- Межклеточные сигнальные вещества

Содержание
- 2. Гормональная регуляция обмена веществ Гормоны – биологически активные вещества разной химической природы, секретируются железами внутренней секреции
- 3. Химическая природа гормонов 1. Гидрофильные: белки, пептиды, произ-водные аминокислот (кроме тиреоидов) Гормоны гидрофильной природы рецептируются на
- 4. Этапы реализации действия гормонов 1. Регуляция синтеза гормонов (контроль нервной системы; тропная регуляция; механизм обратной связи)
- 5. Этапы реализации действия гормонов 5. Эффекторное звено: взаимоотношения с рецепторами. 6. Биотрансформация и период полужизни гормональных
- 6. Как гормоны изменяют метаболизм в клетках-мишенях? Непосредственное действие гормонов (гидрофобные) или с образованием вторичных мессенджеров –
- 7. Как гормоны изменяют метаболизм в клетках- мишенях? 2. Влияют на транскрипцию и последующий процессинг иРНК 3.
- 8. Основные типы мембранных рецепторов 1) Рецепторы (> 200) сопряженные с G -белковыми комплексами (G-protein coupled receptors
- 9. G – белковые комплексы > 20 G-белков (обладают GTP-азной активностью) Гетеротримеры (α, β, γ – субъединицы),
- 10. G – белковые комплексы Разные типы G –белков: Gs – стимулирует аденилатциклазу Gi – ингибирует аденилатциклазу
- 11. Вторичные мессенджеры биоактивных веществ Мессенджер Источник Эффект цАМФ – аденилатциклаза - активирует протеинкиназу А цГМФ –
- 12. Вторичные мессенджеры биоактивных веществ Мессенджер – Источник – Эффект Инозитолтрифосфат – фосфолипаза С – активирует Ca2+-каналы
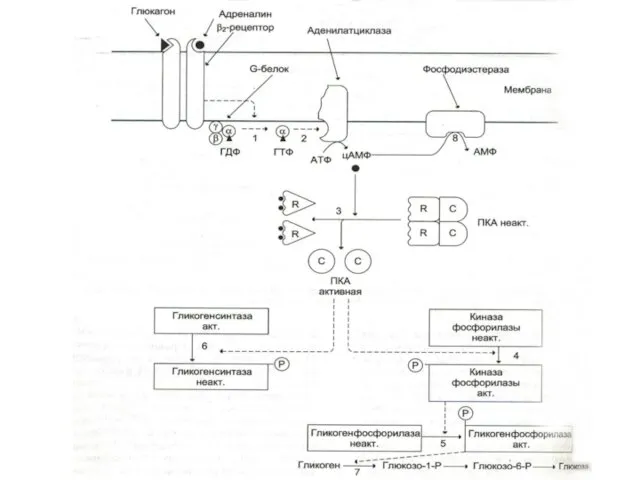
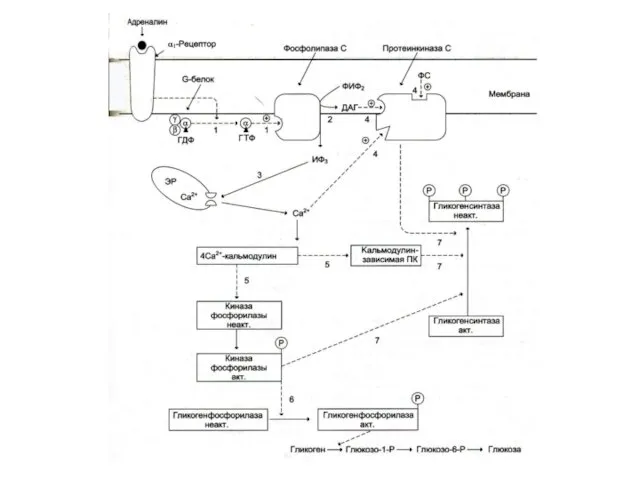
- 15. Фосфопротеинкиназы (ФПК) Активация протеинкиназ и последующее фосфорилирование ими разнообразных белковых субстратов вызывает широкий спектр эффектов вторичных
- 16. Фосфопротеинкиназы ФПКG – гомодимер, активируется аллостерически 4 молекулами цГМФ. ФПКС – гетеродимер (RC), R – субъединица
- 17. 2) Рецепторы = ионные каналы Связывание с лигандом приводит к изменению конформации рецептора, что позволяет специфическим
- 18. 3) Рецепторы, обладающие ферментативной активностью 1. Рецепторы, ассоциированные с гуанилатциклазной активностью 2. Рецепторы, ассоциированные с фосфатазной
- 19. 4) Рецепторы, не обладающие собственной каталитической активностью После связывания лиганда такие рецепторы связывают цитоплазматические протеинкиназы (они
- 20. Передача сигнала через внутриклеточные рецепторы Тиреоидные гормоны (йодированные производные тирозина) и стероиды проникают через цитоплазматическую мембрану
- 21. Гормоны гипоталамуса Нейропептиды гипоталамуса объединяют высшие отделы ЦНС и эндокринную систему По системе портальных сосудов поступают
- 22. Гормоны гипоталамуса Либерины (7 шт): тиролиберин (трипептид), гонадолиберин (декапептид), кортиколиберин (41 аминокислота), соматолиберин (40-44 аминокислот) и
- 23. Гормоны гипоталамуса Действие: 1) Нейропептиды рецептируются на поверхности соответствующих клеток гипофиза и активируют (либерины) или ингибируют
- 24. Гормоны гипоталамуса 3) Нейрогормоны (вазопрессин = АДГ и окситоцин) – сходные по структуре нонапептиды с дисульфидными
- 25. Гормоны гипоталамуса Рецепторы для вазопрессина, вероятно, аналогичны (V1) для АДГ. Мишенью для окситоцина являются клетки гладких
- 26. Гормоны гипофиза 1) соматотропный гормон, 2) пролактин сходные по структуре белки (191 и 199 остатков аминокислот).
- 27. Гормон роста, СТГ Единственный видоспецифичный из всех гормонов гипофиза. Синтез и секреция стимулируются соматолиберином, тиреолиберином, эндорфином,
- 28. Гормон роста, СТГ Рецепторы – на плазматической мембране скелетных и мышечных тканей и практически всех внутренних
- 29. Гормон роста, СТГ Первичные эффекты СТГ сходны с инсулином: запасание клетками глюкозы и активация липогенеза. Вторичные
- 30. Гормон роста, СТГ СТГ способствует биосинтезу инсулина в поджелудочной железе и соматомединов (ИФР) в клетках-мишенях. ИФР
- 31. 2) Пролактин Эстрогены в период беременности резко увеличивают количество лактотрофных клеток гипофиза. Импульсная секреция пролактина возрастает
- 32. ТТГ, ЛГ, ФСГ (ХГ) тиреотропный, лютеинизирующий, фолликулостимулирующий гормоны (ХГ – хорионический гонадотропин) Гликопротеины, димеры из α-
- 33. Проопиомеланокортин (ПОМК) Под влиянием кортиколиберина гипоталамуса в аденогипофизе синтезируется крупный белковый предшественник (265 аминокислот). Лимитированный протеолиз
- 34. Адренокортикотропный гормон (АКТГ) Пептид (39 аминокислот). Синтез стимулируется кортиколиберином, стрессовыми воздействиями. Циркадный ритм: минимум – вечером,
- 35. Адренокортикотропный гормон (АКТГ) АКТГ имеет срочные и отдаленные механизмы, надежно обеспечивающие реакцию Холестерол ⇨ПРЕГНЕНОЛОН Активирует аденилатциклазу,
- 36. Гормоны щитовидной железы Под влиянием ТТГ в фолликулах синтезируется тиреоглобулин (гликопротеин) и секретируется во внеклеточное пространство,
- 37. Три- и тетрайодтиронин (тироксин) Большая часть гормонов находится в плазме в связанном с белком (неактивном) состоянии.
- 38. Три- и тетрайодтиронин (тироксин) Липофильные йодированные производные тирозина проникают в клетки и связываются с ядерными рецепторами,
- 39. Паратиреоидный гормон Белок, 84 аминокислоты (синтез идет в виде препрогормона 115 АК). Секреция стимулируется снижением уровня
- 40. Паратиреоидный гормон Стимулированные паратгормоном остеобласты секретируют ИФР и цитокины, индуцирующие в остеокластах синтез коллагеназы и фосфатазы
- 41. Кальцитонин Низкомолекулярный белок (синтез в виде препрогормона из 136 аминокислот) Синтезируется в щитовидной, паращитовидных железах, тимусе,
- 42. Гормоны поджелудочной железы В островках Лангерганса: А клетки – глюкагон В клетки – инсулин Д клетки
- 43. инсулин Белок из двух полипептидных цепей (21 и 30 аминокислот), соединенных 2 дисульфидными мостиками. Может образовывать
- 44. Инсулин Стимулируют его синтез и секрецию – глюкоза, аргинин и лизин, гормоны ЖКТ, СТГ, кортизол, эстрогены.
- 45. Рецептор к инсулину Гликопротеиновый рецептор (IR) с тирозинкиназной активностью и способностью к автофосфорилированию и фосфорилированию целого
- 46. Глюкагон Полипептид (35 аминокислот). Препроглюкагон – белок (124 аминокислоты). Не найден транспортный белок. Угнетается секреция приемом
- 47. Надпочечники (МОЗГОВОЙ СЛОЙ) Левандовский (1899г.) выявил сходство эффектов экстракта надпочечников и активации симпатической нервной системы (мозговой
- 48. адреналин Катехоламины (адреналин в большей степени) осуществляют первую линию защиты организма при стрессе (гормон «бегства и
- 49. адреналин Секреция адреналина стимулируется тревожным состоянием, психическим возбуждением, гипоксией, гипогликемией. Тиреоидные гормоны увеличивают число β-адренорецепторов. Глюкокортикоиды
- 50. Синтез катехоламинов фенилаланин ? ДОФА ? дофамин ? норадреналин ? адреналин (ферменты – НАДФ-гидроксилазы, метилтрансфераза). Катехоламины
- 51. Инактивация катехоламинов Концентрация в крови – 0,05 нг/л, при стрессе – до 0,3 нг/л. При тяжелом
- 52. Метаболические эффекты адреналина Печень: активация гликогенфосфорилазы, нейрогенная острая гипергликемия. Одновременно – снижение секреции инсулина и продление
- 53. Стероиды коры надпочечников Под влиянием кортиколиберина гипоталамуса и АКТГ гипофиза (max – утром, min – вечером)
- 54. Инактивация стероидов Период полужизни: 0,5 – 1,5 часа. Кортикостероиды и андрогены выделяются в виде 17-кетостероидов (окисление
- 55. кортикостероиды Для кортикостероидов характерна дозозависимость, вплоть до противоположных эффектов, различное влияние на разные мишени (тканеспецифичность). Метаболические
- 56. Метаболические эффекты кортикостероидов Белковый обмен: ↓поглощения аминокислот (кроме печени), ↑протеолиза, ↓транскрипции и трансляции (иммунодепрессия), ↑экскреции аминокислот,
- 57. Минералокортикоид альдостерон Регуляция синтеза и секреции: АКТГ (в меньшей степени), в большей степени ↓[Na+] в крови
- 58. Половые стероиды Гонадолиберины гипоталамуса и гонадотропные гормоны гипофиза стимулируют синтез (через активацию стероидгидроксилазных ферментов) и секрецию
- 60. Скачать презентацию

Гормональная регуляция обмена веществ
Гормоны – биологически активные вещества
разной химической природы,
Гормональная регуляция обмена веществ
Гормоны – биологически активные вещества разной химической природы,

Химическая природа гормонов
1. Гидрофильные: белки, пептиды, произ-водные аминокислот (кроме тиреоидов)
Гормоны
Химическая природа гормонов
1. Гидрофильные: белки, пептиды, произ-водные аминокислот (кроме тиреоидов)
Гормоны

Этапы реализации действия гормонов
1. Регуляция синтеза гормонов
(контроль нервной системы; тропная
Этапы реализации действия гормонов
1. Регуляция синтеза гормонов (контроль нервной системы; тропная

Этапы реализации действия гормонов
5. Эффекторное звено: взаимоотношения с рецепторами.
6. Биотрансформация
Этапы реализации действия гормонов
5. Эффекторное звено: взаимоотношения с рецепторами.
6. Биотрансформация

Как гормоны изменяют метаболизм в клетках-мишенях?
Непосредственное действие гормонов (гидрофобные) или
Как гормоны изменяют метаболизм в клетках-мишенях?
Непосредственное действие гормонов (гидрофобные) или

Как гормоны изменяют метаболизм в клетках- мишенях?
2. Влияют на транскрипцию и
Как гормоны изменяют метаболизм в клетках- мишенях?
2. Влияют на транскрипцию и

Основные типы мембранных рецепторов
1) Рецепторы (> 200) сопряженные с G -белковыми
Основные типы мембранных рецепторов
1) Рецепторы (> 200) сопряженные с G -белковыми

G – белковые комплексы
> 20 G-белков (обладают GTP-азной активностью)
Гетеротримеры (α, β,
G – белковые комплексы
> 20 G-белков (обладают GTP-азной активностью)
Гетеротримеры (α, β,

G – белковые комплексы
Разные типы G –белков:
Gs – стимулирует аденилатциклазу
Gi –
G – белковые комплексы
Разные типы G –белков:
Gs – стимулирует аденилатциклазу
Gi –

Вторичные мессенджеры биоактивных веществ
Мессенджер Источник Эффект
цАМФ – аденилатциклаза - активирует
протеинкиназу
Вторичные мессенджеры биоактивных веществ
Мессенджер Источник Эффект
цАМФ – аденилатциклаза - активирует
протеинкиназу

Вторичные мессенджеры биоактивных веществ
Мессенджер – Источник – Эффект
Инозитолтрифосфат – фосфолипаза
Вторичные мессенджеры биоактивных веществ
Мессенджер – Источник – Эффект
Инозитолтрифосфат – фосфолипаза

Фосфопротеинкиназы (ФПК)
Активация протеинкиназ и последующее фосфорилирование ими разнообразных белковых субстратов
Фосфопротеинкиназы (ФПК)
Активация протеинкиназ и последующее фосфорилирование ими разнообразных белковых субстратов

Фосфопротеинкиназы
ФПКG – гомодимер, активируется аллостерически 4 молекулами цГМФ.
ФПКС – гетеродимер (RC),
Фосфопротеинкиназы
ФПКG – гомодимер, активируется аллостерически 4 молекулами цГМФ.
ФПКС – гетеродимер (RC),

2) Рецепторы = ионные каналы
Связывание с лигандом приводит к изменению конформации
2) Рецепторы = ионные каналы
Связывание с лигандом приводит к изменению конформации

3) Рецепторы, обладающие ферментативной активностью
1. Рецепторы, ассоциированные с гуанилатциклазной активностью
2.
3) Рецепторы, обладающие ферментативной активностью
1. Рецепторы, ассоциированные с гуанилатциклазной активностью
2.

4) Рецепторы, не обладающие собственной каталитической активностью
После связывания лиганда
такие рецепторы
4) Рецепторы, не обладающие собственной каталитической активностью
После связывания лиганда такие рецепторы

Передача сигнала через внутриклеточные рецепторы
Тиреоидные гормоны (йодированные производные тирозина) и стероиды
Передача сигнала через внутриклеточные рецепторы
Тиреоидные гормоны (йодированные производные тирозина) и стероиды

Гормоны гипоталамуса
Нейропептиды гипоталамуса
объединяют высшие отделы ЦНС
и эндокринную систему
По системе
Гормоны гипоталамуса
Нейропептиды гипоталамуса
объединяют высшие отделы ЦНС
и эндокринную систему
По системе

Гормоны гипоталамуса
Либерины (7 шт): тиролиберин (трипептид), гонадолиберин (декапептид), кортиколиберин (41 аминокислота),
Гормоны гипоталамуса
Либерины (7 шт): тиролиберин (трипептид), гонадолиберин (декапептид), кортиколиберин (41 аминокислота),

Гормоны гипоталамуса
Действие:
1) Нейропептиды рецептируются на поверхности соответствующих клеток гипофиза и активируют
Гормоны гипоталамуса
Действие:
1) Нейропептиды рецептируются на поверхности соответствующих клеток гипофиза и активируют

Гормоны гипоталамуса
3) Нейрогормоны (вазопрессин = АДГ и окситоцин) – сходные по
Гормоны гипоталамуса
3) Нейрогормоны (вазопрессин = АДГ и окситоцин) – сходные по

Гормоны гипоталамуса
Рецепторы для вазопрессина, вероятно, аналогичны (V1) для АДГ.
Мишенью для
Гормоны гипоталамуса
Рецепторы для вазопрессина, вероятно, аналогичны (V1) для АДГ.
Мишенью для

Гормоны гипофиза
1) соматотропный гормон, 2) пролактин
сходные по структуре белки (191
Гормоны гипофиза
1) соматотропный гормон, 2) пролактин
сходные по структуре белки (191

Гормон роста, СТГ
Единственный видоспецифичный из всех гормонов гипофиза.
Синтез и секреция стимулируются
Гормон роста, СТГ
Единственный видоспецифичный из всех гормонов гипофиза.
Синтез и секреция стимулируются

Гормон роста, СТГ
Рецепторы – на плазматической мембране скелетных и мышечных тканей
Гормон роста, СТГ
Рецепторы – на плазматической мембране скелетных и мышечных тканей

Гормон роста, СТГ
Первичные эффекты СТГ сходны с инсулином: запасание клетками глюкозы
Гормон роста, СТГ
Первичные эффекты СТГ сходны с инсулином: запасание клетками глюкозы

Гормон роста, СТГ
СТГ способствует биосинтезу инсулина в поджелудочной железе и соматомединов
Гормон роста, СТГ
СТГ способствует биосинтезу инсулина в поджелудочной железе и соматомединов

2) Пролактин
Эстрогены в период беременности резко увеличивают количество лактотрофных клеток гипофиза.
Импульсная
2) Пролактин
Эстрогены в период беременности резко увеличивают количество лактотрофных клеток гипофиза.
Импульсная

ТТГ, ЛГ, ФСГ (ХГ)
тиреотропный, лютеинизирующий, фолликулостимулирующий гормоны
(ХГ – хорионический гонадотропин)
Гликопротеины,
ТТГ, ЛГ, ФСГ (ХГ)
тиреотропный, лютеинизирующий, фолликулостимулирующий гормоны
(ХГ – хорионический гонадотропин)
Гликопротеины,

Проопиомеланокортин (ПОМК)
Под влиянием кортиколиберина гипоталамуса в аденогипофизе синтезируется крупный белковый предшественник
Проопиомеланокортин (ПОМК)
Под влиянием кортиколиберина гипоталамуса в аденогипофизе синтезируется крупный белковый предшественник

Адренокортикотропный гормон (АКТГ)
Пептид (39 аминокислот).
Синтез стимулируется кортиколиберином, стрессовыми воздействиями.
Циркадный
Адренокортикотропный гормон (АКТГ)
Пептид (39 аминокислот).
Синтез стимулируется кортиколиберином, стрессовыми воздействиями.
Циркадный

Адренокортикотропный гормон (АКТГ)
АКТГ имеет срочные и отдаленные механизмы, надежно обеспечивающие реакцию
Адренокортикотропный гормон (АКТГ)
АКТГ имеет срочные и отдаленные механизмы, надежно обеспечивающие реакцию

Гормоны щитовидной железы
Под влиянием ТТГ в фолликулах синтезируется тиреоглобулин (гликопротеин) и
Гормоны щитовидной железы
Под влиянием ТТГ в фолликулах синтезируется тиреоглобулин (гликопротеин) и

Три- и тетрайодтиронин (тироксин)
Большая часть гормонов находится в плазме в связанном
Три- и тетрайодтиронин (тироксин)
Большая часть гормонов находится в плазме в связанном

Три- и тетрайодтиронин (тироксин)
Липофильные йодированные производные тирозина проникают в клетки и
Три- и тетрайодтиронин (тироксин)
Липофильные йодированные производные тирозина проникают в клетки и

Паратиреоидный гормон
Белок, 84 аминокислоты (синтез идет в виде препрогормона 115 АК).
Секреция
Паратиреоидный гормон
Белок, 84 аминокислоты (синтез идет в виде препрогормона 115 АК).
Секреция

Паратиреоидный гормон
Стимулированные паратгормоном остеобласты секретируют ИФР и цитокины, индуцирующие в остеокластах
Паратиреоидный гормон
Стимулированные паратгормоном остеобласты секретируют ИФР и цитокины, индуцирующие в остеокластах

Кальцитонин
Низкомолекулярный белок (синтез в виде препрогормона из 136 аминокислот)
Синтезируется в щитовидной,
Кальцитонин
Низкомолекулярный белок (синтез в виде препрогормона из 136 аминокислот)
Синтезируется в щитовидной,

Гормоны поджелудочной железы
В островках Лангерганса:
А клетки – глюкагон
В клетки –
Гормоны поджелудочной железы
В островках Лангерганса:
А клетки – глюкагон
В клетки –

инсулин
Белок из двух полипептидных цепей
(21 и 30 аминокислот), соединенных
2
инсулин
Белок из двух полипептидных цепей (21 и 30 аминокислот), соединенных 2

Инсулин
Стимулируют его синтез и секрецию – глюкоза, аргинин и лизин, гормоны
Инсулин
Стимулируют его синтез и секрецию – глюкоза, аргинин и лизин, гормоны

Рецептор к инсулину
Гликопротеиновый рецептор (IR) с тирозинкиназной активностью и способностью к
Рецептор к инсулину
Гликопротеиновый рецептор (IR) с тирозинкиназной активностью и способностью к

Глюкагон
Полипептид (35 аминокислот).
Препроглюкагон – белок (124 аминокислоты).
Не найден транспортный белок.
Угнетается секреция
Глюкагон
Полипептид (35 аминокислот).
Препроглюкагон – белок (124 аминокислоты).
Не найден транспортный белок.
Угнетается секреция

Надпочечники
(МОЗГОВОЙ СЛОЙ)
Левандовский (1899г.) выявил сходство эффектов экстракта надпочечников и активации симпатической
Надпочечники
(МОЗГОВОЙ СЛОЙ)
Левандовский (1899г.) выявил сходство эффектов экстракта надпочечников и активации симпатической

адреналин
Катехоламины (адреналин в большей степени) осуществляют первую линию защиты организма при
адреналин
Катехоламины (адреналин в большей степени) осуществляют первую линию защиты организма при

адреналин
Секреция адреналина стимулируется тревожным состоянием, психическим возбуждением, гипоксией, гипогликемией.
Тиреоидные гормоны увеличивают
адреналин
Секреция адреналина стимулируется тревожным состоянием, психическим возбуждением, гипоксией, гипогликемией.
Тиреоидные гормоны увеличивают

Синтез катехоламинов
фенилаланин ? ДОФА ? дофамин ? норадреналин ? адреналин (ферменты
Синтез катехоламинов
фенилаланин ? ДОФА ? дофамин ? норадреналин ? адреналин (ферменты

Инактивация катехоламинов
Концентрация в крови – 0,05 нг/л, при стрессе – до
Инактивация катехоламинов
Концентрация в крови – 0,05 нг/л, при стрессе – до

Метаболические эффекты адреналина
Печень: активация гликогенфосфорилазы, нейрогенная острая гипергликемия.
Одновременно – снижение
Метаболические эффекты адреналина
Печень: активация гликогенфосфорилазы, нейрогенная острая гипергликемия.
Одновременно – снижение

Стероиды коры надпочечников
Под влиянием кортиколиберина гипоталамуса и АКТГ гипофиза (max –
Стероиды коры надпочечников
Под влиянием кортиколиберина гипоталамуса и АКТГ гипофиза (max –

Инактивация стероидов
Период полужизни: 0,5 – 1,5 часа.
Кортикостероиды и андрогены выделяются в
Инактивация стероидов
Период полужизни: 0,5 – 1,5 часа.
Кортикостероиды и андрогены выделяются в

кортикостероиды
Для кортикостероидов характерна дозозависимость, вплоть до противоположных эффектов, различное влияние на
кортикостероиды
Для кортикостероидов характерна дозозависимость, вплоть до противоположных эффектов, различное влияние на

Метаболические эффекты кортикостероидов
Белковый обмен:
↓поглощения аминокислот (кроме печени), ↑протеолиза, ↓транскрипции и
Метаболические эффекты кортикостероидов
Белковый обмен:
↓поглощения аминокислот (кроме печени), ↑протеолиза, ↓транскрипции и

Минералокортикоид альдостерон
Регуляция синтеза и секреции:
АКТГ (в меньшей степени), в большей
Минералокортикоид альдостерон
Регуляция синтеза и секреции: АКТГ (в меньшей степени), в большей

Половые стероиды
Гонадолиберины гипоталамуса и гонадотропные гормоны гипофиза стимулируют синтез (через активацию
Половые стероиды
Гонадолиберины гипоталамуса и гонадотропные гормоны гипофиза стимулируют синтез (через активацию
 Презентация Портфолио воспитанника ДОУ
Презентация Портфолио воспитанника ДОУ Конституційне право України
Конституційне право України Презентация к уроку окружающий мир 3 класс на тему Разнообразие животных
Презентация к уроку окружающий мир 3 класс на тему Разнообразие животных Настольные игры
Настольные игры Псевдоперитонеальды синдром
Псевдоперитонеальды синдром Вина-специалитеты
Вина-специалитеты Protection of animals - activity aimed at improving the content and treatment of animals, to prevent cruelty to animals
Protection of animals - activity aimed at improving the content and treatment of animals, to prevent cruelty to animals Арабское лекарствоведение
Арабское лекарствоведение Актуальные проблемы дисциплин экономики труда и управления персоналом организации: обзор тематики за 2015-2017 годы
Актуальные проблемы дисциплин экономики труда и управления персоналом организации: обзор тематики за 2015-2017 годы Начало реформации в Европе
Начало реформации в Европе Joinery work on the roof
Joinery work on the roof Решение квадратных уравнений различного вида разными способами
Решение квадратных уравнений различного вида разными способами Отношение российского общества к употреблению феминитивов в речи и естественному изменению языка
Отношение российского общества к употреблению феминитивов в речи и естественному изменению языка Урок географии в 7 классе по теме Страны Северной Европы
Урок географии в 7 классе по теме Страны Северной Европы international trade of canada
international trade of canada Пассажирские перевозки на железнодорожном транспорте
Пассажирские перевозки на железнодорожном транспорте Презентация Порядок заполнения электронами уровней и подуровней Диск
Презентация Порядок заполнения электронами уровней и подуровней Диск Использование ИКТ на уроках в коррекционной школе VIII вида
Использование ИКТ на уроках в коррекционной школе VIII вида Экосистемы и человеческие экосистемы. Их сходства и различия
Экосистемы и человеческие экосистемы. Их сходства и различия Развитие и коррекция познавательной деятельности детей через дидактические игры.
Развитие и коррекция познавательной деятельности детей через дидактические игры. Использование игровых методов при принятии решений в условиях риска и неопределенности понятие об игровых методах. Тема 8
Использование игровых методов при принятии решений в условиях риска и неопределенности понятие об игровых методах. Тема 8 Час Земли
Час Земли Семиотика урологических заболеваений
Семиотика урологических заболеваений Герои Афганской войны (25 декабря 1979 – 15 февраля 1989)
Герои Афганской войны (25 декабря 1979 – 15 февраля 1989) Новые виды низкоинтенсивной физиотерапии
Новые виды низкоинтенсивной физиотерапии Introduction into botany. Structure of plant cell. Plant tissues and their classification
Introduction into botany. Structure of plant cell. Plant tissues and their classification Параметры и характеристики приемных устройств
Параметры и характеристики приемных устройств Виды киноискусства. Документальное кино
Виды киноискусства. Документальное кино