗОВАНИЕ И РАСПАД цГМФ
ФЕРМЕНТ, КОТОРЫЙ ОБРАЗУЕТ цГМФ, ЭТО ГУАНИЛАТЦИКЛАЗА
ФЕРМЕНТ, КОТОРЫЙ ЕГО
ОБРАЗОВАНИЕ И РАСПАД цГМФ
ФЕРМЕНТ, КОТОРЫЙ ОБРАЗУЕТ цГМФ, ЭТО ГУАНИЛАТЦИКЛАЗА
ФЕРМЕНТ, КОТОРЫЙ ЕГО
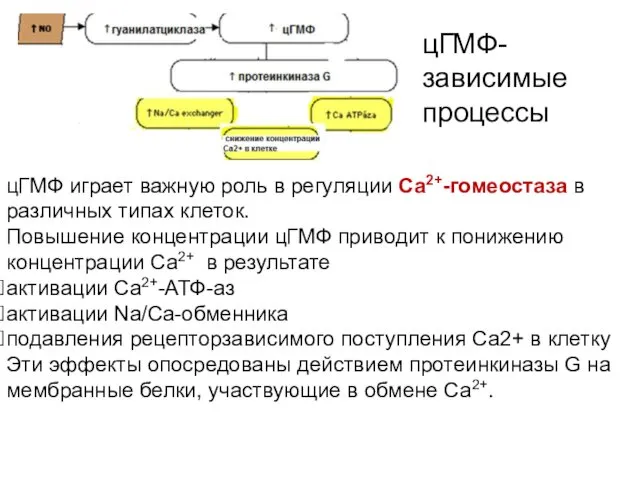
цГМФ играет важную роль в регуляции Са2+-гомеостаза в различных типах клеток.
цГМФ играет важную роль в регуляции Са2+-гомеостаза в различных типах клеток.

ПК G фосфорилирует Na,K-АТФазу эпителиоцитов почечных канальцев, что приводит к торможению
ПК G фосфорилирует Na,K-АТФазу эпителиоцитов почечных канальцев, что приводит к торможению
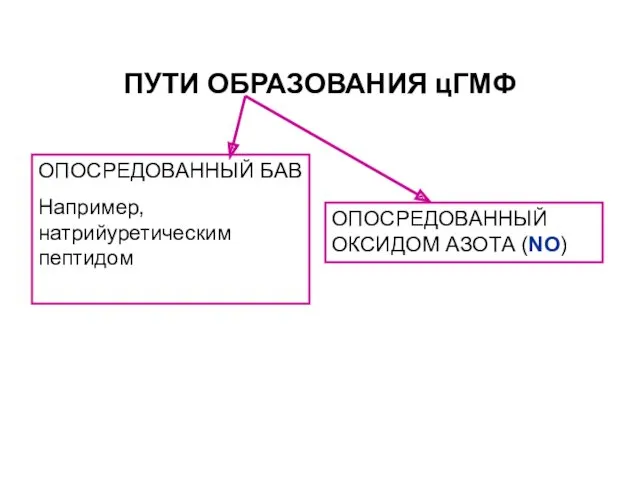
ПУТИ ОБРАЗОВАНИЯ цГМФ
ОПОСРЕДОВАННЫЙ БАВ
Например, натрийуретическим пептидом
ОПОСРЕДОВАННЫЙ ОКСИДОМ АЗОТА (NO)
ПУТИ ОБРАЗОВАНИЯ цГМФ
ОПОСРЕДОВАННЫЙ БАВ
Например, натрийуретическим пептидом
ОПОСРЕДОВАННЫЙ ОКСИДОМ АЗОТА (NO)
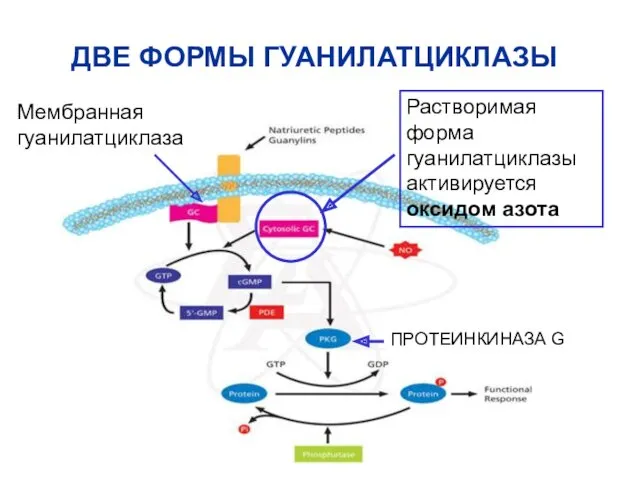
ДВЕ ФОРМЫ ГУАНИЛАТЦИКЛАЗЫ
Мембранная гуанилатциклаза
Растворимая форма гуанилатциклазы активируется оксидом азота
ПРОТЕИНКИНАЗА G
ДВЕ ФОРМЫ ГУАНИЛАТЦИКЛАЗЫ
Мембранная гуанилатциклаза
Растворимая форма гуанилатциклазы активируется оксидом азота
ПРОТЕИНКИНАЗА G

МЕХАНИЗМ ДЕЙСТВИЯ НАТРИЙУРЕТИЧЕСКОГО ПЕПТИДА
NaУП + R → ↑ГЦ → ↑цГМФ →
МЕХАНИЗМ ДЕЙСТВИЯ НАТРИЙУРЕТИЧЕСКОГО ПЕПТИДА
NaУП + R → ↑ГЦ → ↑цГМФ →

ОСОБЕННОСТИ цГМФ-ЗАВИСИМОЙ РЕГУЛЯЦИИ
ОТСУТСТВУЮТ G-БЕЛКИ
СВЯЗЫВАНИЕ АГОНИСТА С РЕЦЕПТОРОМ ВСЕГДА ПРИВОДИТ К АКТИВАЦИИ
ОСОБЕННОСТИ цГМФ-ЗАВИСИМОЙ РЕГУЛЯЦИИ
ОТСУТСТВУЮТ G-БЕЛКИ
СВЯЗЫВАНИЕ АГОНИСТА С РЕЦЕПТОРОМ ВСЕГДА ПРИВОДИТ К АКТИВАЦИИ
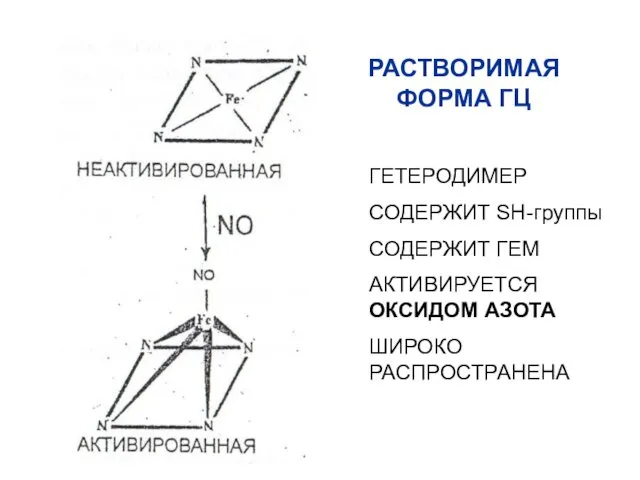
РАСТВОРИМАЯ ФОРМА ГЦ
ГЕТЕРОДИМЕР
СОДЕРЖИТ SH-группы
СОДЕРЖИТ ГЕМ
АКТИВИРУЕТСЯ ОКСИДОМ АЗОТА
ШИРОКО РАСПРОСТРАНЕНА
РАСТВОРИМАЯ ФОРМА ГЦ
ГЕТЕРОДИМЕР
СОДЕРЖИТ SH-группы
СОДЕРЖИТ ГЕМ
АКТИВИРУЕТСЯ ОКСИДОМ АЗОТА
ШИРОКО РАСПРОСТРАНЕНА
 Презентация Елабуга - жемчужина Прикамья
Презентация Елабуга - жемчужина Прикамья Мультимедийные технологии
Мультимедийные технологии Михаил Михайлович Сперанский. Сословная структура населения России по проекту М.М. Сперанского
Михаил Михайлович Сперанский. Сословная структура населения России по проекту М.М. Сперанского Сохранение церковного наследия в с. Красное
Сохранение церковного наследия в с. Красное Методическая разработка Дети Герои Великой отечественной войны
Методическая разработка Дети Герои Великой отечественной войны Content Brief
Content Brief Выступление из опыта работы по теме: Сказкотерапия как инновационная технология духовно-нравственного воспитания дошкольников
Выступление из опыта работы по теме: Сказкотерапия как инновационная технология духовно-нравственного воспитания дошкольников Воскресная школа покрова Пресвятой Богородицы
Воскресная школа покрова Пресвятой Богородицы Конституция Республики Казахстан
Конституция Республики Казахстан Родительское собрание Счастливая семья. Какая она?
Родительское собрание Счастливая семья. Какая она? Педагогический проект в стране знаков и правил
Педагогический проект в стране знаков и правил Пути введения лекарственных средств. Фармакокинетика
Пути введения лекарственных средств. Фармакокинетика Покупка имущества на торгах по банкротству
Покупка имущества на торгах по банкротству Свойства степени с натуральным показателем. 7 класс
Свойства степени с натуральным показателем. 7 класс Производство конструкционных материалов
Производство конструкционных материалов Мастер-класс ученика: Поздравительная открытка.
Мастер-класс ученика: Поздравительная открытка. Управление заказами
Управление заказами Аммиак. Соли аммония. Методы получения
Аммиак. Соли аммония. Методы получения Қазіргі жағдайда балық шаруашылығын ұйымдастыру
Қазіргі жағдайда балық шаруашылығын ұйымдастыру Radiation Biophysics
Radiation Biophysics Случаи сложения вида +5
Случаи сложения вида +5 23 февраля - День Защитника Отечества
23 февраля - День Защитника Отечества Сестринские манипуляции. Постановка согревающего компресса
Сестринские манипуляции. Постановка согревающего компресса 20231227_mandaloterapiya_1
20231227_mandaloterapiya_1 Кормление телят на ферме
Кормление телят на ферме Мой прадедушка: Герой на войне, Герой вне войны
Мой прадедушка: Герой на войне, Герой вне войны Септический шок
Септический шок Переработка Pb - руд
Переработка Pb - руд