- Молекулярная биология. Курс лекций

Содержание
- 2. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Содержание: Основные этапы возникновения и развития молекулярной биологии. Методы
- 3. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Содержание: Репарация повреждений ДНК. Генетическая рекомбинация. Экспрессионный аппарат клетки.
- 4. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Лекция 1. Основные этапы возникновения и развития молекулярной биологии.
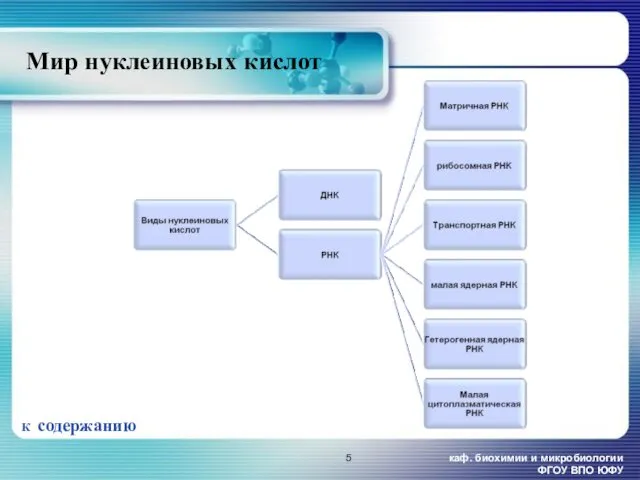
- 5. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Мир нуклеиновых кислот К содержанию
- 6. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Информационная связь между ДНК, РНК и белками К содержанию
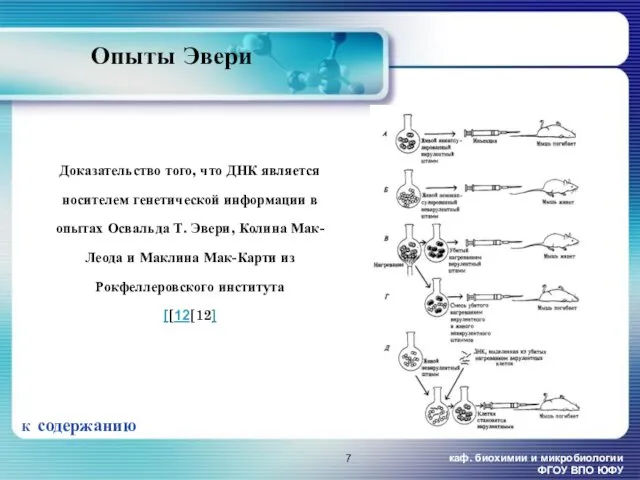
- 7. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ К содержанию Опыты Эвери Доказательство того, что ДНК является
- 8. Общая схема эксперимента Альфреда Д. Херши и Марты Чейз в опытах с применением радиоактивных меток (1952
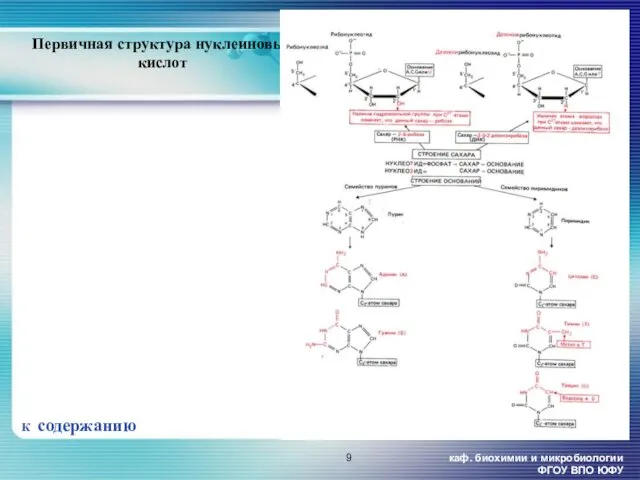
- 9. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Первичная структура нуклеиновых кислот К содержанию
- 10. Нуклеиновые кислоты являются биологическими полимерами, мономерными звеньями которых являются нуклеотиды – фосфорные эфиры нуклеозидов, которые, в
- 11. Азотистые основания, входящие в состав нуклеотидов ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Пиримидиновые Пуриновые
- 12. Первичная структура молекулы ДНК и РНК ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Первичная структура молекулы
- 13. Взаимодействия между гетероциклическими основаниями в нуклеиновых кислотах [12]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Специфическая
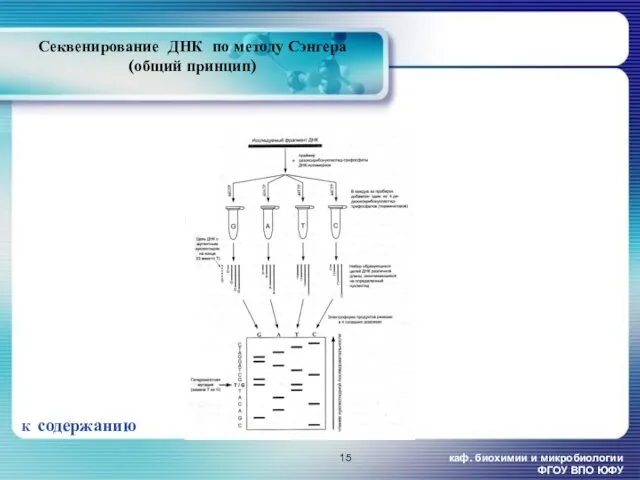
- 14. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Методы определения первичной структуры ДНК. Метод секвенирования ДНК по
- 15. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Секвенирование ДНК по методу Сэнгера (общий принцип) К содержанию
- 16. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Результаты секвенирования ДНК/РНК К содержанию
- 17. Физико-химические свойства ДНК. Денатурация и ренатурация ДНК [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Денатурация
- 18. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Лекция 2. Молекулярная организация генетического материала биологических систем. К
- 19. Постулирование модели двойной спирали ДНК американским генетиком Джеймсом Уотсоном и английским физиком Френсисом Криком [7]. ГОУ
- 20. Дж. Уотсон и Ф. Крик у одной из своих моделей ДНК (Кембридж. 1963 год) ГОУ ВПО
- 21. Идеализированная картина дифракции ДНК. Результаты этих измерений привели к построению модели ДНК. ГОУ ВПО ЮФУ, каф.
- 22. Вторичная структура ДНК – двойная спираль. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 23. Спиральная структура молекулы ДНК [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Молекула ДНК обычно находится
- 24. Полиморфизм двойной спирали ДНК [1, 7]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии А, В, С,
- 25. Характеристика конформационных состояний ДНК ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 26. Упаковка ДНК. Гистоны и нуклеосомы [7, 10]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 27. Характеристика гистонов, входящих в состав гистонового октамера ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 28. Уровни компактизации молекулы ДНК [4]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Свободная ДНК скручивается вокруг
- 29. Разнообразие форм ДНК [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Линейная двухцепочечная ДНК Кольцевая двухцепочечная
- 30. ДНК кинетопласта из Leishmania tarentolae [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Электронная микрофотография ДНК
- 31. Сверхспирализация ДНК. Топоизомеразы. Репликация ДНК сталкивается с серьёзными топологическим проблемами: возникновение положительной (+) или отрицательной (-)
- 32. Реакции, катализируемые топоизомеразами типа 1 [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 33. Реакции, катализируемые топоизомеразами типа 2 ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Топоизомеразы типа 2 катализируют
- 34. Работа ДНК-гиразы ГОУ ВПО ЮФУ, каф. биохимии и микробиологии В бактериях обнаружена ДНК-гираза. Показано образование отрицательных
- 35. Рибонуклеиновые кислоты. Основные виды и функции. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 36. Сравнительная структура ДНК и РНК [5]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 37. Вторичная и третичная структура однотяжевых РНК [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Структурная схема
- 38. Типы «третичных» внутримолекулярных взаимодействий. между отдельными элементами вторичной структуры молекулы РНК [Типы «третичных» внутримолекулярных взаимодействий. между
- 39. Структура тРНК [12]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Последовательность тРНК включает 70-90 нуклеотидов и
- 40. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Лекция 3. Мозаичная организация генома эукариот и вирусных систем.
- 41. Строение прокариотических генов [1 ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 42. Оперонная организация генов прокариот [1]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 43. Гетеродуплексы ДНК - мРНК ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Показаны петли, образующиеся в ходе
- 44. Организация прерывистого гена ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Организация прерывистого гена ДНК, содержащую ген
- 45. Структура β – глобинового гена и созревание его транскрипта ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 46. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Строение эукариотического гена, кодирующего белок [1].
- 47. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Организация генетического материала различных форм живой материи
- 48. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Передача генетического материала в клетке Процесс передачи генетической информации
- 49. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Виды сплайсинга Образование ковалентных связей между фрагментами ДНК, синтезированными
- 50. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Аутосплайсинг 26 S рРНК у тетрахимены в процессе образования
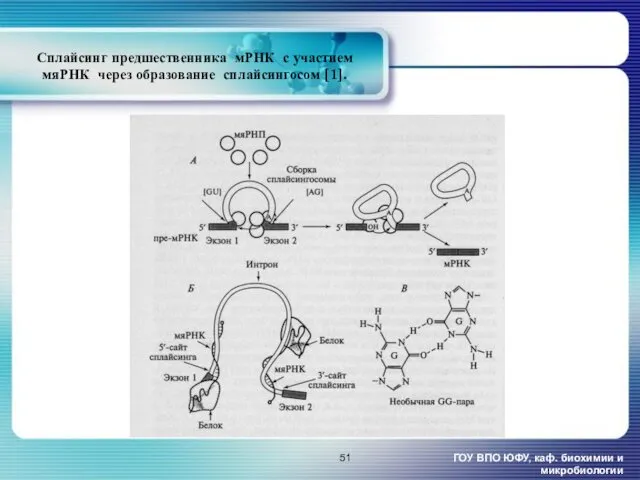
- 51. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Сплайсинг предшественника мРНК с участием мяРНК через образование сплайсингосом
- 52. Механизмы ферментативного сплайсинга ядерных генов животных. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 53. Альтернативный сплайсинг при экспрессии гена кальцитонина крысы. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 54. Типы альтернативного сплайсинга пре-мРНК [1]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 55. Процессинг тРНК. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Процессинг тРНК из молекулы-предшественника тРНК (Tyr) E.
- 56. Процессинг рРНК у прокариот. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Б. Спейсерные последовательности, расположенные в
- 57. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Лекция 4. Нестабильность генома. Мобильные генетические элементы про- и
- 58. Классификация мобильных элементов прокариот. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Транспозоны Tn 3- подобные элементы
- 59. Классификация мобильных элементов прокариот. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии IS-элементы – это сегменты ДНК,
- 60. Типичные IS-элементы E. coli [1] ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 61. Принципиальное строение составного транспозона Tn10. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 62. R-плазмида со всеми инсерционными последовательностями (IS) и транспозонами (Tn) [2] ГОУ ВПО ЮФУ, каф. биохимии и
- 63. Механизмы перемещения мобильных элементов бактерий [1]
- 64. Механизм транспозиции Tn
- 65. Расщепление обеих цепей эндонуклеазой. Места расщеплений отстоят друг от друга на несколько пар оснований, в результате
- 66. Эукариотические транспозоны
- 67. Ретротранспозон класса 1 и его транскрипт [2]
- 68. Модель транспозиции ретрогенов при участии РНК-посредника [2] В результате транскрипции и процессинга, включающего сплайсинг интронов,образуется функциональная
- 69. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Свойства транспозонов варьирует от 10-7 до 10-4 и зависит
- 70. Биологическая роль транспозонов
- 71. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Лекция 5. Репликация ДНК. Особенности биосинтеза вирусных нуклеиновых кислот.
- 72. Предполагаемые модели репликации дуплексной ДНК [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии консервативный полуконсервативный дисперсионный
- 73. Экспериментальные подтверждения модели Уотсона-Крика в работах Мэтью Мезельсона и Франклина Сталя [12]. ГОУ ВПО ЮФУ, каф.
- 74. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии А. Распределение плотности молекул ДНК, наблюдавшееся Мезелсоном и Сталем
- 75. Удлинение цепи ДНК. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Удлинение цепи происходит путём последовательного присоединения
- 76. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии ДНК - полимеразы.
- 77. ДНК-полимеразы ГОУ ВПО ЮФУ, каф. биохимии и микробиологии ДНК-полимераза — фермент, участвующий в репликации ДНк. Ферменты
- 78. ДНК-полимераза -3 ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 79. Белки, входящие в состав репликативного комплекса ]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 80. Инициация репликации ДНК [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 81. Полуконсервативный механизм репликации ДНК E. coli [9]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 82. Репликация ДНК [Репликация ДНК [ 7Репликация ДНК [ 7]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 83. ДНК-лигаза. Механизм лигазной реакции [12]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 84. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Репликативные механизмы различных форм живой материи
- 85. Репликация хромосомной ДНК эукариот [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Репликация идёт в двух
- 86. Начало репликации [2]. Репликация инициируется в специфическом участае ДНК, точке начала репликации (ori). Репликативная вилка перемещается
- 87. Репликация по типу катящегося кольца [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 88. Поочерёдная репликация цепей ДНК аденовируса [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 89. Репликация теломерных участков эукариотических хромосом [1]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии А- возникновение недореплицирования
- 90. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Общий принцип полимеразной цепной реакции. Денатурация двойной спирали ДНК-матрицы
- 91. Сравнительная структура ДНК и РНК [5]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 92. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Лекция 6. Репарация повреждений ДНК. К содержанию
- 93. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Изменения генома. К содержанию Мобильные элементы генома
- 94. Мутагенные агенты и типы повреждений молекулы ДНК [4]. ФГОУ ВПО ЮФУ, каф. биохимии и микробиологии 1.
- 95. Повреждения в нуклеотидах ДНК. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Дезаминирование Апуринизация
- 96. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Повреждения в нуклеотидах ДНК [1]. Метилирование Размыкание кольца Окисление
- 97. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Типы репарации. К содержанию Эксцизионная репарация Для репарации необходима
- 98. Прямая репарация [1] ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Дезалкилирование О6-метилгуаниновых остатков катализируется специфической О6-метилгуанозиналкилтрансферазой
- 99. Схема эксцизионной репарации [1]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии ДНК-N-гликозилаза удаляет повреждённое основание, АР-эндонуклеаза
- 100. Схема нуклеотидной эксцизионной репарации у E. coli с участием эксинуклеазы [1]. ГОУ ВПО ЮФУ, каф. биохимии
- 101. Ферментативная репарация циклобутановых тиминовых димеров [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Ферментативный комплекс uvrABC
- 102. Репарация с помощью N- гликозилаз. [2] ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 103. Делеции и амплификации, возникающие при репликации или репарации [1]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 104. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Некоторые гены SOS-системы и результат их индукции. К содержанию
- 105. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Лекция 7. Генетическая рекомбинация. К содержанию
- 106. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Типы рекомбинации.
- 107. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Рекомбинация сестринских хроматид периферических лимфоцитов человека
- 108. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Гомологичная рекомбинация Комплементарное спаривание цепей, принадлежащих разным дуплексам обеспечивает
- 109. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Структура Р. Холидея или полухиазма. При рекомбинации происходит обмен
- 110. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Миграция ветви.
- 111. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Изомеризация полухиазмы. Если развернуть полухиазму в направлении, указанном стрелками,
- 112. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Способы разрезания полухиазмы.
- 113. Общая рекомбинация с образованием двухцепочечного разрыва [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 114. Сайт-специфическая рекомбинация [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Специфические последовательности имеются в обеих участвующих
- 115. Нереципрокная гомологичная рекомбинация как один из этапов репарации ДНК в месте образования пиримидиновых димеров [2]. ГОУ
- 116. RecA-зависимая общая рекомбинация ДНК E/ coli [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 117. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Лекция 8. Экспрессионный аппарат клетки. Транскрипция. К содержанию
- 118. Пространственная модель комплекса РНК-полимеразы E. coli с фрагментом ДНК-матрицы [9]. ГОУ ВПО ЮФУ, каф. биохимии и
- 119. Схема синтеза РНК с помощью РНК-полимеразы [9]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии РНК –
- 120. Функциональные участки ДНК-зависимой РНК-полимеразы [ ДНК-зависимой РНК-полимеразы [8 ДНК-зависимой РНК-полимеразы [8] ГОУ ВПО ЮФУ, каф. биохимии
- 121. Типичный промотор E. coli [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Транскрипция инициируется при образовании
- 122. Взаимное расположение блока Прибнова и блока -35 [ блока Прибнова и блока -35 [11 блока Прибнова
- 123. Очерёдность процессов транскрипции, трансляции и деградации иРНК у бактерий [8] . ГОУ ВПО ЮФУ, каф. биохимии
- 124. Схема транскрипции, катализируемой РНК-полимеразой [ катализируемой РНК-полимеразой [1 катализируемой РНК-полимеразой [1]. ГОУ ВПО ЮФУ, каф. биохимии
- 125. Терминация транскрипции. Примеры шпилек в ρ-независимых и ρ-зависимых терминаторах [2]. ГОУ ВПО ЮФУ, каф. биохимии и
- 126. Влияние ρ-белка на остановку РНК-полимеразы в терминаторе, сопровождающуюся терминацией транскрипции с отделением фермента и синтезированной к
- 127. Эукариотическая иРНК [Эукариотическая иРНК [11Эукариотическая иРНК [11]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Эукариотическая иРНК
- 128. Структура «кэпов», расположенных на 5`- конце эукариотических иРНК [ на 5`- конце эукариотических иРНК [11 на
- 129. Обратная транскрипция [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Схематическое изображение обратной транскрипции ретровирусной РНК,
- 130. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Лекция 9. Генетический код. Молекулярная организация тРНК и рибосом
- 131. Свойства генетического кода: ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Генетический код является квазидуплетным. В генетическом
- 132. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Таблица генетического кода К содержанию
- 133. AUG – Start-кодоны (на 5`- конце мРНК кодирует стартовую аминокислоту f-Met-tRNA UGA, UAA, UAG – Stop
- 134. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Отклонения универсальности генетического кода. К содержанию Ile
- 135. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Вырожденность генетического кода К содержанию
- 136. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Гипотеза неоднозначного соответствия Ф. Крика (1956) (гипотеза «качения» К
- 137. Рамка считывания. Поскольку каждый кодон содержит три нуклеотида, один и тот же генетический текст можно прочитать
- 138. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Структурно-функциональная организация тРНК. К содержанию
- 139. Образование аминоацил-тРНК ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 140. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Состав прокариотических и эукариотических рибосом. К содержанию
- 141. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Схема строения рибосомы бактерий по А. С. Спирину, 1986.
- 142. Строение рибосомы ФГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 143. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Лекция 10. Трансляция: условия и энергетика. Рабочий цикл рибосомы.
- 144. Основные участники этапов трансляции. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 145. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Ферменты (факторы) трансляции про- и эукариот. К содержанию
- 146. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Схема последовательности событий в процессе инициации трансляции прокариот. К
- 147. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ «Рабочий» (элонгационный) цикл рибосомы К содержанию
- 148. каф. биохимии и микробиологии ФГОУ ВПО ЮФУ Терминация трансляции прокариот. К содержанию
- 149. Лактозный оперон E. coli и ген lac-репрессора [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 150. Репрессия lac-оперона гомотетрамерным lac-репрессором и индукция lac-оперона после связывания с репрессором β-галактозидного индуктора. ГОУ ВПО ЮФУ,
- 151. Нуклеотидные последовательности, принимающие участие в регуляции экспресии lac-оперона. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 152. «Цинковые пальцы» - элементы сверхвторичной структуры в ДНК-связывающих белках, участвующих в регуляции транскрипции [1]. ГОУ ВПО
- 153. Схема взаимодействия димерной молекулы фактора транскрипции, содержащей «лейциновые молнии», с молекулой ДНК. ГОУ ВПО ЮФУ, каф.
- 154. Комплексы белков, участвующих в регуляции транскрипции у эукариот. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии R-
- 155. Терминация транскрипции. Примеры шпилек в ρ-независимых и ρ-зависимых терминаторах. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
- 156. Влияние ρ-белка на остановку РНК-полимеразы в терминаторе, сопровождающуюся терминацией транскрипции с отделением фермента и синтезированной к
- 157. Список литературных источников. Коничев А. С., Севастьянова Г. А. Молекулярная биология. М.: Издательский центр «Академия», 2003.
- 159. Скачать презентацию

каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Содержание:
Основные этапы возникновения и развития
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Содержание:
Основные этапы возникновения и развития

каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Содержание:
Репарация повреждений ДНК.
Генетическая рекомбинация.
Экспрессионный
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Содержание:
Репарация повреждений ДНК.
Генетическая рекомбинация.
Экспрессионный

каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Лекция 1.
Основные этапы возникновения и
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Лекция 1.
Основные этапы возникновения и
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Мир нуклеиновых кислот
К содержанию
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Мир нуклеиновых кислот
К содержанию

каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Информационная связь между ДНК, РНК
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Информационная связь между ДНК, РНК
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
К содержанию
Опыты Эвери
Доказательство того, что
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
К содержанию
Опыты Эвери
Доказательство того, что
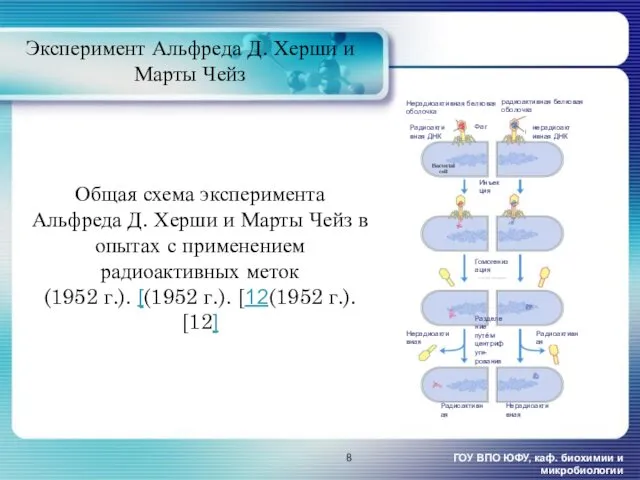
Общая схема эксперимента Альфреда Д. Херши и Марты Чейз в опытах
Общая схема эксперимента Альфреда Д. Херши и Марты Чейз в опытах
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Первичная структура нуклеиновых кислот
К содержанию
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Первичная структура нуклеиновых кислот
К содержанию
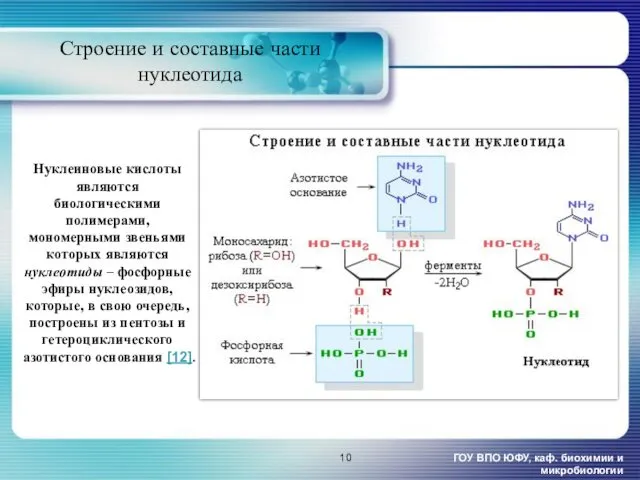
Нуклеиновые кислоты являются биологическими полимерами, мономерными звеньями которых являются нуклеотиды –
Нуклеиновые кислоты являются биологическими полимерами, мономерными звеньями которых являются нуклеотиды –
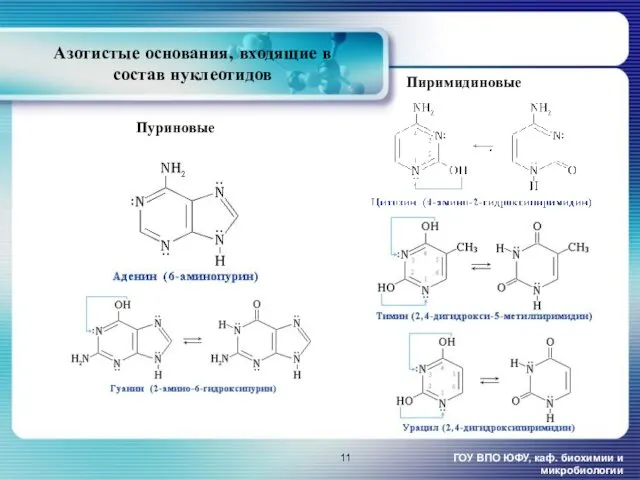
Азотистые основания, входящие в состав нуклеотидов
ГОУ ВПО ЮФУ, каф. биохимии и
Азотистые основания, входящие в состав нуклеотидов
ГОУ ВПО ЮФУ, каф. биохимии и
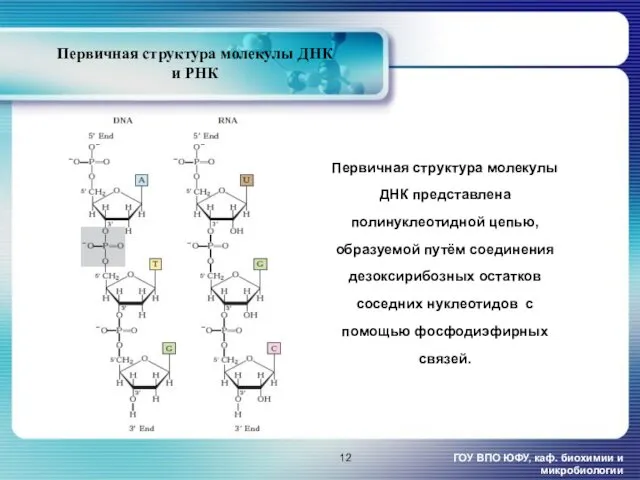
Первичная структура молекулы ДНК и РНК
ГОУ ВПО ЮФУ, каф. биохимии и
Первичная структура молекулы ДНК и РНК
ГОУ ВПО ЮФУ, каф. биохимии и
![Взаимодействия между гетероциклическими основаниями в нуклеиновых кислотах [12]. ГОУ ВПО](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-12.jpg)
Взаимодействия между гетероциклическими основаниями в нуклеиновых кислотах [12].
ГОУ ВПО ЮФУ, каф.
Взаимодействия между гетероциклическими основаниями в нуклеиновых кислотах [12].
ГОУ ВПО ЮФУ, каф.

каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Методы определения первичной
структуры ДНК.
Метод
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Методы определения первичной структуры ДНК. Метод
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Секвенирование ДНК по методу Сэнгера
(общий
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Секвенирование ДНК по методу Сэнгера (общий
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
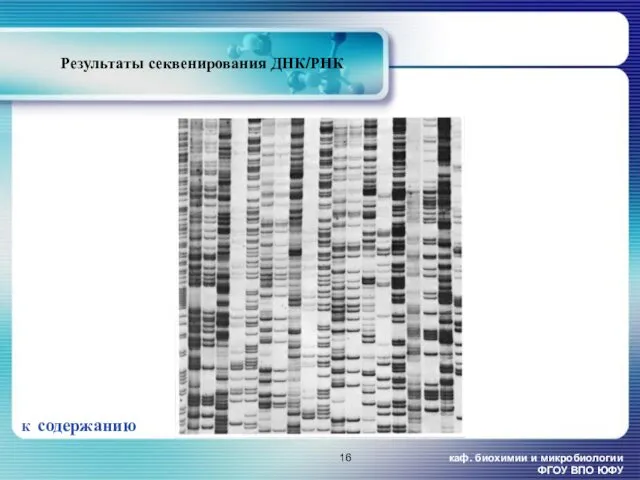
Результаты секвенирования ДНК/РНК
К содержанию
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Результаты секвенирования ДНК/РНК
К содержанию
![Физико-химические свойства ДНК. Денатурация и ренатурация ДНК [2]. ГОУ ВПО](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-16.jpg)
Физико-химические свойства ДНК.
Денатурация и ренатурация ДНК [2].
ГОУ ВПО ЮФУ, каф.
Физико-химические свойства ДНК.
Денатурация и ренатурация ДНК [2].
ГОУ ВПО ЮФУ, каф.

каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Лекция 2.
Молекулярная организация генетического материала
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Лекция 2.
Молекулярная организация генетического материала

Постулирование модели двойной спирали ДНК американским генетиком Джеймсом Уотсоном и английским
Постулирование модели двойной спирали ДНК американским генетиком Джеймсом Уотсоном и английским

Дж. Уотсон и Ф. Крик у одной из своих моделей ДНК
Дж. Уотсон и Ф. Крик у одной из своих моделей ДНК

Идеализированная картина дифракции ДНК. Результаты этих измерений привели к построению модели
Идеализированная картина дифракции ДНК. Результаты этих измерений привели к построению модели
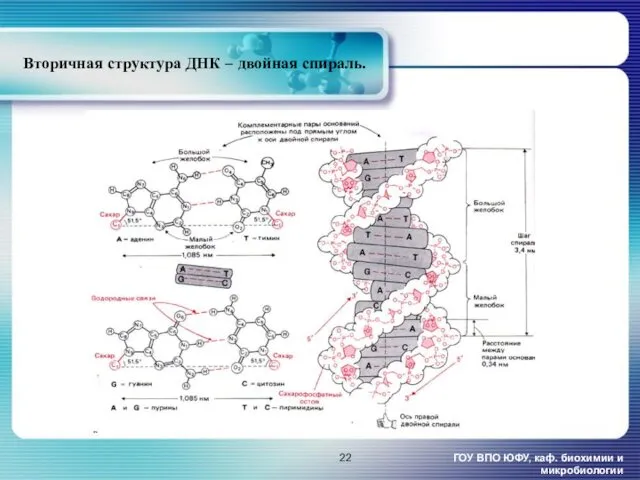
Вторичная структура ДНК – двойная спираль.
ГОУ ВПО ЮФУ, каф. биохимии и
Вторичная структура ДНК – двойная спираль.
ГОУ ВПО ЮФУ, каф. биохимии и
![Спиральная структура молекулы ДНК [2]. ГОУ ВПО ЮФУ, каф. биохимии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-22.jpg)
Спиральная структура молекулы ДНК [2].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Молекула
Спиральная структура молекулы ДНК [2].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Молекула
![Полиморфизм двойной спирали ДНК [1, 7]. ГОУ ВПО ЮФУ, каф.](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-23.jpg)
Полиморфизм двойной спирали ДНК [1, 7].
ГОУ ВПО ЮФУ, каф. биохимии и
Полиморфизм двойной спирали ДНК [1, 7].
ГОУ ВПО ЮФУ, каф. биохимии и
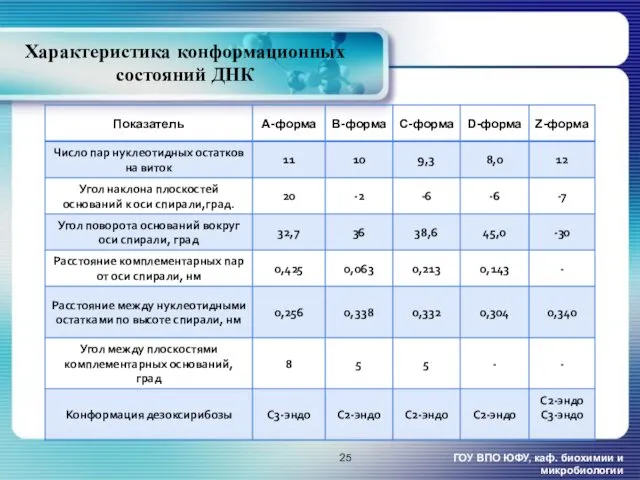
Характеристика конформационных состояний ДНК
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Характеристика конформационных состояний ДНК
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
![Упаковка ДНК. Гистоны и нуклеосомы [7, 10]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-25.jpg)
Упаковка ДНК. Гистоны и нуклеосомы [7, 10].
ГОУ ВПО ЮФУ, каф.
Упаковка ДНК. Гистоны и нуклеосомы [7, 10].
ГОУ ВПО ЮФУ, каф.

Характеристика гистонов, входящих в состав гистонового октамера
ГОУ ВПО ЮФУ, каф. биохимии
Характеристика гистонов, входящих в состав гистонового октамера
ГОУ ВПО ЮФУ, каф. биохимии
![Уровни компактизации молекулы ДНК [4]. ГОУ ВПО ЮФУ, каф. биохимии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-27.jpg)
Уровни компактизации молекулы ДНК [4].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Свободная
Уровни компактизации молекулы ДНК [4].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Свободная
![Разнообразие форм ДНК [2]. ГОУ ВПО ЮФУ, каф. биохимии и](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-28.jpg)
Разнообразие форм ДНК [2].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Линейная двухцепочечная
Разнообразие форм ДНК [2].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Линейная двухцепочечная
![ДНК кинетопласта из Leishmania tarentolae [2]. ГОУ ВПО ЮФУ, каф.](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-29.jpg)
ДНК кинетопласта из Leishmania tarentolae [2].
ГОУ ВПО ЮФУ, каф. биохимии и
ДНК кинетопласта из Leishmania tarentolae [2].
ГОУ ВПО ЮФУ, каф. биохимии и
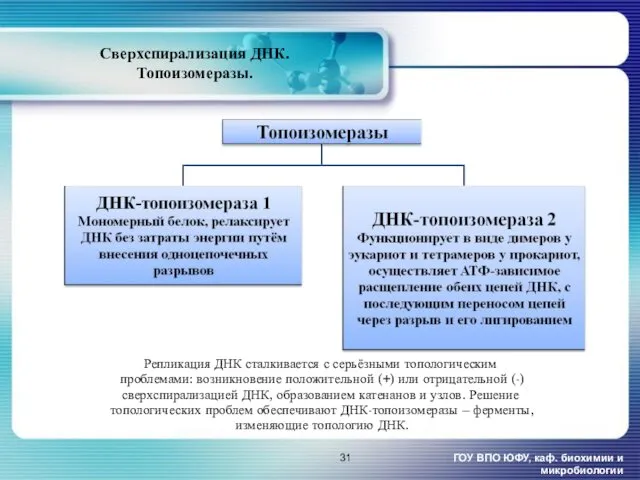
Сверхспирализация ДНК.
Топоизомеразы.
Репликация ДНК сталкивается с серьёзными топологическим
проблемами: возникновение положительной
Сверхспирализация ДНК.
Топоизомеразы.
Репликация ДНК сталкивается с серьёзными топологическим
проблемами: возникновение положительной
![Реакции, катализируемые топоизомеразами типа 1 [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-31.jpg)
Реакции, катализируемые
топоизомеразами типа 1 [2].
ГОУ ВПО ЮФУ, каф. биохимии и
Реакции, катализируемые
топоизомеразами типа 1 [2].
ГОУ ВПО ЮФУ, каф. биохимии и

Реакции, катализируемые топоизомеразами типа 2
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Топоизомеразы
Реакции, катализируемые топоизомеразами типа 2
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Топоизомеразы

Работа ДНК-гиразы
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
В бактериях обнаружена ДНК-гираза.
Работа ДНК-гиразы
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
В бактериях обнаружена ДНК-гираза.
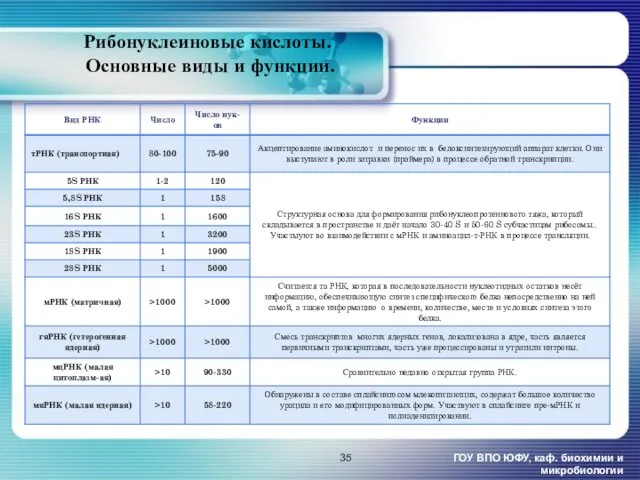
Рибонуклеиновые кислоты.
Основные виды и функции.
ГОУ ВПО ЮФУ, каф. биохимии и
Рибонуклеиновые кислоты.
Основные виды и функции.
ГОУ ВПО ЮФУ, каф. биохимии и
![Сравнительная структура ДНК и РНК [5]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-35.jpg)
Сравнительная структура ДНК и РНК [5].
ГОУ ВПО ЮФУ, каф. биохимии и
Сравнительная структура ДНК и РНК [5].
ГОУ ВПО ЮФУ, каф. биохимии и
![Вторичная и третичная структура однотяжевых РНК [2]. ГОУ ВПО ЮФУ,](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-36.jpg)
Вторичная и третичная структура однотяжевых РНК [2].
ГОУ ВПО ЮФУ, каф. биохимии
Вторичная и третичная структура однотяжевых РНК [2].
ГОУ ВПО ЮФУ, каф. биохимии

Типы «третичных» внутримолекулярных взаимодействий. между отдельными элементами вторичной структуры молекулы РНК
Типы «третичных» внутримолекулярных взаимодействий. между отдельными элементами вторичной структуры молекулы РНК
![Структура тРНК [12]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-38.jpg)
Структура тРНК [12].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Последовательность тРНК включает
Структура тРНК [12].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Последовательность тРНК включает

каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Лекция 3.
Мозаичная организация генома эукариот
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Лекция 3.
Мозаичная организация генома эукариот
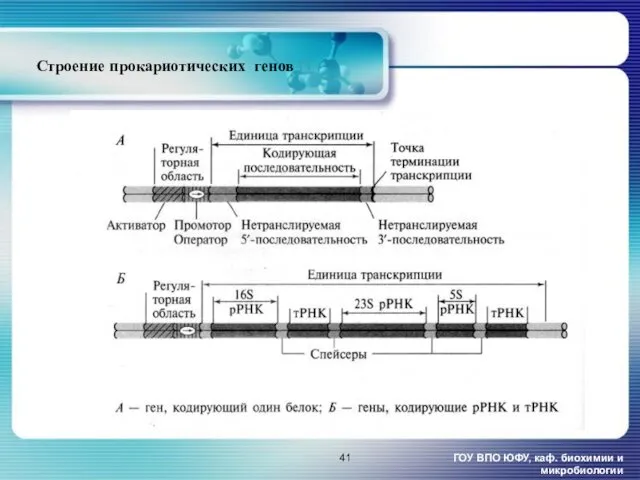
Строение прокариотических генов [1
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Строение прокариотических генов [1
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
![Оперонная организация генов прокариот [1]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-41.jpg)
Оперонная организация генов прокариот [1].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Оперонная организация генов прокариот [1].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии

Гетеродуплексы ДНК - мРНК
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Показаны петли,
Гетеродуплексы ДНК - мРНК
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Показаны петли,

Организация прерывистого гена
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Организация прерывистого гена
Организация прерывистого гена
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Организация прерывистого гена

Структура β – глобинового гена и созревание его транскрипта
ГОУ ВПО ЮФУ,
Структура β – глобинового гена и созревание его транскрипта
ГОУ ВПО ЮФУ,
![ГОУ ВПО ЮФУ, каф. биохимии и микробиологии Строение эукариотического гена, кодирующего белок [1].](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-45.jpg)
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Строение эукариотического гена, кодирующего белок
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Строение эукариотического гена, кодирующего белок
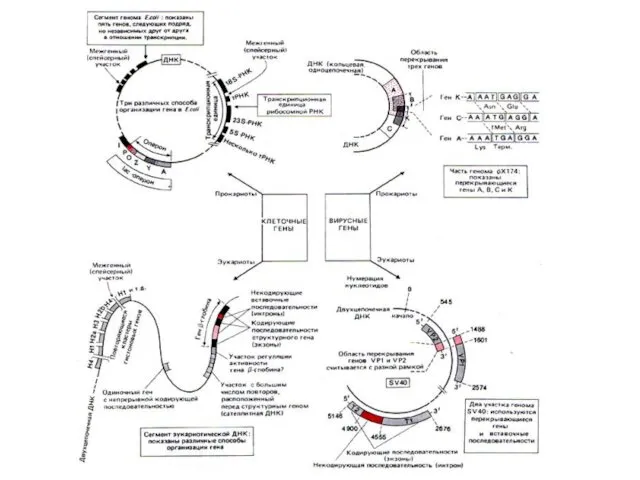
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Организация генетического материала различных форм
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Организация генетического материала различных форм

ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Передача генетического материала в клетке
Процесс
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Передача генетического материала в клетке
Процесс
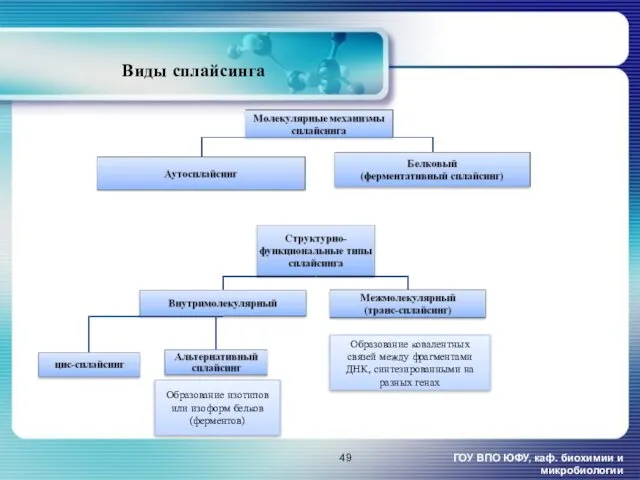
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Виды сплайсинга
Образование ковалентных связей между
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Виды сплайсинга
Образование ковалентных связей между

ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Аутосплайсинг 26 S рРНК
у
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Аутосплайсинг 26 S рРНК у
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Сплайсинг предшественника мРНК с участием
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Сплайсинг предшественника мРНК с участием

Механизмы ферментативного сплайсинга ядерных генов животных.
ГОУ ВПО ЮФУ, каф. биохимии и
Механизмы ферментативного сплайсинга ядерных генов животных.
ГОУ ВПО ЮФУ, каф. биохимии и

Альтернативный сплайсинг при экспрессии гена кальцитонина крысы.
ГОУ ВПО ЮФУ, каф. биохимии
Альтернативный сплайсинг при экспрессии гена кальцитонина крысы.
ГОУ ВПО ЮФУ, каф. биохимии
![Типы альтернативного сплайсинга пре-мРНК [1]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-53.jpg)
Типы альтернативного сплайсинга
пре-мРНК [1].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Типы альтернативного сплайсинга
пре-мРНК [1].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии

Процессинг тРНК.
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Процессинг тРНК из молекулы-предшественника
Процессинг тРНК.
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Процессинг тРНК из молекулы-предшественника
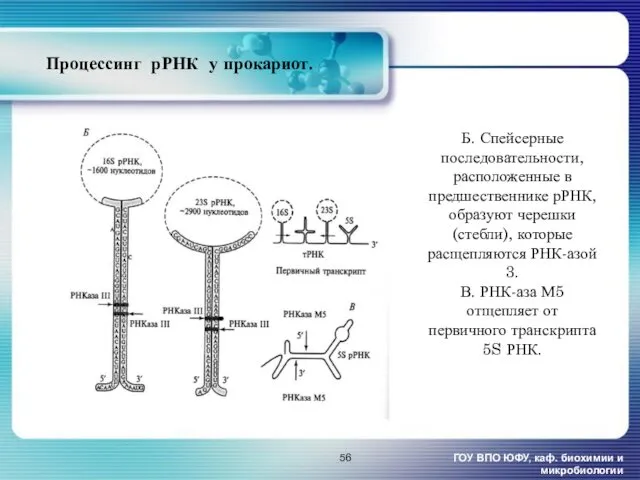
Процессинг рРНК у прокариот.
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Б. Спейсерные
Процессинг рРНК у прокариот.
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Б. Спейсерные

каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Лекция 4.
Нестабильность генома.
Мобильные генетические
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Лекция 4.
Нестабильность генома.
Мобильные генетические
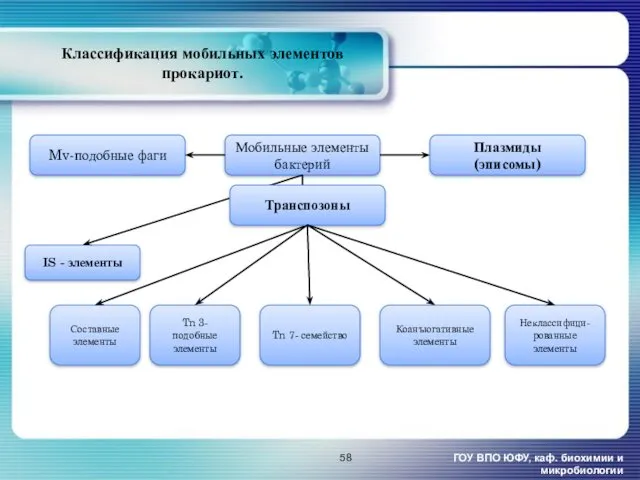
Классификация мобильных элементов прокариот.
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Транспозоны
Tn 3-
Классификация мобильных элементов прокариот.
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Транспозоны
Tn 3-
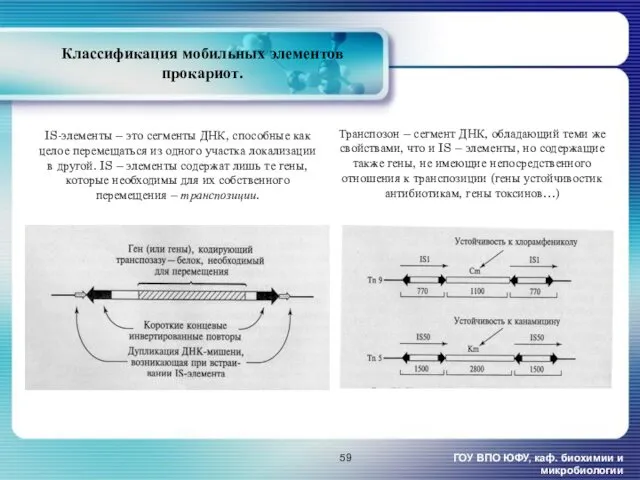
Классификация мобильных элементов прокариот.
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
IS-элементы –
Классификация мобильных элементов прокариот.
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
IS-элементы –
![Типичные IS-элементы E. coli [1] ГОУ ВПО ЮФУ, каф. биохимии и микробиологии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-59.jpg)
Типичные IS-элементы E. coli [1]
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Типичные IS-элементы E. coli [1]
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии

Принципиальное строение составного транспозона Tn10.
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Принципиальное строение составного транспозона Tn10.
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
![R-плазмида со всеми инсерционными последовательностями (IS) и транспозонами (Tn) [2]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-61.jpg)
R-плазмида со всеми инсерционными
последовательностями (IS) и транспозонами (Tn) [2]
ГОУ ВПО
R-плазмида со всеми инсерционными
последовательностями (IS) и транспозонами (Tn) [2]
ГОУ ВПО
![Механизмы перемещения мобильных элементов бактерий [1]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-62.jpg)
Механизмы перемещения
мобильных элементов бактерий [1]
Механизмы перемещения
мобильных элементов бактерий [1]

Механизм транспозиции Tn
Механизм транспозиции Tn

Расщепление обеих цепей эндонуклеазой. Места расщеплений отстоят друг от друга на
Расщепление обеих цепей эндонуклеазой. Места расщеплений отстоят друг от друга на
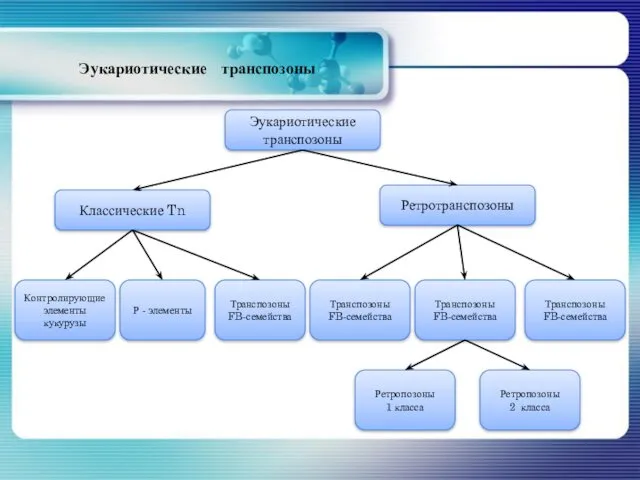
Эукариотические транспозоны
Эукариотические транспозоны
![Ретротранспозон класса 1 и его транскрипт [2]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-66.jpg)
Ретротранспозон класса 1 и его транскрипт [2]
Ретротранспозон класса 1 и его транскрипт [2]
![Модель транспозиции ретрогенов при участии РНК-посредника [2] В результате транскрипции](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-67.jpg)
Модель транспозиции ретрогенов при участии РНК-посредника [2]
В результате транскрипции и
Модель транспозиции ретрогенов при участии РНК-посредника [2]
В результате транскрипции и
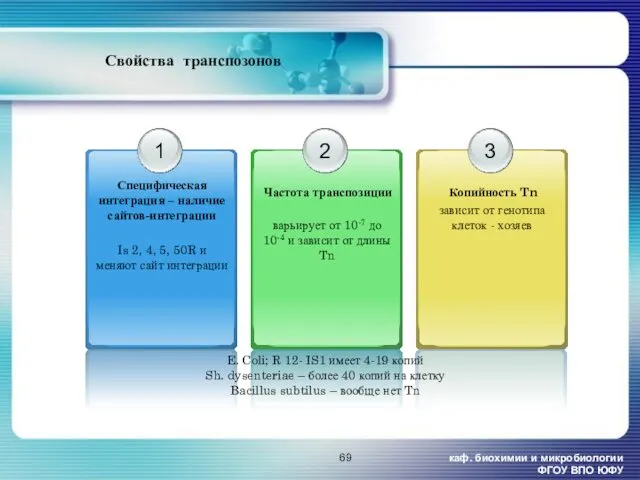
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Свойства транспозонов
варьирует от 10-7
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Свойства транспозонов
варьирует от 10-7

Биологическая роль транспозонов
Биологическая роль транспозонов

каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Лекция 5.
Репликация ДНК.
Особенности биосинтеза
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Лекция 5.
Репликация ДНК.
Особенности биосинтеза
![Предполагаемые модели репликации дуплексной ДНК [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии консервативный полуконсервативный дисперсионный](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-71.jpg)
Предполагаемые модели
репликации дуплексной ДНК [2].
ГОУ ВПО ЮФУ, каф. биохимии и
Предполагаемые модели
репликации дуплексной ДНК [2].
ГОУ ВПО ЮФУ, каф. биохимии и

Экспериментальные подтверждения
модели Уотсона-Крика
в работах Мэтью Мезельсона
и Франклина Сталя
Экспериментальные подтверждения модели Уотсона-Крика в работах Мэтью Мезельсона и Франклина Сталя

ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
А. Распределение плотности молекул ДНК,
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
А. Распределение плотности молекул ДНК,
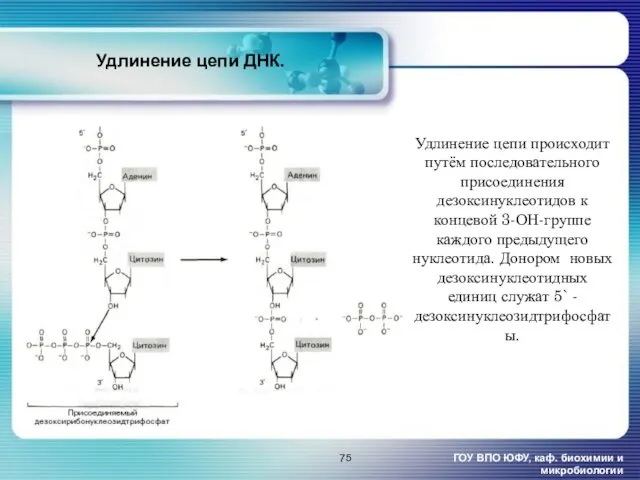
Удлинение цепи ДНК.
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Удлинение цепи происходит
Удлинение цепи ДНК.
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Удлинение цепи происходит

ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
ДНК - полимеразы.
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
ДНК - полимеразы.

ДНК-полимеразы
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
ДНК-полимераза — фермент, участвующий в
ДНК-полимеразы
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
ДНК-полимераза — фермент, участвующий в
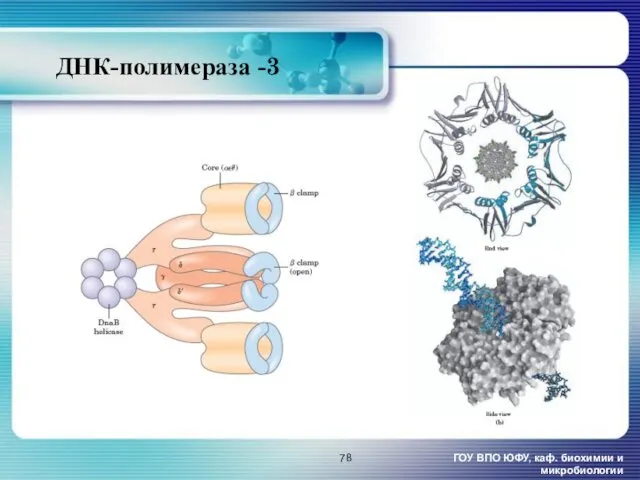
ДНК-полимераза -3
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
ДНК-полимераза -3
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
![Белки, входящие в состав репликативного комплекса ]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-78.jpg)
Белки, входящие в состав
репликативного комплекса ].
ГОУ ВПО ЮФУ, каф. биохимии
Белки, входящие в состав
репликативного комплекса ].
ГОУ ВПО ЮФУ, каф. биохимии
![Инициация репликации ДНК [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-79.jpg)
Инициация репликации ДНК [2].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Инициация репликации ДНК [2].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
![Полуконсервативный механизм репликации ДНК E. coli [9]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-80.jpg)
Полуконсервативный механизм репликации ДНК E. coli [9].
ГОУ ВПО ЮФУ, каф. биохимии
Полуконсервативный механизм репликации ДНК E. coli [9].
ГОУ ВПО ЮФУ, каф. биохимии
![Репликация ДНК [Репликация ДНК [ 7Репликация ДНК [ 7]. ГОУ](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-81.jpg)
Репликация ДНК [Репликация ДНК [ 7Репликация ДНК [ 7].
ГОУ ВПО
Репликация ДНК [Репликация ДНК [ 7Репликация ДНК [ 7].
ГОУ ВПО
![ДНК-лигаза. Механизм лигазной реакции [12]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-82.jpg)
ДНК-лигаза. Механизм лигазной реакции [12].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
ДНК-лигаза. Механизм лигазной реакции [12].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии

ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Репликативные механизмы различных форм живой
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Репликативные механизмы различных форм живой
![Репликация хромосомной ДНК эукариот [2]. ГОУ ВПО ЮФУ, каф. биохимии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-84.jpg)
Репликация хромосомной ДНК эукариот [2].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Репликация
Репликация хромосомной ДНК эукариот [2].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Репликация
![Начало репликации [2]. Репликация инициируется в специфическом участае ДНК, точке](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-85.jpg)
Начало репликации [2].
Репликация инициируется в специфическом участае ДНК, точке
начала
Начало репликации [2].
Репликация инициируется в специфическом участае ДНК, точке
начала
![Репликация по типу катящегося кольца [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-86.jpg)
Репликация по типу катящегося кольца [2].
ГОУ ВПО ЮФУ, каф. биохимии и
Репликация по типу катящегося кольца [2].
ГОУ ВПО ЮФУ, каф. биохимии и
![Поочерёдная репликация цепей ДНК аденовируса [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-87.jpg)
Поочерёдная репликация
цепей ДНК аденовируса [2].
ГОУ ВПО ЮФУ, каф. биохимии
Поочерёдная репликация
цепей ДНК аденовируса [2].
ГОУ ВПО ЮФУ, каф. биохимии
![Репликация теломерных участков эукариотических хромосом [1]. ГОУ ВПО ЮФУ, каф.](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-88.jpg)
Репликация теломерных
участков эукариотических хромосом [1].
ГОУ ВПО ЮФУ, каф. биохимии и
Репликация теломерных
участков эукариотических хромосом [1].
ГОУ ВПО ЮФУ, каф. биохимии и

ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Общий принцип полимеразной цепной реакции.
Денатурация
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Общий принцип полимеразной цепной реакции.
Денатурация
![Сравнительная структура ДНК и РНК [5]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-90.jpg)
Сравнительная структура ДНК и РНК [5].
ГОУ ВПО ЮФУ, каф. биохимии и
Сравнительная структура ДНК и РНК [5].
ГОУ ВПО ЮФУ, каф. биохимии и

каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Лекция 6.
Репарация повреждений ДНК.
К
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Лекция 6.
Репарация повреждений ДНК.
К
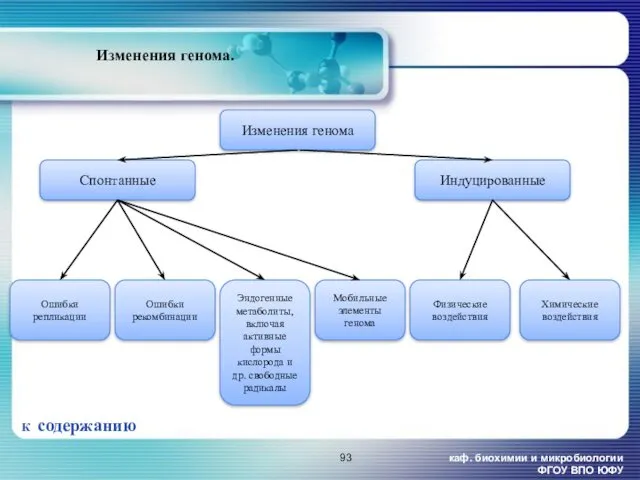
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Изменения генома.
К содержанию
Мобильные элементы генома
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Изменения генома.
К содержанию
Мобильные элементы генома
![Мутагенные агенты и типы повреждений молекулы ДНК [4]. ФГОУ ВПО](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-93.jpg)
Мутагенные агенты и типы
повреждений молекулы ДНК [4].
ФГОУ ВПО ЮФУ, каф.
Мутагенные агенты и типы
повреждений молекулы ДНК [4].
ФГОУ ВПО ЮФУ, каф.
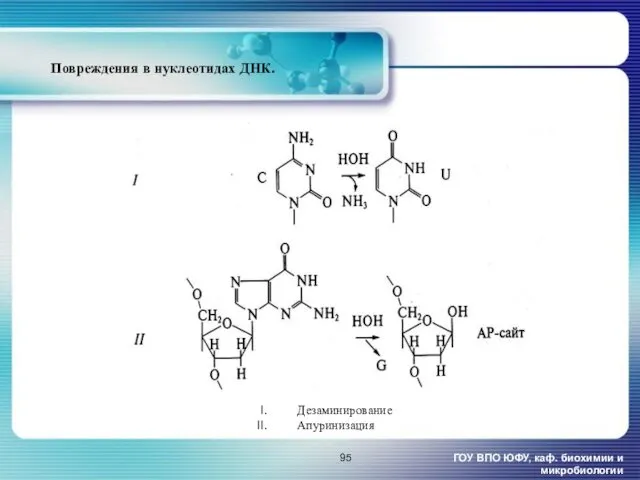
Повреждения в нуклеотидах ДНК.
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Дезаминирование
Апуринизация
Повреждения в нуклеотидах ДНК.
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Дезаминирование
Апуринизация
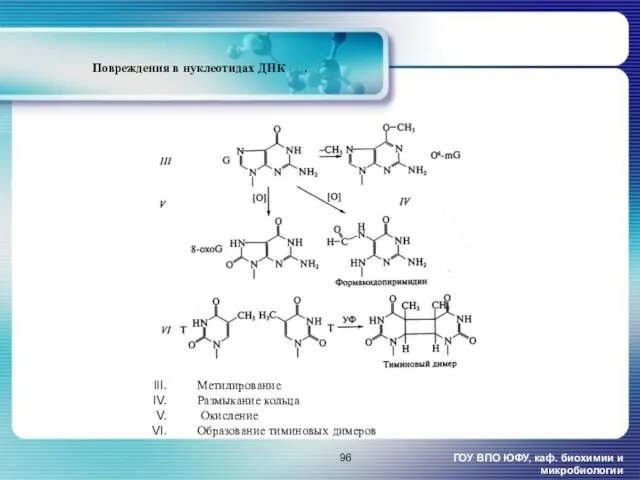
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Повреждения в нуклеотидах ДНК [1].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Повреждения в нуклеотидах ДНК [1].
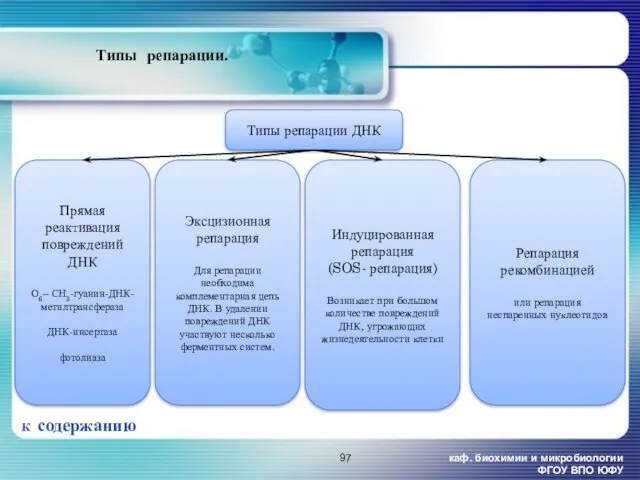
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Типы репарации.
К содержанию
Эксцизионная репарация
Для репарации
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Типы репарации.
К содержанию
Эксцизионная репарация
Для репарации
![Прямая репарация [1] ГОУ ВПО ЮФУ, каф. биохимии и микробиологии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-97.jpg)
Прямая репарация [1]
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Дезалкилирование
О6-метилгуаниновых
остатков
Прямая репарация [1]
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Дезалкилирование
О6-метилгуаниновых
остатков
![Схема эксцизионной репарации [1]. ГОУ ВПО ЮФУ, каф. биохимии и](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-98.jpg)
Схема эксцизионной репарации [1].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
ДНК-N-гликозилаза удаляет
Схема эксцизионной репарации [1].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
ДНК-N-гликозилаза удаляет
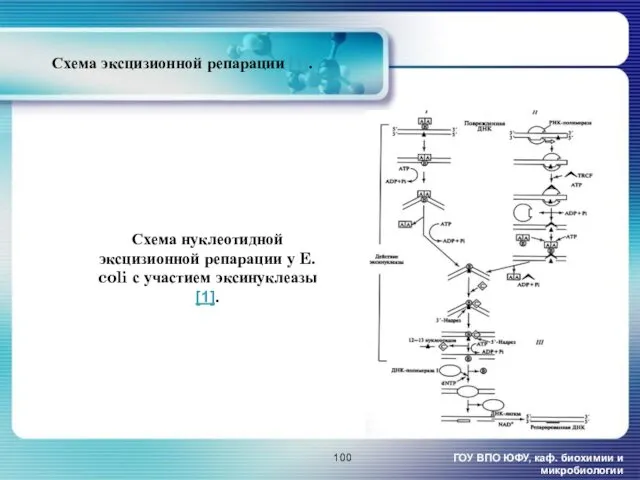
Схема нуклеотидной эксцизионной репарации у E. coli с участием эксинуклеазы [1].
ГОУ
Схема нуклеотидной эксцизионной репарации у E. coli с участием эксинуклеазы [1].
ГОУ
![Ферментативная репарация циклобутановых тиминовых димеров [2]. ГОУ ВПО ЮФУ, каф.](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-100.jpg)
Ферментативная репарация
циклобутановых тиминовых димеров [2].
ГОУ ВПО ЮФУ, каф. биохимии и
Ферментативная репарация
циклобутановых тиминовых димеров [2].
ГОУ ВПО ЮФУ, каф. биохимии и
![Репарация с помощью N- гликозилаз. [2] ГОУ ВПО ЮФУ, каф. биохимии и микробиологии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-101.jpg)
Репарация с помощью N- гликозилаз. [2]
ГОУ ВПО ЮФУ, каф. биохимии и
Репарация с помощью N- гликозилаз. [2]
ГОУ ВПО ЮФУ, каф. биохимии и
![Делеции и амплификации, возникающие при репликации или репарации [1]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-102.jpg)
Делеции и амплификации,
возникающие при репликации или репарации [1].
ГОУ ВПО ЮФУ,
Делеции и амплификации,
возникающие при репликации или репарации [1].
ГОУ ВПО ЮФУ,

каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Некоторые гены SOS-системы и результат
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Некоторые гены SOS-системы и результат

каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Лекция 7.
Генетическая рекомбинация.
К содержанию
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Лекция 7.
Генетическая рекомбинация.
К содержанию
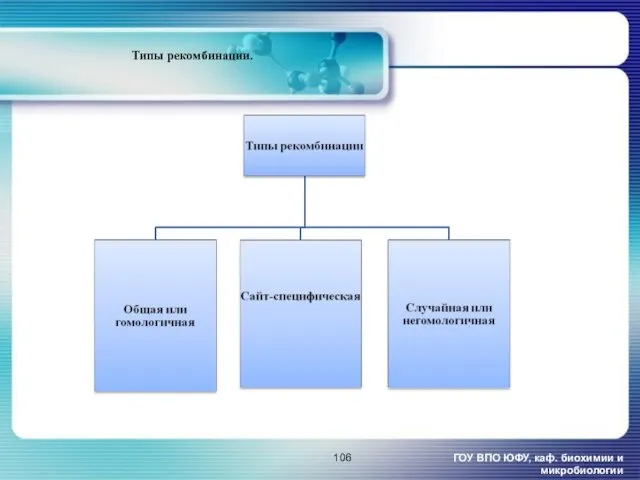
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Типы рекомбинации.
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Типы рекомбинации.
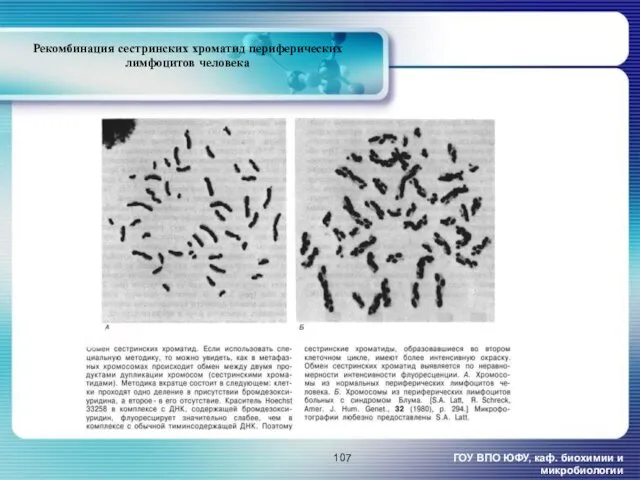
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Рекомбинация сестринских хроматид периферических лимфоцитов
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Рекомбинация сестринских хроматид периферических лимфоцитов

ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Гомологичная рекомбинация
Комплементарное спаривание цепей, принадлежащих
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Гомологичная рекомбинация
Комплементарное спаривание цепей, принадлежащих
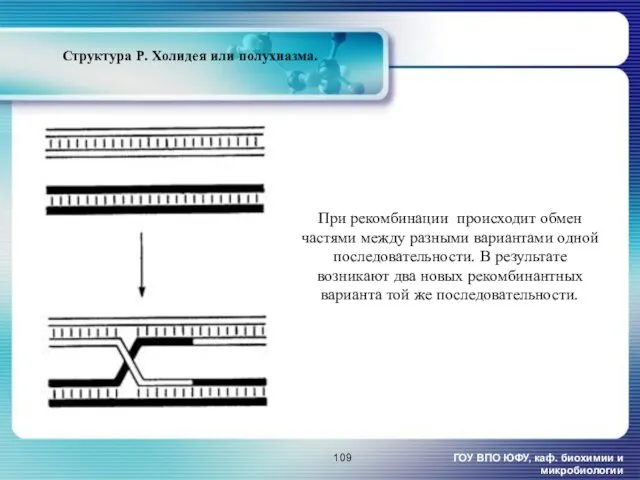
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Структура Р. Холидея или полухиазма.
При
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Структура Р. Холидея или полухиазма.
При
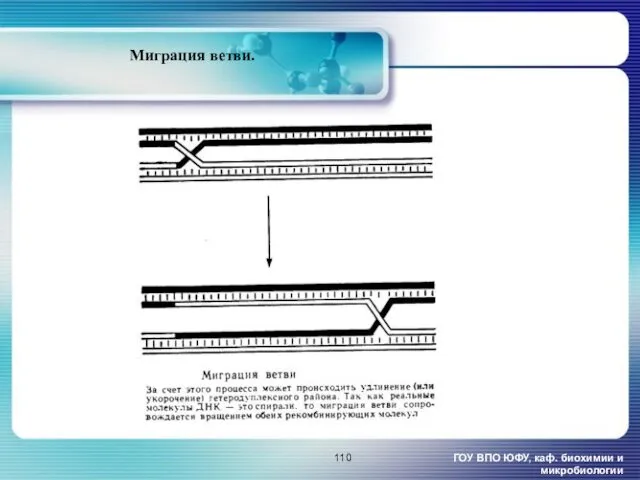
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Миграция ветви.
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Миграция ветви.

ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Изомеризация полухиазмы.
Если развернуть полухиазму в
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Изомеризация полухиазмы.
Если развернуть полухиазму в

ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Способы разрезания полухиазмы.
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Способы разрезания полухиазмы.
![Общая рекомбинация с образованием двухцепочечного разрыва [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-112.jpg)
Общая рекомбинация с образованием двухцепочечного разрыва [2].
ГОУ ВПО ЮФУ, каф. биохимии
Общая рекомбинация с образованием двухцепочечного разрыва [2].
ГОУ ВПО ЮФУ, каф. биохимии
![Сайт-специфическая рекомбинация [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-113.jpg)
Сайт-специфическая рекомбинация [2].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Специфические последовательности имеются
Сайт-специфическая рекомбинация [2].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Специфические последовательности имеются
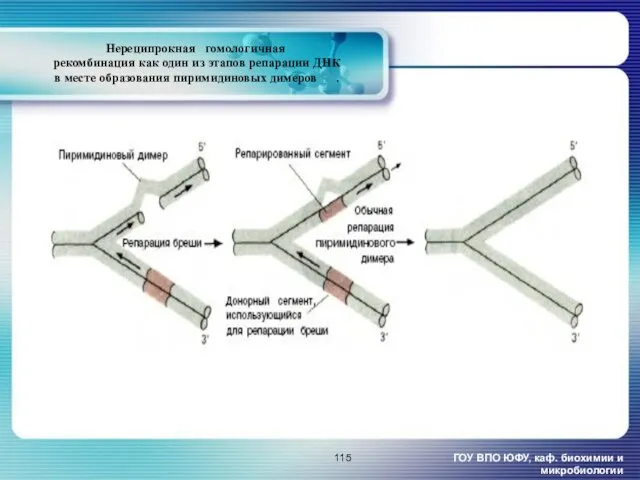
Нереципрокная гомологичная
рекомбинация как один из этапов репарации ДНК
в месте
Нереципрокная гомологичная рекомбинация как один из этапов репарации ДНК в месте
![RecA-зависимая общая рекомбинация ДНК E/ coli [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-115.jpg)
RecA-зависимая общая рекомбинация ДНК E/ coli [2].
ГОУ ВПО ЮФУ, каф. биохимии
RecA-зависимая общая рекомбинация ДНК E/ coli [2].
ГОУ ВПО ЮФУ, каф. биохимии

каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Лекция 8.
Экспрессионный аппарат клетки. Транскрипция.
К
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Лекция 8.
Экспрессионный аппарат клетки. Транскрипция.
К
![Пространственная модель комплекса РНК-полимеразы E. coli с фрагментом ДНК-матрицы [9].](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-117.jpg)
Пространственная модель комплекса
РНК-полимеразы E. coli
с фрагментом ДНК-матрицы [9].
ГОУ ВПО
Пространственная модель комплекса
РНК-полимеразы E. coli
с фрагментом ДНК-матрицы [9].
ГОУ ВПО
![Схема синтеза РНК с помощью РНК-полимеразы [9]. ГОУ ВПО ЮФУ,](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-118.jpg)
Схема синтеза РНК с помощью РНК-полимеразы [9].
ГОУ ВПО ЮФУ, каф. биохимии
Схема синтеза РНК с помощью РНК-полимеразы [9].
ГОУ ВПО ЮФУ, каф. биохимии
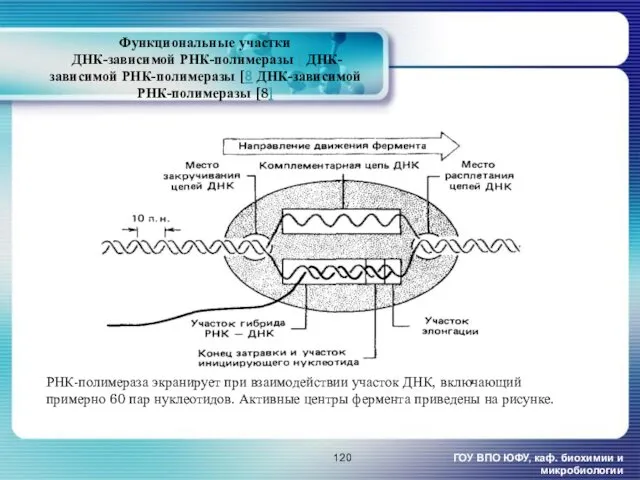
Функциональные участки
ДНК-зависимой РНК-полимеразы [ ДНК-зависимой РНК-полимеразы [8 ДНК-зависимой РНК-полимеразы [8]
ГОУ
Функциональные участки
ДНК-зависимой РНК-полимеразы [ ДНК-зависимой РНК-полимеразы [8 ДНК-зависимой РНК-полимеразы [8]
ГОУ
![Типичный промотор E. coli [2]. ГОУ ВПО ЮФУ, каф. биохимии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-120.jpg)
Типичный промотор E. coli [2].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Транскрипция
Типичный промотор E. coli [2].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Транскрипция
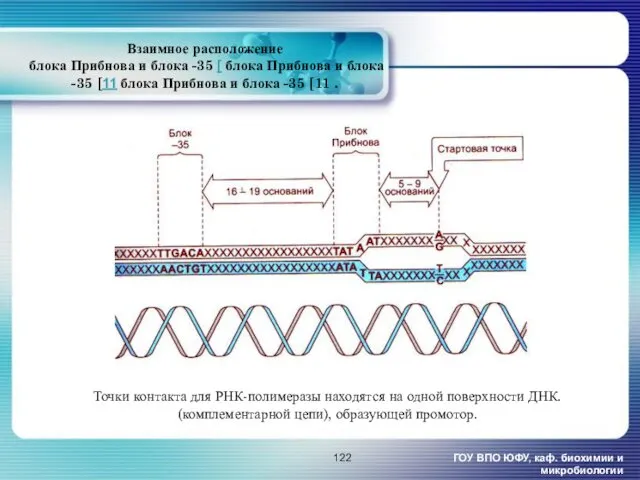
Взаимное расположение
блока Прибнова и блока -35 [ блока Прибнова и
Взаимное расположение блока Прибнова и блока -35 [ блока Прибнова и
![Очерёдность процессов транскрипции, трансляции и деградации иРНК у бактерий [8]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-122.jpg)
Очерёдность процессов транскрипции, трансляции и деградации иРНК у бактерий [8] .
ГОУ
Очерёдность процессов транскрипции, трансляции и деградации иРНК у бактерий [8] .
ГОУ
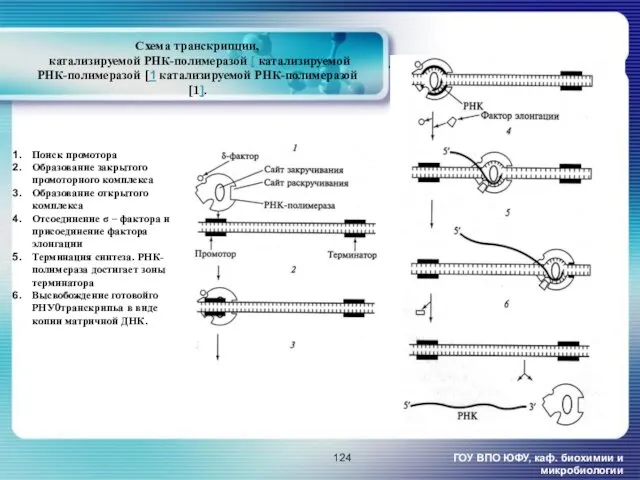
Схема транскрипции,
катализируемой РНК-полимеразой [ катализируемой РНК-полимеразой [1 катализируемой РНК-полимеразой [1].
ГОУ
Схема транскрипции,
катализируемой РНК-полимеразой [ катализируемой РНК-полимеразой [1 катализируемой РНК-полимеразой [1].
ГОУ
![Терминация транскрипции. Примеры шпилек в ρ-независимых и ρ-зависимых терминаторах [2].](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-124.jpg)
Терминация транскрипции.
Примеры шпилек в ρ-независимых
и ρ-зависимых терминаторах [2].
ГОУ ВПО
Терминация транскрипции.
Примеры шпилек в ρ-независимых
и ρ-зависимых терминаторах [2].
ГОУ ВПО

Влияние ρ-белка на остановку РНК-полимеразы в терминаторе, сопровождающуюся терминацией транскрипции с
Влияние ρ-белка на остановку РНК-полимеразы в терминаторе, сопровождающуюся терминацией транскрипции с
![Эукариотическая иРНК [Эукариотическая иРНК [11Эукариотическая иРНК [11]. ГОУ ВПО ЮФУ,](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-126.jpg)
Эукариотическая иРНК [Эукариотическая иРНК [11Эукариотическая иРНК [11].
ГОУ ВПО ЮФУ, каф. биохимии
Эукариотическая иРНК [Эукариотическая иРНК [11Эукариотическая иРНК [11].
ГОУ ВПО ЮФУ, каф. биохимии
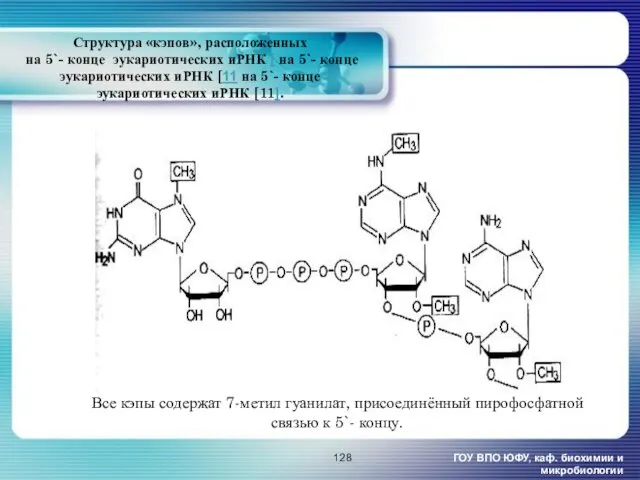
Структура «кэпов», расположенных
на 5`- конце эукариотических иРНК [ на 5`-
Структура «кэпов», расположенных на 5`- конце эукариотических иРНК [ на 5`-
![Обратная транскрипция [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-128.jpg)
Обратная транскрипция [2].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Схематическое изображение обратной
Обратная транскрипция [2].
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Схематическое изображение обратной

каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Лекция 9.
Генетический код.
Молекулярная организация
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Лекция 9.
Генетический код.
Молекулярная организация

Свойства генетического кода:
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Генетический код
Свойства генетического кода:
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Генетический код
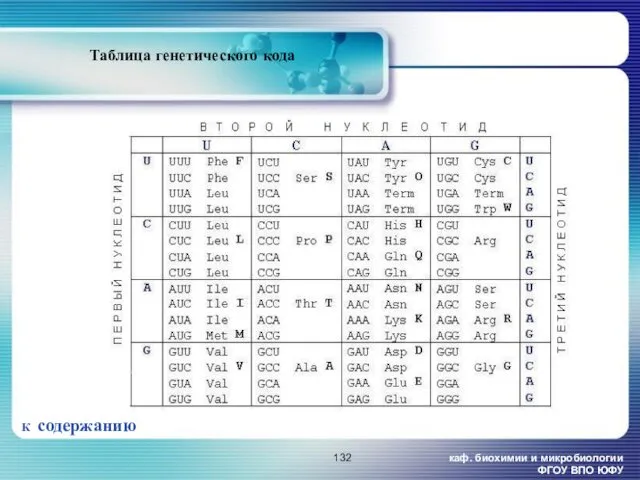
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Таблица генетического кода
К содержанию
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Таблица генетического кода
К содержанию
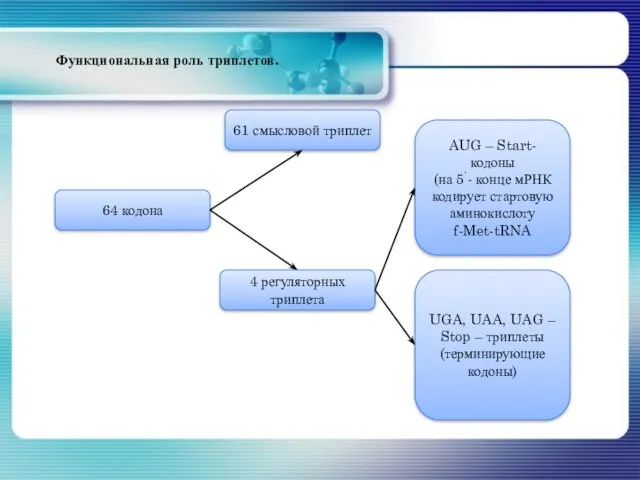
AUG – Start-кодоны
(на 5`- конце мРНК кодирует стартовую аминокислоту f-Met-tRNA
UGA,
AUG – Start-кодоны
(на 5`- конце мРНК кодирует стартовую аминокислоту f-Met-tRNA
UGA,
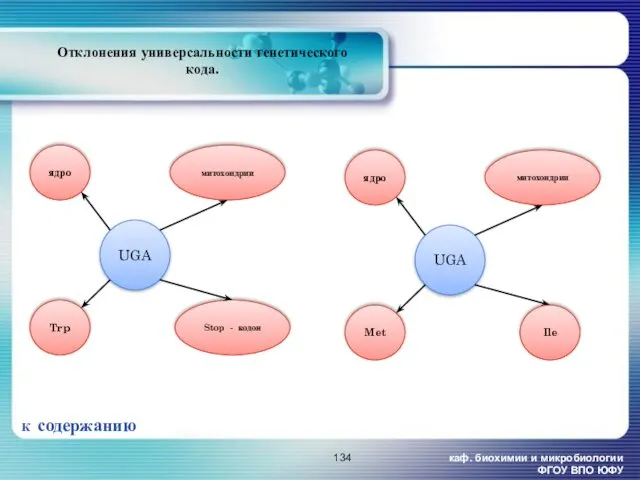
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Отклонения универсальности генетического кода.
К содержанию
Ile
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Отклонения универсальности генетического кода.
К содержанию
Ile
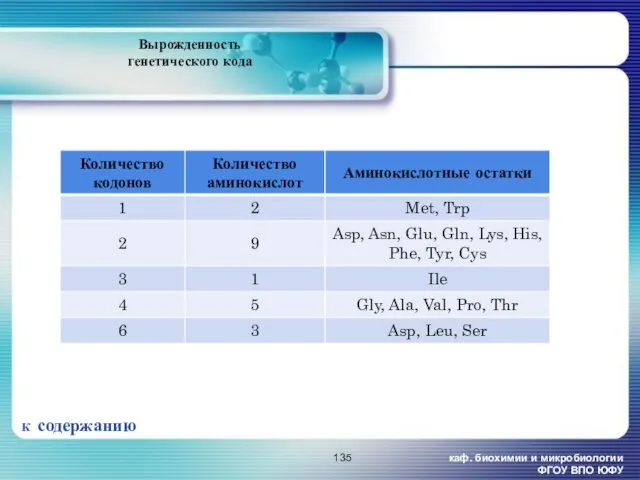
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Вырожденность генетического кода
К содержанию
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Вырожденность генетического кода
К содержанию

каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Гипотеза неоднозначного соответствия Ф. Крика
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Гипотеза неоднозначного соответствия Ф. Крика

Рамка считывания.
Поскольку каждый кодон содержит три нуклеотида, один и тот же
Рамка считывания.
Поскольку каждый кодон содержит три нуклеотида, один и тот же
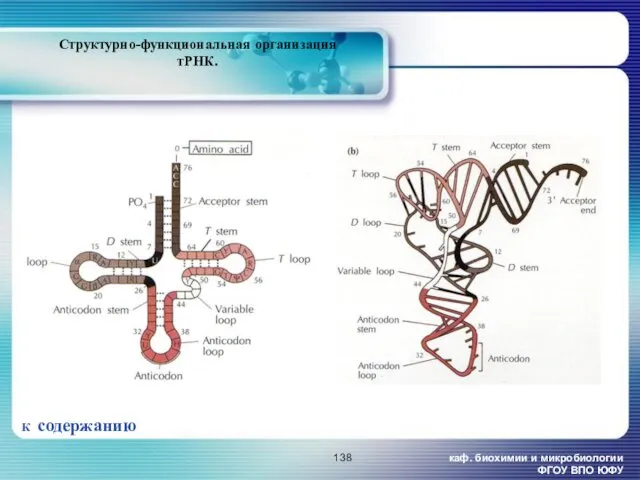
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Структурно-функциональная организация тРНК.
К содержанию
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Структурно-функциональная организация тРНК.
К содержанию
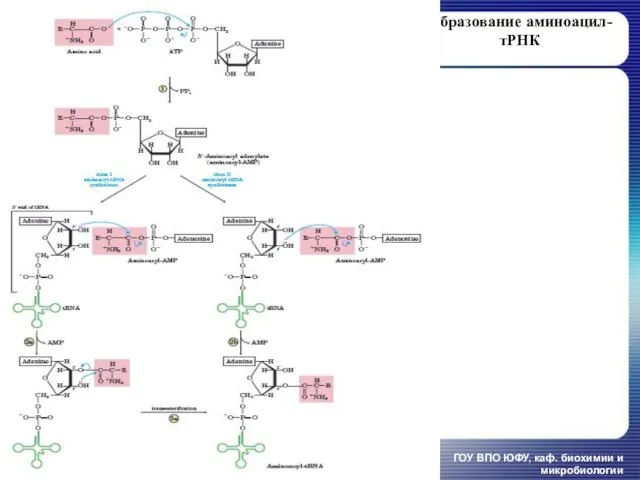
Образование аминоацил-тРНК
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Образование аминоацил-тРНК
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
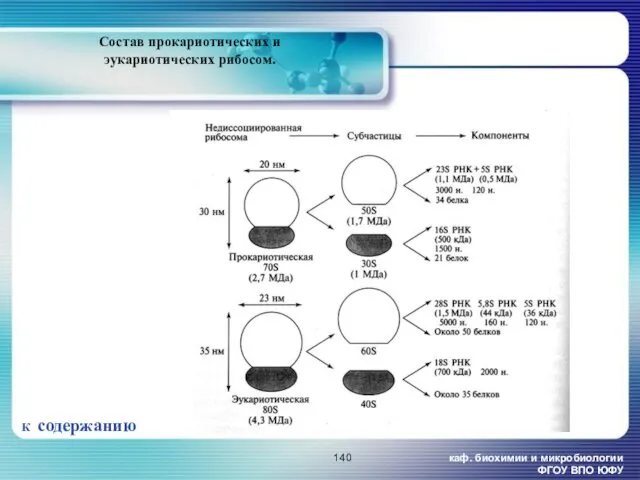
Состав прокариотических и эукариотических рибосом.
К
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Состав прокариотических и эукариотических рибосом.
К
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
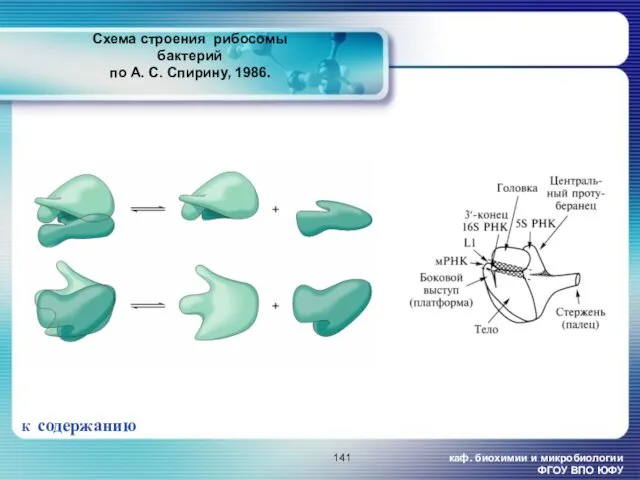
Схема строения рибосомы бактерий
по
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Схема строения рибосомы бактерий по
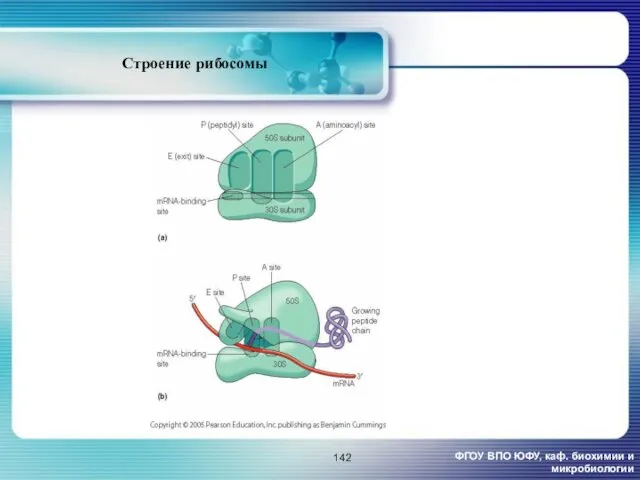
Строение рибосомы
ФГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Строение рибосомы
ФГОУ ВПО ЮФУ, каф. биохимии и микробиологии

каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Лекция 10.
Трансляция: условия и энергетика.
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Лекция 10.
Трансляция: условия и энергетика.
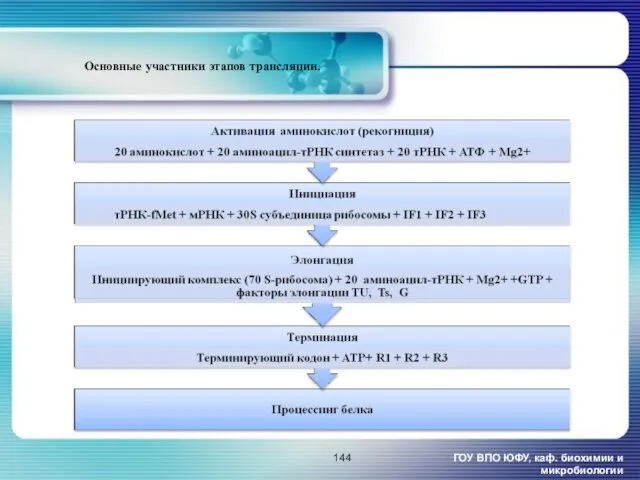
Основные участники этапов трансляции.
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии
Основные участники этапов трансляции.
ГОУ ВПО ЮФУ, каф. биохимии и микробиологии

каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Ферменты (факторы) трансляции про- и
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Ферменты (факторы) трансляции про- и
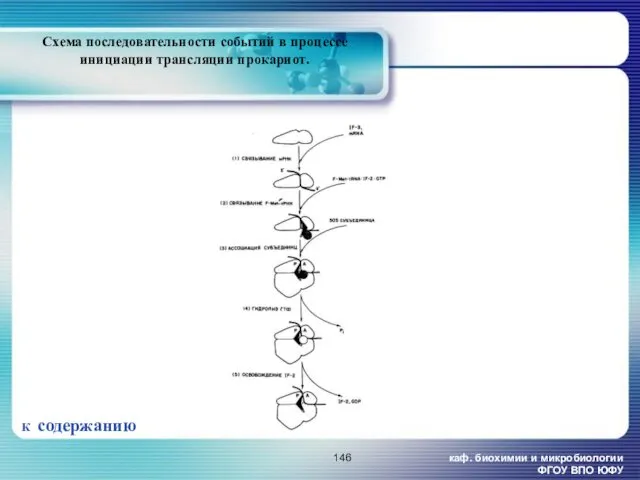
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Схема последовательности событий в процессе
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Схема последовательности событий в процессе
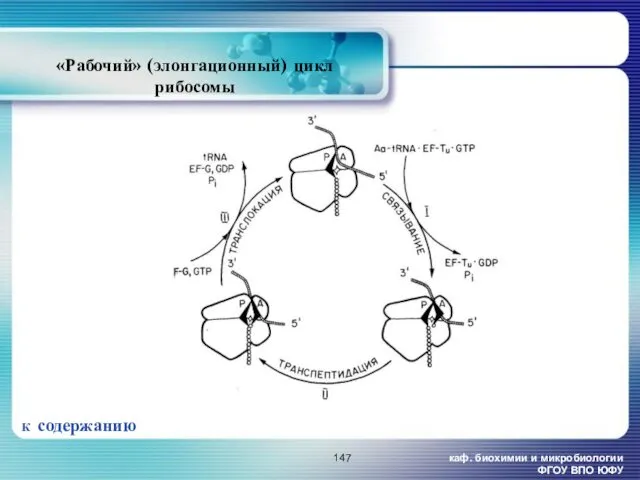
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
«Рабочий» (элонгационный) цикл рибосомы
К содержанию
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
«Рабочий» (элонгационный) цикл рибосомы
К содержанию
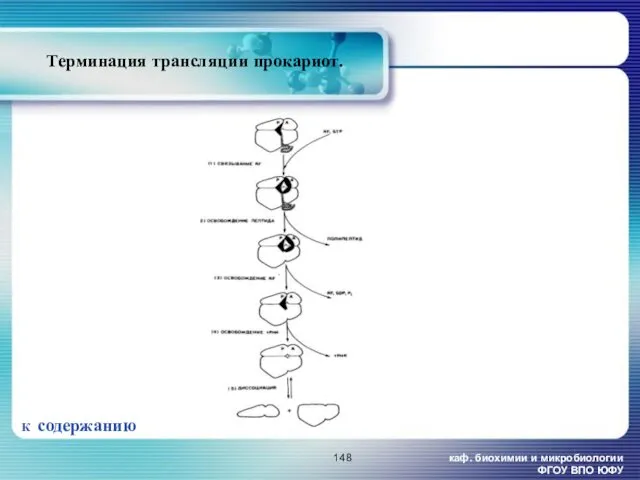
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Терминация трансляции прокариот.
К содержанию
каф. биохимии и микробиологии ФГОУ ВПО ЮФУ
Терминация трансляции прокариот.
К содержанию
![Лактозный оперон E. coli и ген lac-репрессора [2]. ГОУ ВПО ЮФУ, каф. биохимии и микробиологии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/58085/slide-148.jpg)
Лактозный оперон E. coli и ген lac-репрессора [2].
ГОУ ВПО ЮФУ, каф.
Лактозный оперон E. coli и ген lac-репрессора [2].
ГОУ ВПО ЮФУ, каф.
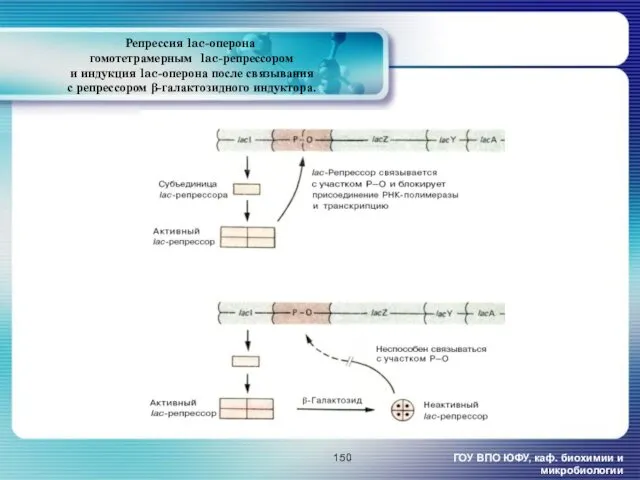
Репрессия lac-оперона
гомотетрамерным lac-репрессором
и индукция lac-оперона после связывания
с репрессором
Репрессия lac-оперона гомотетрамерным lac-репрессором и индукция lac-оперона после связывания с репрессором
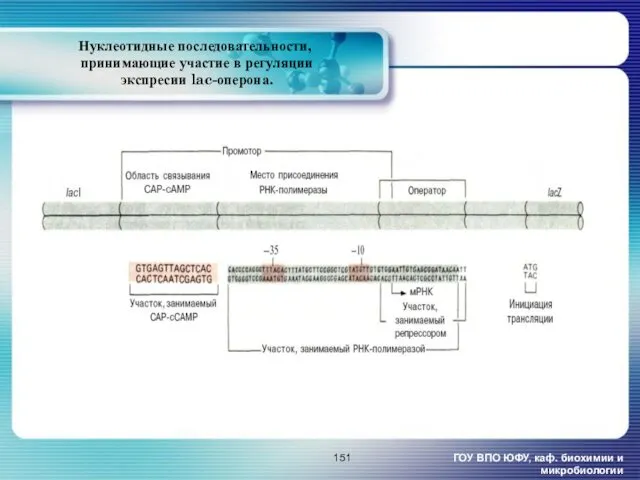
Нуклеотидные последовательности,
принимающие участие в регуляции
экспресии lac-оперона.
ГОУ ВПО ЮФУ, каф.
Нуклеотидные последовательности,
принимающие участие в регуляции
экспресии lac-оперона.
ГОУ ВПО ЮФУ, каф.
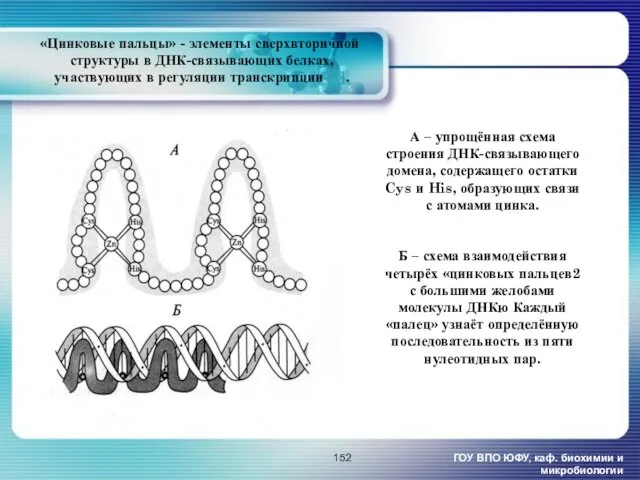
«Цинковые пальцы» - элементы сверхвторичной
структуры в ДНК-связывающих белках,
участвующих в
«Цинковые пальцы» - элементы сверхвторичной структуры в ДНК-связывающих белках, участвующих в
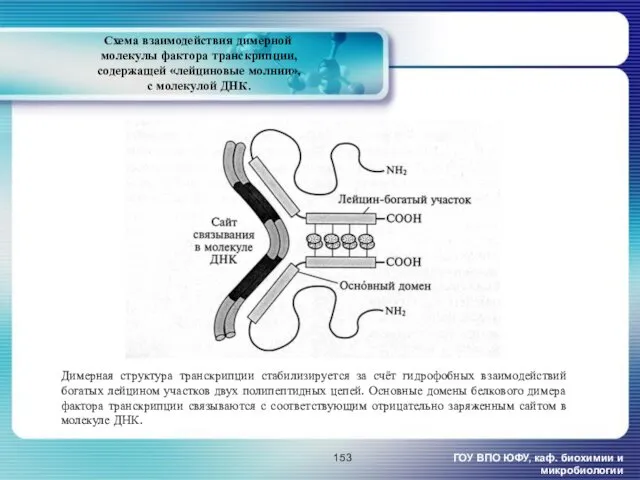
Схема взаимодействия димерной
молекулы фактора транскрипции,
содержащей «лейциновые молнии»,
с молекулой
Схема взаимодействия димерной молекулы фактора транскрипции, содержащей «лейциновые молнии», с молекулой
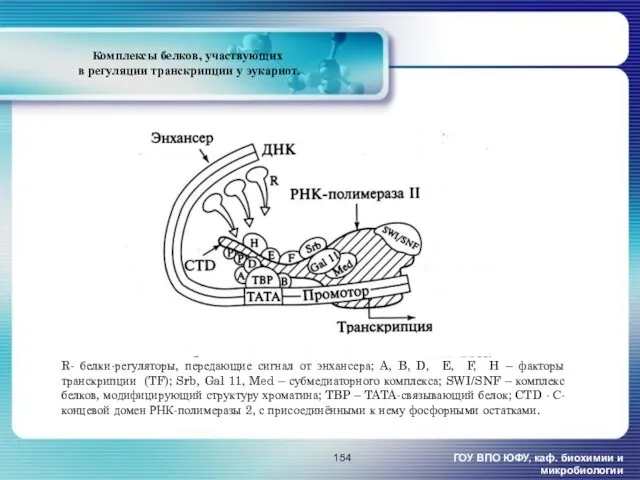
Комплексы белков, участвующих
в регуляции транскрипции у эукариот.
ГОУ ВПО ЮФУ, каф.
Комплексы белков, участвующих
в регуляции транскрипции у эукариот.
ГОУ ВПО ЮФУ, каф.

Терминация транскрипции.
Примеры шпилек в ρ-независимых
и ρ-зависимых терминаторах.
ГОУ ВПО ЮФУ,
Терминация транскрипции.
Примеры шпилек в ρ-независимых
и ρ-зависимых терминаторах.
ГОУ ВПО ЮФУ,

Влияние ρ-белка на остановку РНК-полимеразы в терминаторе, сопровождающуюся терминацией транскрипции с
Влияние ρ-белка на остановку РНК-полимеразы в терминаторе, сопровождающуюся терминацией транскрипции с

Список литературных источников.
Коничев А. С., Севастьянова Г. А. Молекулярная биология. М.:
Список литературных источников.
Коничев А. С., Севастьянова Г. А. Молекулярная биология. М.:
 Программа организации внеурочной деятельности учащихся начальной школы
Программа организации внеурочной деятельности учащихся начальной школы Диагностика, техническое обслуживание коробки перемены передач ВАЗ-2107
Диагностика, техническое обслуживание коробки перемены передач ВАЗ-2107 Тамақтану денсаулық факторы. Алиментарлық аурулар
Тамақтану денсаулық факторы. Алиментарлық аурулар Акционерное общество Всероссийский нефтегазовый научно-исследовательский институт имени академика А.П.Крылова
Акционерное общество Всероссийский нефтегазовый научно-исследовательский институт имени академика А.П.Крылова Проникаем внутрь Android. Практика
Проникаем внутрь Android. Практика Электродуговая сталеплавильная печь
Электродуговая сталеплавильная печь Мультимедийная разработка урока по предмету Музыкальный инструмент. Фортепиано.
Мультимедийная разработка урока по предмету Музыкальный инструмент. Фортепиано. презентация стили семейного воспитания работа с родителями(собрание) подготовила Ивсина Т.З. г.Армавир, МБДОУ № 33.
презентация стили семейного воспитания работа с родителями(собрание) подготовила Ивсина Т.З. г.Армавир, МБДОУ № 33. Занятие по внеурочной деятельности. Школа здоровья. Советы доктора ВОДЫ
Занятие по внеурочной деятельности. Школа здоровья. Советы доктора ВОДЫ Экономические методы государственного регулирования
Экономические методы государственного регулирования Группа компаний ОАО Интертрансэкспедиция
Группа компаний ОАО Интертрансэкспедиция Правила поведения в общественных местах
Правила поведения в общественных местах Сказочная страна. (ИЗО, 1 класс)
Сказочная страна. (ИЗО, 1 класс) Памятник Солдату и Матросу
Памятник Солдату и Матросу Перспективность социальных сетей в продвижении бизнеса
Перспективность социальных сетей в продвижении бизнеса Индивидуальный и рыночный спрос. Эластичность рыночного спроса
Индивидуальный и рыночный спрос. Эластичность рыночного спроса Презентация к коррекционно-развивающей программе Путешествие по стране Добра
Презентация к коррекционно-развивающей программе Путешествие по стране Добра Галилео Галилей
Галилео Галилей Оперативная память
Оперативная память Битва под Москвой 30 сентября 1941 г. - 20 апреля 1942 г
Битва под Москвой 30 сентября 1941 г. - 20 апреля 1942 г ФУТБОЛ
ФУТБОЛ Презентация к стихотворению Весенняя гроза Ф. Тютчева
Презентация к стихотворению Весенняя гроза Ф. Тютчева Регуляция и патология липидного обмена
Регуляция и патология липидного обмена Испытание трубопроводов на прочность и плотность
Испытание трубопроводов на прочность и плотность Свойства квадратных корней
Свойства квадратных корней Поглощения буровых и тампонажных растворов. Лекция 2
Поглощения буровых и тампонажных растворов. Лекция 2 Презентация Школьная библиотека
Презентация Школьная библиотека Тригонометрические уравнения
Тригонометрические уравнения