- Молекулярные механизмы регуляции поведения. Серотонин. (Лекция 6)

Содержание
- 2. История Был впервые открыт в кишечнике и назван энтерамином (Espamer, 1940). Позже был открыт в плазме
- 3. Биологически активные индолы Молекула серотонина содержит индольное кольцо и аминогруппу. Производные индола широко распространены в природе
- 4. Метод Фалька-Хилларпа Картирование серотониновых нейронов и их проекций в мозге in situ осуществляется с помощью реакции
- 5. Серотониновая система мозга
- 6. Ядра шва среднего мозга
- 7. Тела серотониновых нейронов в дорзальном ядре шва среднего мозга
- 8. Тонкие и четковидные аксоны в коре мозга крысы
- 9. Спайковая активность серотонинового нейрона Серотониновые нейроны разряжаются с частотой 1-3 спайка/с. Серотониновая система мозга самая экспансивная.
- 10. Синтез серотонина
- 11. Триптофангидроксилаза Триптофангидроксилаза (ТПГ, КФ.1.14.16.4) катализирует гидроксилирование триптофана до серотонина – ключевую стадию синтеза медиатора. Существуют две
- 12. Две формы триптофангидроксилазы Существуют две формы триптофангидроксилазы, ТПГ1 и ТПГ2 (Walther et al., 2003). ТПГ2 кодируется
- 13. C1473G полиморфизм в гене tph2 мыши найден в 2004 г., а G1463A полиморфизм в гене TPH2
- 14. Влияние нокаута гена mTph2 на 5-HT нейроны Нокаут гена mTph2 полностью редуцирует Tph2 or 5-HT в
- 15. Влияние нокаута гена mTph2 на жизнеспособность Tph2-/- мыши характеризуются задержкой роста и дефицитом IGF-1 в крови
- 16. Влияние нокаута гена mTph2 на физиологические функции и поведение Tph2-/- мыши характеризуются увеличением времени сна, снижением
- 17. ДНК мышей 10 линий генотипировали с праймерами, специфическими для C или G аллелей. Обнаружено 7 линий
- 18. Активность ТПГ в среднем мозге мышей с генотипом C/C (Pro) вдвое выше, чем с генотипом G/G
- 19. Сцепление C1473G полиморфизма с активностью ТПГ2 в мозге мышей F2 между линиями C57BL/6 и CC57BR Активность
- 20. Распределение полиморфизма C1473G в популяциях диких мышей (Osipova et al., Genes Brain Behav, 2010).
- 21. Регуляция активности ТПГ2 Изменения активности, вызванные мутациями 11 экзона гена ТПГ2, названы конститутивными. Другая изменчивость активности
- 22. Разрушение серотонина Мыши и люди с генетическим нокаутом гена МАО А характеризуются высокой агрессивностью.
- 23. Полиморфизм 5-HTTLPR промоторе гена транспортера серотонина человека 5-HTT является молекулярным фактором пространственной и временной регуляции функции
- 24. Рецепторы серотонина
- 25. Рецепторы серотонина (механизм)
- 26. 5-HT1A рецептор Кодируется безинтронным геном, локализованным в 13 хромосоме мыши и 5 хромосоме человека. Является пресинаптическим
- 27. Агонисты 5-HT1A рецепторов тормозят спайковую активность серотониновых нейронов и снижают секрецию медиатора
- 28. 5-HT1B рецепторы Кодируются безинтронным геном локализованным на 9 хромосоме мыши и 6 хромосоме человека. Пресинаптические рецепторы.
- 29. 5-HT2A рецептор Кодируется геном, локализованным на 14 хромосоме мыши и 13 хромосоме человека. Содержит два интрона.
- 30. Серотонин и сон M. Jouvet (1967-1969) впервые показал, что снижение уровня серотонина ведет к нарушению сна.
- 31. 5-HT1A вовлечены в регуляцию температуры тела Агонист 5-HT1A рецептора, 8-OH-DPAT, вызывает дозозависимое снижение температуры тела у
- 32. 5-HT в терморегуляции Влияние агониста 5-HT3 рецепторов m-CPBG на температуру тела мышей Влияние агониста 5-HT7 рецепторов
- 33. Участие 5-HT2A рецепторов в регуляции температуры тела у мышей
- 34. Серотонин и гипотермия Глубокая гипотермия, сопровождающаяся снижением температуры тела до 20-25°С, приводит к обратимой активации ТПГ2
- 35. Зимняя спячка у сусликов Уникальная природная адаптация к переживанию неблагоприятных условий – сочетание сна с глубокой
- 36. Пробуждение от зимней спячки
- 37. Блокада синтеза серотонина п-ХФА вызывает пробуждение
- 38. 5-гидрокситриптофан тормозит пробуждение
- 39. Увеличение активности ТПГ-2 у спящих сусликов
- 40. ВПАДЕНИЕ В ЗИМНЮЮ СПЯЧКУ АКТИВИРУЕТ ТПГ-2
- 42. Скачать презентацию
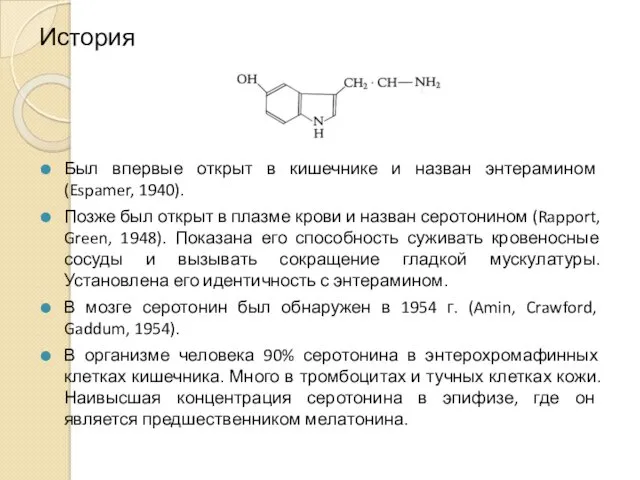
История
Был впервые открыт в кишечнике и назван энтерамином (Espamer, 1940).
Позже был
История
Был впервые открыт в кишечнике и назван энтерамином (Espamer, 1940).
Позже был
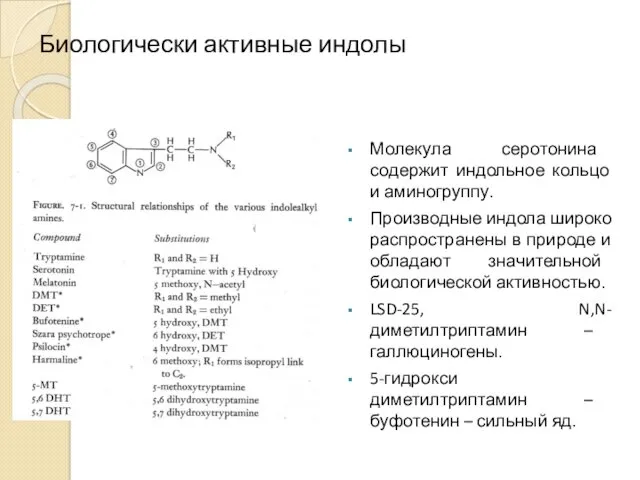
Биологически активные индолы
Молекула серотонина содержит индольное кольцо и аминогруппу.
Производные индола широко
Биологически активные индолы
Молекула серотонина содержит индольное кольцо и аминогруппу.
Производные индола широко
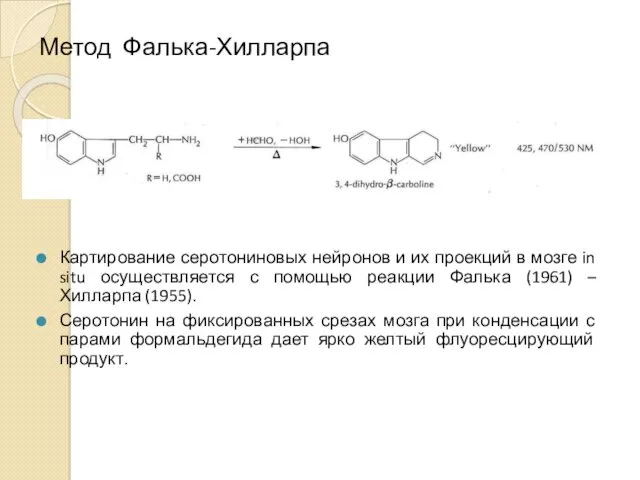
Метод Фалька-Хилларпа
Картирование серотониновых нейронов и их проекций в мозге in situ
Метод Фалька-Хилларпа
Картирование серотониновых нейронов и их проекций в мозге in situ
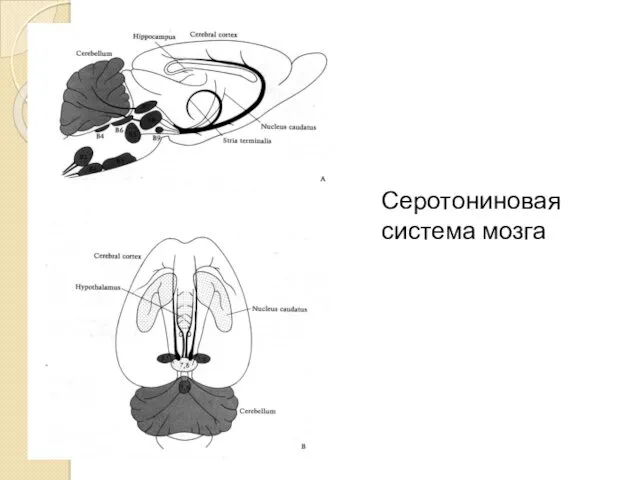
Серотониновая система мозга
Серотониновая система мозга
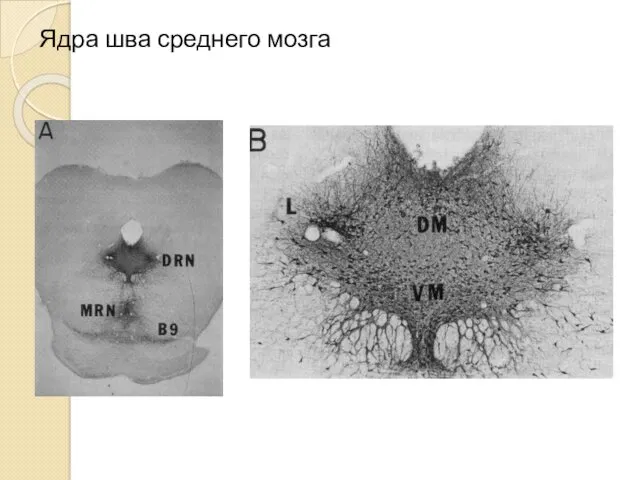
Ядра шва среднего мозга
Ядра шва среднего мозга
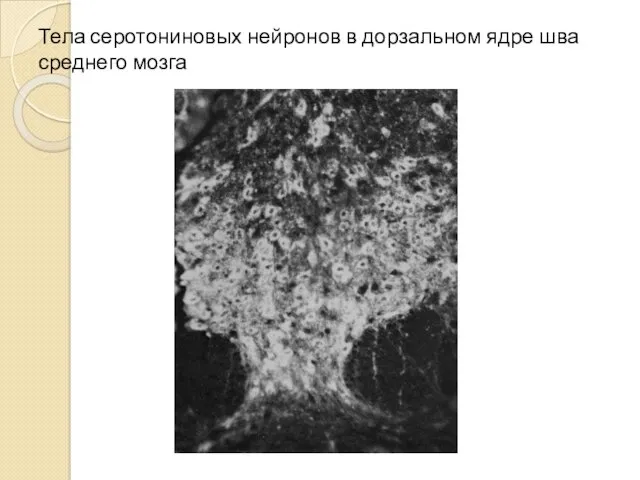
Тела серотониновых нейронов в дорзальном ядре шва среднего мозга
Тела серотониновых нейронов в дорзальном ядре шва среднего мозга
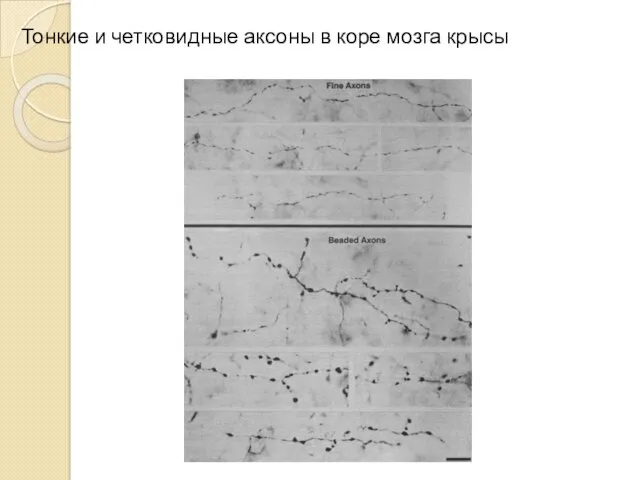
Тонкие и четковидные аксоны в коре мозга крысы
Тонкие и четковидные аксоны в коре мозга крысы
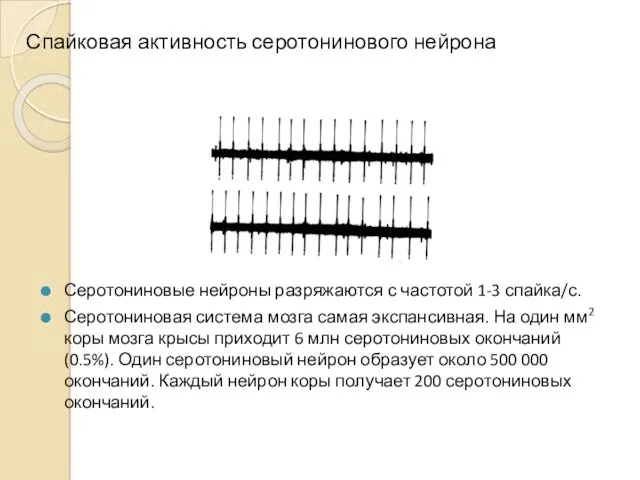
Спайковая активность серотонинового нейрона
Серотониновые нейроны разряжаются с частотой 1-3 спайка/с.
Серотониновая система
Спайковая активность серотонинового нейрона
Серотониновые нейроны разряжаются с частотой 1-3 спайка/с.
Серотониновая система
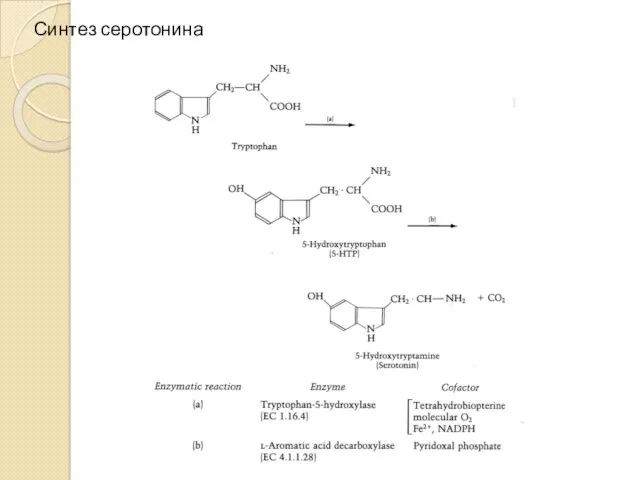
Синтез серотонина
Синтез серотонина

Триптофангидроксилаза
Триптофангидроксилаза (ТПГ, КФ.1.14.16.4) катализирует гидроксилирование триптофана до серотонина – ключевую стадию
Триптофангидроксилаза
Триптофангидроксилаза (ТПГ, КФ.1.14.16.4) катализирует гидроксилирование триптофана до серотонина – ключевую стадию
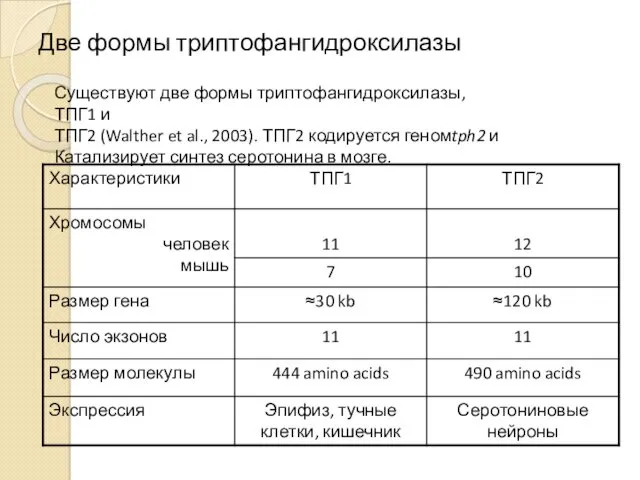
Две формы триптофангидроксилазы
Существуют две формы триптофангидроксилазы, ТПГ1 и
ТПГ2 (Walther et al.,
Две формы триптофангидроксилазы
Существуют две формы триптофангидроксилазы, ТПГ1 и
ТПГ2 (Walther et al.,
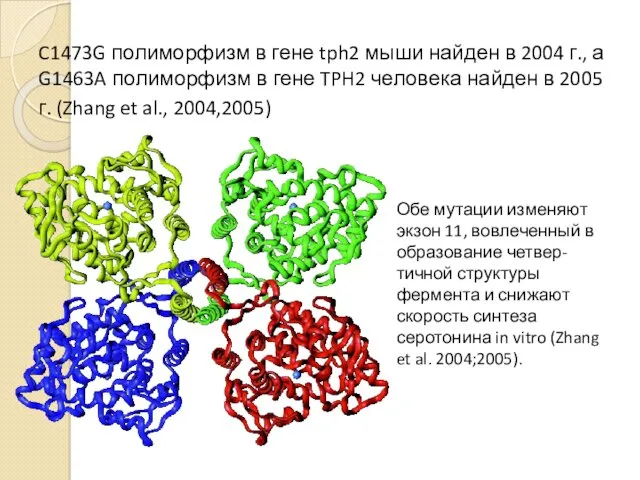
C1473G полиморфизм в гене tph2 мыши найден в 2004 г., а
C1473G полиморфизм в гене tph2 мыши найден в 2004 г., а
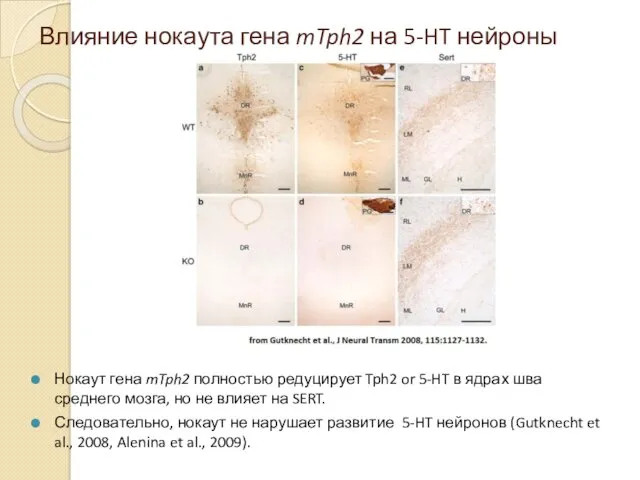
Влияние нокаута гена mTph2 на 5-HT нейроны
Нокаут гена mTph2 полностью редуцирует
Влияние нокаута гена mTph2 на 5-HT нейроны
Нокаут гена mTph2 полностью редуцирует
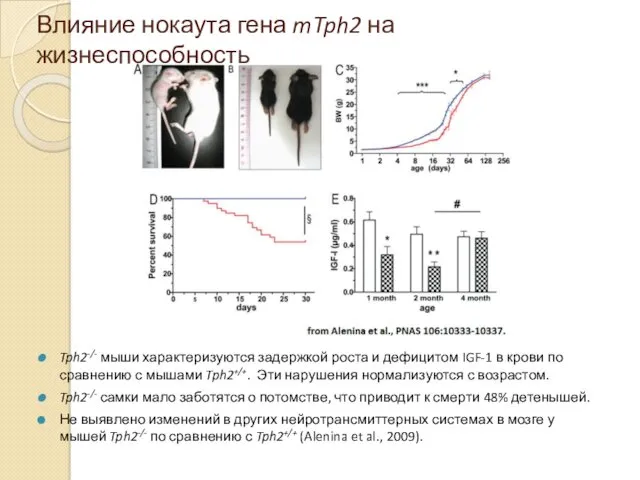
Влияние нокаута гена mTph2 на жизнеспособность
Tph2-/- мыши характеризуются задержкой роста и
Влияние нокаута гена mTph2 на жизнеспособность
Tph2-/- мыши характеризуются задержкой роста и
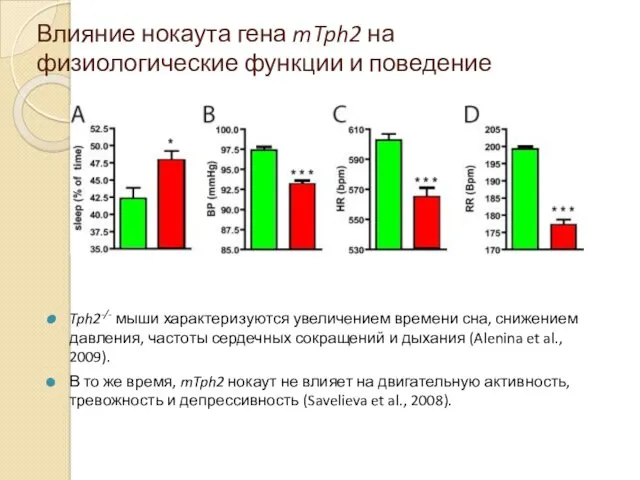
Влияние нокаута гена mTph2 на физиологические функции и поведение
Tph2-/- мыши характеризуются
Влияние нокаута гена mTph2 на физиологические функции и поведение
Tph2-/- мыши характеризуются

ДНК мышей 10 линий генотипировали с праймерами, специфическими для C или
ДНК мышей 10 линий генотипировали с праймерами, специфическими для C или
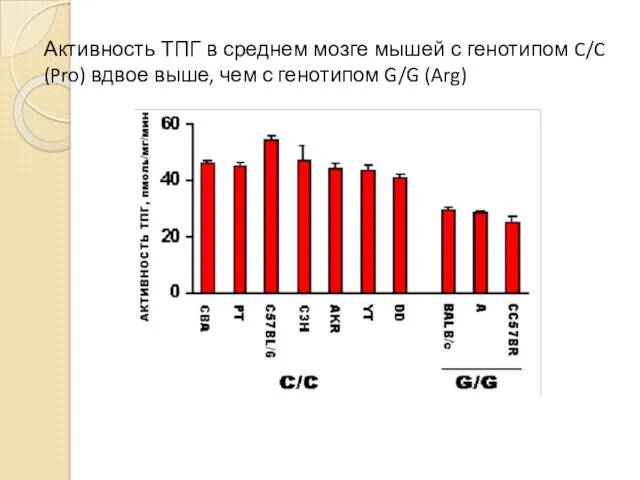
Активность ТПГ в среднем мозге мышей с генотипом C/C (Pro) вдвое
Активность ТПГ в среднем мозге мышей с генотипом C/C (Pro) вдвое
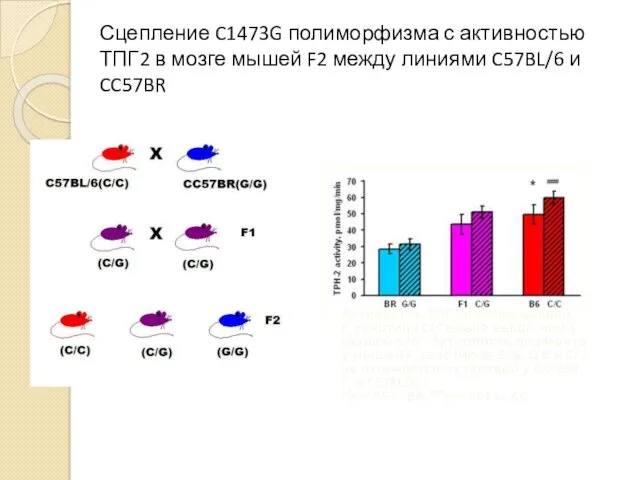
Сцепление C1473G полиморфизма с активностью ТПГ2 в мозге мышей F2 между
Сцепление C1473G полиморфизма с активностью ТПГ2 в мозге мышей F2 между
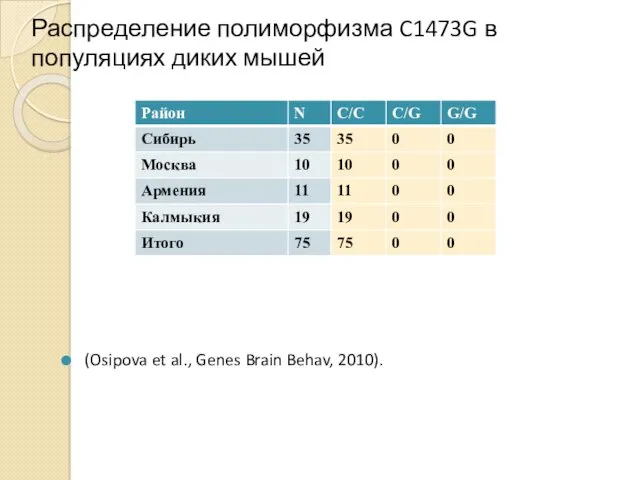
Распределение полиморфизма C1473G в популяциях диких мышей
(Osipova et al., Genes Brain
Распределение полиморфизма C1473G в популяциях диких мышей
(Osipova et al., Genes Brain

Регуляция активности ТПГ2
Изменения активности, вызванные мутациями 11 экзона гена ТПГ2, названы
Регуляция активности ТПГ2
Изменения активности, вызванные мутациями 11 экзона гена ТПГ2, названы
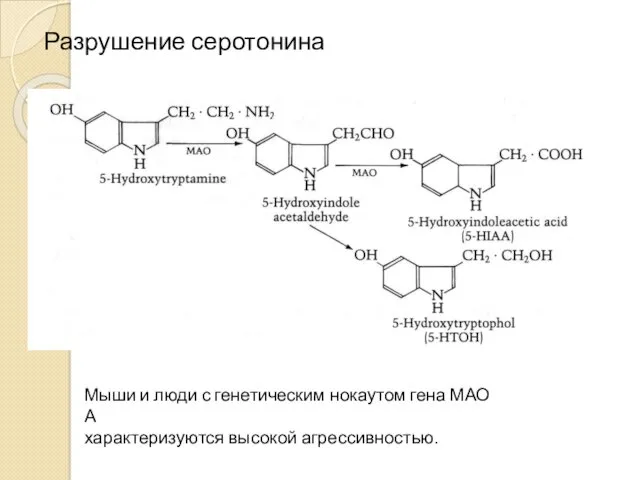
Разрушение серотонина
Мыши и люди с генетическим нокаутом гена МАО А
характеризуются
Разрушение серотонина
Мыши и люди с генетическим нокаутом гена МАО А
характеризуются
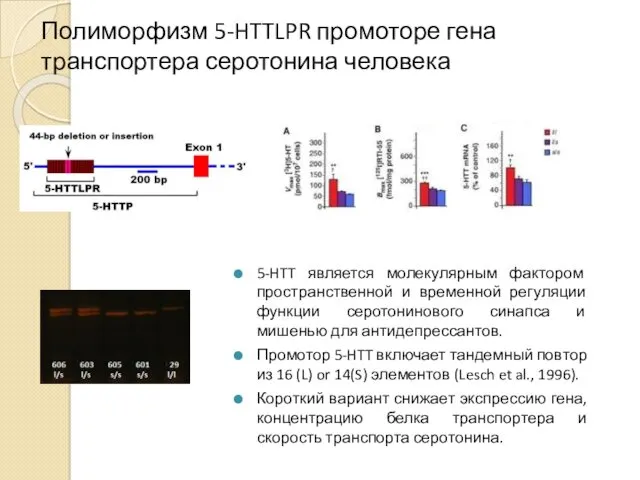
Полиморфизм 5-HTTLPR промоторе гена транспортера серотонина человека
5-HTT является молекулярным фактором пространственной
Полиморфизм 5-HTTLPR промоторе гена транспортера серотонина человека
5-HTT является молекулярным фактором пространственной
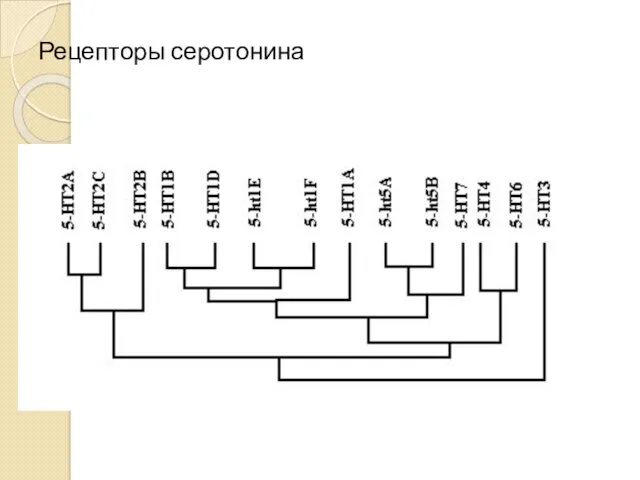
Рецепторы серотонина
Рецепторы серотонина
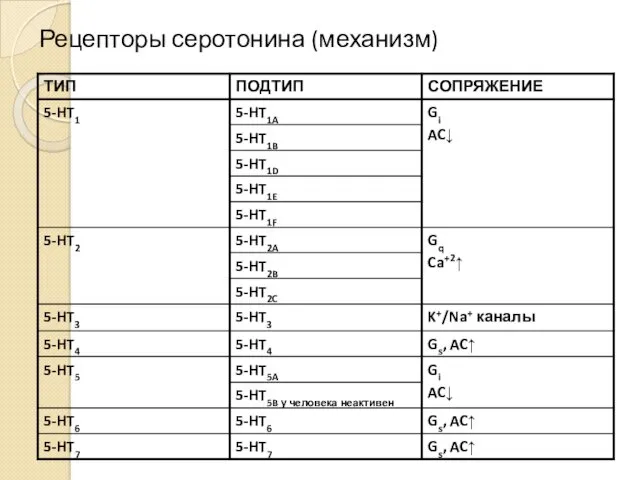
Рецепторы серотонина (механизм)
Рецепторы серотонина (механизм)

5-HT1A рецептор
Кодируется безинтронным геном, локализованным в 13 хромосоме мыши и 5
5-HT1A рецептор
Кодируется безинтронным геном, локализованным в 13 хромосоме мыши и 5
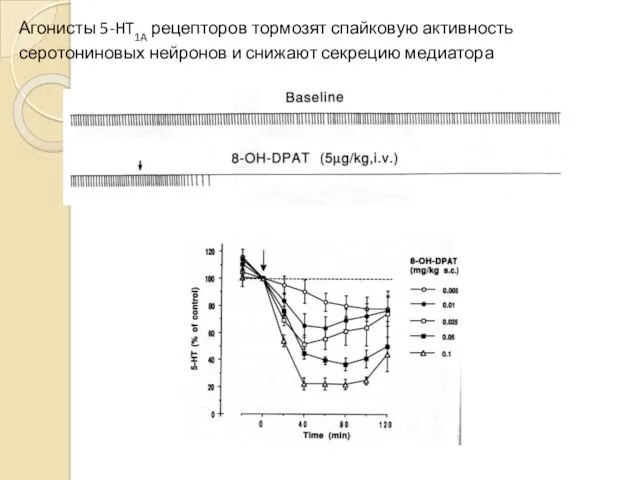
Агонисты 5-HT1A рецепторов тормозят спайковую активность серотониновых нейронов и снижают секрецию
Агонисты 5-HT1A рецепторов тормозят спайковую активность серотониновых нейронов и снижают секрецию

5-HT1B рецепторы
Кодируются безинтронным геном локализованным на 9 хромосоме мыши и 6
5-HT1B рецепторы
Кодируются безинтронным геном локализованным на 9 хромосоме мыши и 6

5-HT2A рецептор
Кодируется геном, локализованным на 14 хромосоме мыши и 13 хромосоме
5-HT2A рецептор
Кодируется геном, локализованным на 14 хромосоме мыши и 13 хромосоме

Серотонин и сон
M. Jouvet (1967-1969) впервые показал, что снижение уровня серотонина
Серотонин и сон
M. Jouvet (1967-1969) впервые показал, что снижение уровня серотонина

5-HT1A вовлечены в регуляцию температуры тела
Агонист 5-HT1A рецептора, 8-OH-DPAT, вызывает дозозависимое
5-HT1A вовлечены в регуляцию температуры тела
Агонист 5-HT1A рецептора, 8-OH-DPAT, вызывает дозозависимое
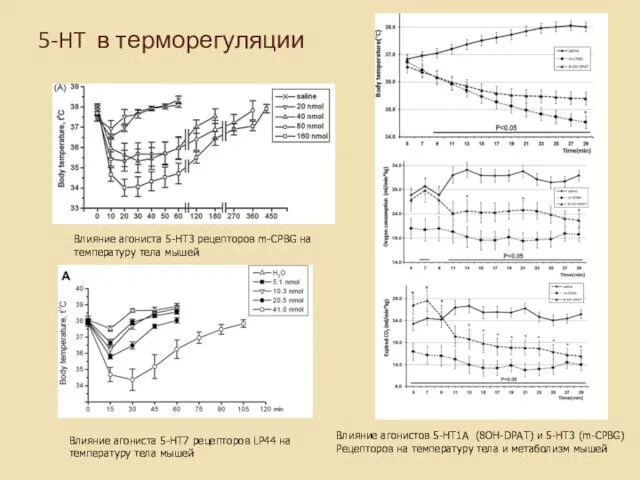
5-HT в терморегуляции
Влияние агониста 5-HT3 рецепторов m-CPBG на
температуру тела мышей
Влияние агониста
5-HT в терморегуляции
Влияние агониста 5-HT3 рецепторов m-CPBG на
температуру тела мышей
Влияние агониста
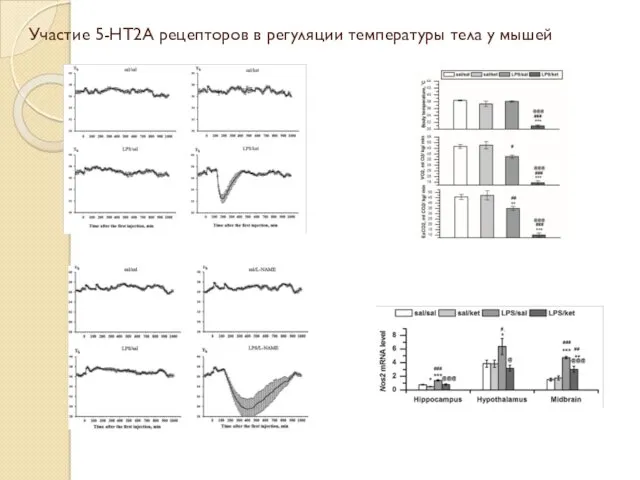
Участие 5-HT2A рецепторов в регуляции температуры тела у мышей
Участие 5-HT2A рецепторов в регуляции температуры тела у мышей

Серотонин и гипотермия
Глубокая гипотермия, сопровождающаяся снижением температуры тела до 20-25°С, приводит
Серотонин и гипотермия
Глубокая гипотермия, сопровождающаяся снижением температуры тела до 20-25°С, приводит

Зимняя спячка у сусликов
Уникальная природная адаптация к переживанию неблагоприятных условий –
Зимняя спячка у сусликов
Уникальная природная адаптация к переживанию неблагоприятных условий –

Пробуждение от зимней спячки
Пробуждение от зимней спячки

Блокада синтеза серотонина п-ХФА вызывает пробуждение
Блокада синтеза серотонина п-ХФА вызывает пробуждение
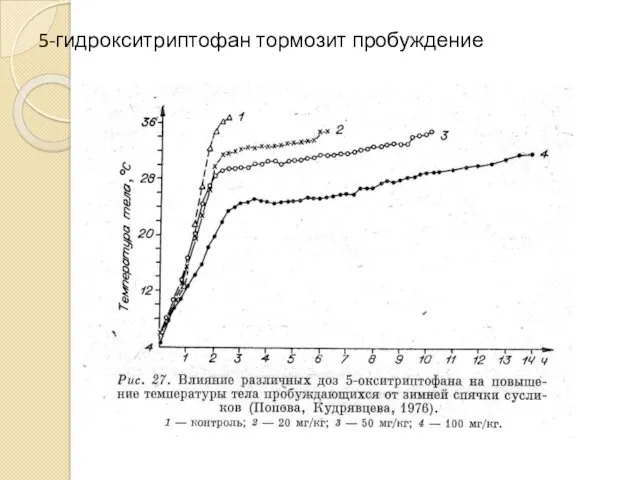
5-гидрокситриптофан тормозит пробуждение
5-гидрокситриптофан тормозит пробуждение
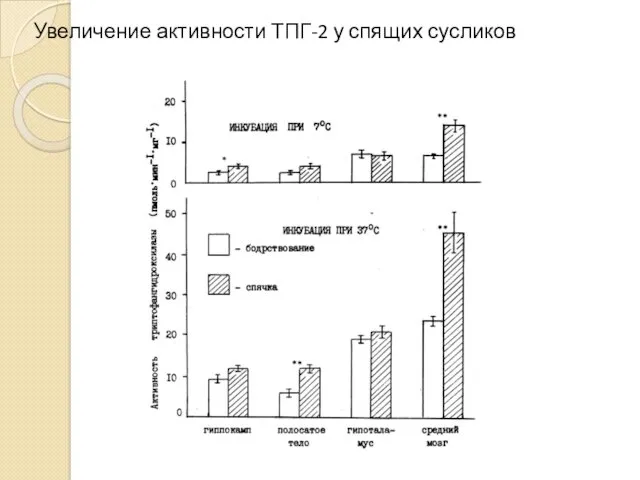
Увеличение активности ТПГ-2 у спящих сусликов
Увеличение активности ТПГ-2 у спящих сусликов

ВПАДЕНИЕ В ЗИМНЮЮ СПЯЧКУ АКТИВИРУЕТ ТПГ-2
ВПАДЕНИЕ В ЗИМНЮЮ СПЯЧКУ АКТИВИРУЕТ ТПГ-2
 Птицы. Что мы о них знаем
Птицы. Что мы о них знаем День энергетика
День энергетика Интерактивная игра Загадки от Бабы Яги
Интерактивная игра Загадки от Бабы Яги Презентация для 8 класса Растворение. Растворимость. Типы растворов.
Презентация для 8 класса Растворение. Растворимость. Типы растворов. Электрические двигатели постоянного тока
Электрические двигатели постоянного тока Понятие культурно-досуговой деятельности
Понятие культурно-досуговой деятельности Портфолио учителя-логопеда Бондаревой Л.Ю.(МБДОУ № 13 г. Архангельска)
Портфолио учителя-логопеда Бондаревой Л.Ю.(МБДОУ № 13 г. Архангельска) Весна
Весна Презентация классного часа Тәмле тел
Презентация классного часа Тәмле тел Жыныс ағзаларының даму ақаулары
Жыныс ағзаларының даму ақаулары Энергоактивные здания с использованием солнечной энергии
Энергоактивные здания с использованием солнечной энергии Консультация для родителей на тему:Почему ребёнок плохо учится?
Консультация для родителей на тему:Почему ребёнок плохо учится? Урал. Географическое положение. История
Урал. Географическое положение. История Презентация к уроку
Презентация к уроку Презентация проекта Новый год у ворот
Презентация проекта Новый год у ворот Организационно-правовые формы предпринимательства
Организационно-правовые формы предпринимательства ИБС. Острый коронарный синдром. Острая сердечная недостаточность. Нестабильная стенокардия
ИБС. Острый коронарный синдром. Острая сердечная недостаточность. Нестабильная стенокардия Construction materials
Construction materials Тема 7. Транспортные узлы. Лекция 2. Транспортный сервис в транспортных узлах
Тема 7. Транспортные узлы. Лекция 2. Транспортный сервис в транспортных узлах ПДД для детей
ПДД для детей Это волшебное слово- мама! Презентация
Это волшебное слово- мама! Презентация История православного Рождества
История православного Рождества Интегрированный урок (математика-география) Применение теоремы Пифагора в сельском хозяйстве
Интегрированный урок (математика-география) Применение теоремы Пифагора в сельском хозяйстве Зима будет Снежной с линией Winter Care
Зима будет Снежной с линией Winter Care Hauberk - прайс-лист
Hauberk - прайс-лист Исследовательская практика младших школьников
Исследовательская практика младших школьников экологический калейдоскоп
экологический калейдоскоп Организация сюжетно-ролевой игр в старшей группеБольница
Организация сюжетно-ролевой игр в старшей группеБольница