- Мышечные ткани

Содержание
- 2. ПО ИСТОЧНИКАМ ПРОИСХОЖДЕНИЯ РАЗЛИЧАЮТ 1.Скелетная мышечная ткань(соматического типа),возникающая из миотомов сомитов. 2.Сердечная мышечная ткань(целомического типа),возникающая из
- 3. ПРОДОЛЖЕНИЕ 5.Мионейральная ткань,возникающая из нейроэктодермы. 6.Миофибробласты,проявляющие свойства фибробластов и гладких миоцитов,их вариантами являются миоидные клетки извитого
- 4. МОРФОЛОГИЧЕСКАЯ КЛАССИФИКАЦИЯ Она основана на наличии или отсутствии исчерченности миофибрилл и различают: 1.исчерченные мышечные ткани, к
- 6. Особенности гладких миоцитов Форма веретеновидная, звездчатая Размер от 20 мкм до 500 мкм Окружены базальной мембраной
- 7. КАВЕОЛЫ Кавеолы- колбовидные впячивания поверхности сарколемы , которые открыты в сторону межклеточного пространства и составляют 1/3
- 8. Гладкая мышечная ткань
- 9. Ультраструктура гладкого миоцита
- 10. Сократительный и опорный аппараты гладкого миоцита Фаза расслабления Фаза сокращения
- 11. АКТИНОВЫЕ МИКРОФИЛАМЕТЫ 1.Количественно преобладают над миозино- выми и состоят из белка - актина. 2.Располагаются в цитоплазме
- 12. Миозиновые микрофиламенты 1.Выявляются в виде нитей только в процессе сокращения. 2.В состоянии покоя миозин находится в
- 13. ПРОДОЛЖЕНИЕ 4.Сборка из молекул миозина толстых миозиновых филаментов,а также взаимо- действие актиновых и миозиновых фила- ментов
- 14. ГЛАДКИЕ МИОЦИТЫ 1.Функционируют в виде комплексов. 2.Комплекс состоит из 10 – 12 миоцитов,которые связаны друг с
- 15. Компоненты миоцитарного комплекса 1.Сократительныые миоциты,выполняющие функцию сокращения. 2.Секреторные миоциты,синтезирующие и секретирующие межклеточное вещество. 3.Миоциты пейсмейкеры, генерирующие
- 16. РЕГЕНЕРАЦИЯ ГЛАДКОЙ МЫШЕЧНОЙ ТКАНИ ОСУЩЕСТВЛЯЕТСЯ ЗА СЧЕТ 1.малодифференцированных клеток. 2.адвентициальных клеток 3.Перицитов 4.При повреждении за счет
- 17. МЕХАНИЗМ СОКРАЩЕНИЯ ГЛАДКИХ МИОЦИТОВ ЗАКЛЮЧАЕТСЯ ВО ВЗАИМОДЕЙСТВИИ АКТИНОВЫХ И МИОЗИНОВЫХ ФИЛАМЕНТОВ, КОТОРОЕ ИНИЦИИРУЕТСЯ ИОНАМИ КАЛЬЦИЯ ВЫДЕЛЯЮТ
- 18. Са – ЗАВИСИМАЯ ЧАСТЬ СОКРАЩЕНИЯ ПРОТЕКАЕТ СЛЕДУЮЩИМ ОБРАЗОМ 1.Действие сократительного стимула(нервный импульс). 2.Происходит открытие кальциевых каналов
- 19. ПРОДОЛЖЕНИЕ 5.Киназа легких цепей миозина фосфорилирует легкие цепи головок миозина и в таком состоянии они могут
- 20. ПРОДОЛЖЕНИЕ 6.Прекращает сокращение фермент фосфатаза миозина,вызывающая дефосфорилирования легких цепей миозина.Особенность гладких мышц заключается в том,что после
- 21. ГИСТОГЕНЕЗ СКЕЛЕТНОЙ МЫШЕЧНОЙ ТКАНИ Источником развития являются миотомы сомитов.Клетки миотомов мигрируют в места закладки мышц и
- 22. продолжение миосимпласт, окруженный базальной мембраной.Начинается синтез миофибрилл.Часть миобластов превращаются в миосателиоциты. 3.Стадия миотубул. Миосимпласты увеличиваются в
- 23. СТРОЕНИЕ ПОПЕРЕЧНО-ПОЛОСАТОГО МЫШЕЧНОГО ВОЛОКНА Структурно-функциональными элементами скелетной мышечной ткани являются мышечные волокна,состоящие из 2-х частей: 1.симпластической
- 24. ПРОДОЛЖЕНИЕ Толщина волокна составляет 50-100 мкм, длина 20 – 30 см.Снаружи симпласт покрыт сарколеммой,состоящей из плазмолеммы
- 25. МИОФИБРИЛЛЫ Это специальные органеллы,занимающие основную часть волокна.В одном волокне их до 2000 . Диаметр миофибрилл составляет
- 26. ПРОДОЛЖЕНИЕ В саркоплазме мышечного волокна находятся большое количество органелл общего значения (За исключением центриолей),включения гликогена, липидов,а
- 27. ПРОДОЛЖЕНИЕ Посередине изотропного диска проходит Z –линия ( на уровне Т- трубочки ),где закрепляются актиновые филаменты
- 28. САРКОМЕР Саркомер - это структурно-функциональная единица миофибриллы,длиной 2,5 - 3 мкм. Саркомер охватывает полный диск А
- 29. ПРОДОЛЖЕНИЕ 2.Миозиновые филаменты располагаются только в анизотропном диске. Светлая Н -полоска состоит только из миозина и
- 30. Скелетная мышечная ткань
- 31. Схема строения поперечнополосатого мышечного волокна
- 32. Актиновые микрофиламенты Диаметр = 5 нм, длина – 1 мкм В их состав входят 3 вида
- 33. АКТИНОВЫЕ МИКРОФИЛАИЕНТЫ Построены следующим образом. Цепочки глобулярных молекул актина образуют двойную спираль, а в желобках этой
- 34. Строение актиновых микрофиламентов
- 35. Тропониновый комплекс состоит из: Субъединицы (ТпТ) , осуществляющая прикрепление тропонинового комплекса к тропомиозину Субъединицы (ТпС), отвечающей
- 36. Строение саркомера
- 37. Миозиновые микрофиламенты включают: Миозин – составляет основную массу Белок титин – наподобие пружины связывает концы толстых
- 38. Строение миозиновых микрофиламентов
- 39. В молекуле миозина различают 1.Каждая миозиновая нить состоит из 300 -400 молекул миозина. Половина молекул обращена
- 40. ПРОДОЛЖЕНИЕ &легкий меромиозин,который обеспечивает агрегацию молекул миозина ТЯЖЕЛЫЙ МЕРОМИОЗИН имеет два фрагмента: S-1 и S-2. 1).
- 41. ПРОДОЛЖЕНИЕ МОЛЕКУЛА МИОЗИНА ИМЕЕТ ДВА шарнирных участка ,которые позволяют молекуле изменять конформацию: *первый находится в области
- 42. Строение молекулы миозина
- 43. Типы мышечных волокон Красные мышечные волокна. Имеют небольшой диаметр Содержат много миоглобина Содержат много митохондрий В
- 44. Типы мышечных волокон 2. Белые мышечные волокна Имеют большой диаметр Содержат мало миоглобина Меньшее содержание митохондрий,
- 45. МЕХАНИЗМ СОКРАЩЕНИЯ ПОПЕРЕЧНО- ПОЛОСАТОГО МЫШЕЧНОГО ВОЛОКНА Важное функциональное значение имеют 1.Т-трубочки,которые образованы впячиванием сарколеммы внутрь симпласта.Располагаются
- 46. БОЛЬШИЕ ТЕРМИНАЛЬНЫЕ ЦИСТЕРНЫ ГЛАДКОЙ ЭПС Здесь депонируются ионы Са,мембрана имеет рецепторы к инозитол -3-фосфат, инозитол -
- 47. КАЛЬЦИЙ-ЗАВИСИМАЯ ЧАСТЬ СОКРАЩЕНИЯ СМЫСЛ ПРОЦЕССА На актине должны открыться участки для связывания миозина,в противном случае миозин
- 48. ПРОДОЛЖЕНИЕ 1.Поступление сократительного импульса. 2.Прохождение импульса по цитомембране. 3.Прохождение импульса по мембране Т-трубочек 4.Выработка инозитол –
- 49. ПРОДОЛЖЕНИЕ 7.Открытие кальциевых каналов в мембранах ретикулума. 8.Выход ионов кальция из ретикулума в цитозоль. 9.Диффузия ионов
- 50. АТФ-ЗАВИСИМАЯ ЧАСТЬ СОКРАЩЕНИЯ 1.Головка миозина присоединяет молекулу АТФ. 2.Головка миозина расщепляет молекулу АТФ до АДФ и
- 51. ПРОДОЛЖЕНИЕ молекулы актина(молекула миозина тянет на себя актин). 4.Головка миозина присоединяет новую молекулу АТФ и только
- 52. Сердечная мышечная ткань Структурно-функциональной единицей являются поперечнополосатые кардиомиоциты Границами клеток являются вставочные пластинки, в пределах которых
- 53. Сердечная мышечная ткань Цепочка кардиомиоцитов образует мышечные волокна Волокна соединяются анастомозами (приобретают сетчатую структуру) Кардиомиоциты по
- 54. Сердечная поперечнополосатая мышечная ткань
- 56. Скачать презентацию

ПО ИСТОЧНИКАМ ПРОИСХОЖДЕНИЯ
РАЗЛИЧАЮТ
1.Скелетная мышечная ткань(соматического
типа),возникающая из миотомов сомитов.
2.Сердечная мышечная ткань(целомического
ПО ИСТОЧНИКАМ ПРОИСХОЖДЕНИЯ
РАЗЛИЧАЮТ
1.Скелетная мышечная ткань(соматического
типа),возникающая из миотомов сомитов.
2.Сердечная мышечная ткань(целомического

ПРОДОЛЖЕНИЕ
5.Мионейральная ткань,возникающая из
нейроэктодермы.
6.Миофибробласты,проявляющие свойства фибробластов и гладких миоцитов,их
вариантами
ПРОДОЛЖЕНИЕ
5.Мионейральная ткань,возникающая из
нейроэктодермы.
6.Миофибробласты,проявляющие свойства фибробластов и гладких миоцитов,их
вариантами

МОРФОЛОГИЧЕСКАЯ КЛАССИФИКАЦИЯ
Она основана на наличии или отсутствии
исчерченности миофибрилл
МОРФОЛОГИЧЕСКАЯ КЛАССИФИКАЦИЯ
Она основана на наличии или отсутствии
исчерченности миофибрилл
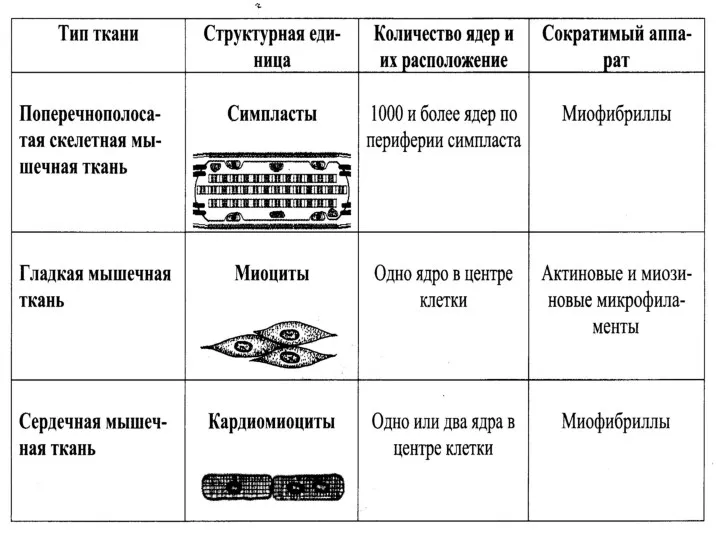

Особенности гладких миоцитов
Форма веретеновидная, звездчатая
Размер от 20 мкм до 500 мкм
Окружены
Особенности гладких миоцитов
Форма веретеновидная, звездчатая
Размер от 20 мкм до 500 мкм
Окружены

КАВЕОЛЫ
Кавеолы- колбовидные впячивания поверхности
сарколемы , которые открыты в сторону
межклеточного пространства
КАВЕОЛЫ
Кавеолы- колбовидные впячивания поверхности
сарколемы , которые открыты в сторону
межклеточного пространства

Гладкая мышечная ткань
Гладкая мышечная ткань
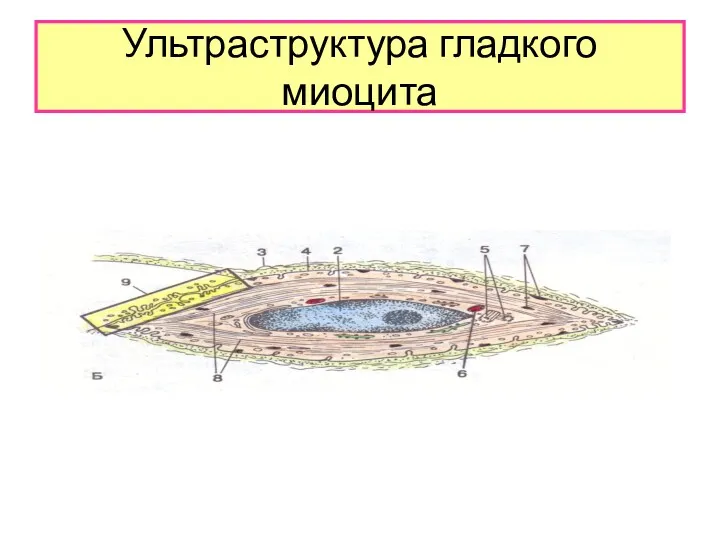
Ультраструктура гладкого миоцита
Ультраструктура гладкого миоцита
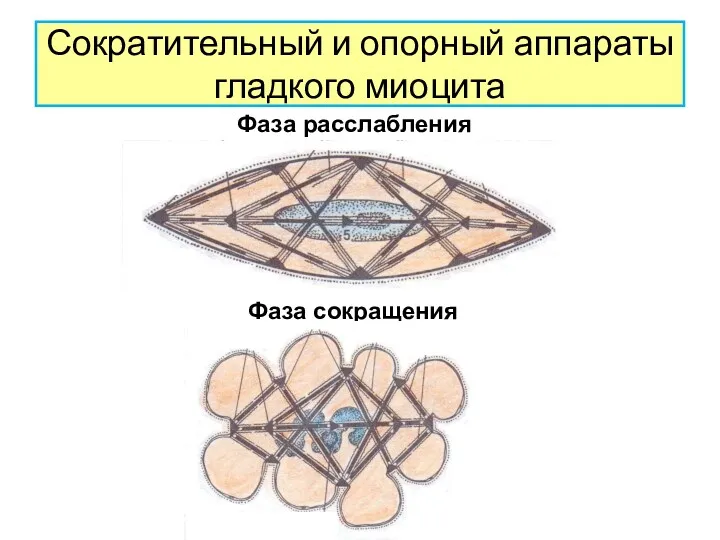
Сократительный и опорный аппараты гладкого миоцита
Фаза расслабления
Фаза сокращения
Сократительный и опорный аппараты гладкого миоцита
Фаза расслабления
Фаза сокращения

АКТИНОВЫЕ МИКРОФИЛАМЕТЫ
1.Количественно преобладают над миозино-
выми и состоят из белка
АКТИНОВЫЕ МИКРОФИЛАМЕТЫ
1.Количественно преобладают над миозино-
выми и состоят из белка

Миозиновые микрофиламенты
1.Выявляются в виде нитей только в процессе
сокращения.
2.В
Миозиновые микрофиламенты
1.Выявляются в виде нитей только в процессе
сокращения.
2.В

ПРОДОЛЖЕНИЕ
4.Сборка из молекул миозина толстых
миозиновых филаментов,а также взаимо-
действие
ПРОДОЛЖЕНИЕ
4.Сборка из молекул миозина толстых
миозиновых филаментов,а также взаимо-
действие

ГЛАДКИЕ МИОЦИТЫ
1.Функционируют в виде комплексов.
2.Комплекс состоит из 10 – 12 миоцитов,которые
ГЛАДКИЕ МИОЦИТЫ
1.Функционируют в виде комплексов.
2.Комплекс состоит из 10 – 12 миоцитов,которые

Компоненты миоцитарного комплекса
1.Сократительныые миоциты,выполняющие
функцию сокращения.
2.Секреторные миоциты,синтезирующие и
секретирующие
Компоненты миоцитарного комплекса
1.Сократительныые миоциты,выполняющие
функцию сокращения.
2.Секреторные миоциты,синтезирующие и
секретирующие

РЕГЕНЕРАЦИЯ ГЛАДКОЙ МЫШЕЧНОЙ ТКАНИ ОСУЩЕСТВЛЯЕТСЯ ЗА СЧЕТ
1.малодифференцированных клеток.
2.адвентициальных клеток
3.Перицитов
4.При
РЕГЕНЕРАЦИЯ ГЛАДКОЙ МЫШЕЧНОЙ ТКАНИ ОСУЩЕСТВЛЯЕТСЯ ЗА СЧЕТ
1.малодифференцированных клеток.
2.адвентициальных клеток
3.Перицитов
4.При

МЕХАНИЗМ СОКРАЩЕНИЯ ГЛАДКИХ
МИОЦИТОВ
ЗАКЛЮЧАЕТСЯ ВО ВЗАИМОДЕЙСТВИИ АКТИНОВЫХ И МИОЗИНОВЫХ ФИЛАМЕНТОВ,
КОТОРОЕ ИНИЦИИРУЕТСЯ
МЕХАНИЗМ СОКРАЩЕНИЯ ГЛАДКИХ
МИОЦИТОВ
ЗАКЛЮЧАЕТСЯ ВО ВЗАИМОДЕЙСТВИИ АКТИНОВЫХ И МИОЗИНОВЫХ ФИЛАМЕНТОВ, КОТОРОЕ ИНИЦИИРУЕТСЯ

Са – ЗАВИСИМАЯ ЧАСТЬ СОКРАЩЕНИЯ
ПРОТЕКАЕТ СЛЕДУЮЩИМ ОБРАЗОМ
1.Действие сократительного стимула(нервный
импульс).
2.Происходит открытие
Са – ЗАВИСИМАЯ ЧАСТЬ СОКРАЩЕНИЯ
ПРОТЕКАЕТ СЛЕДУЮЩИМ ОБРАЗОМ
1.Действие сократительного стимула(нервный
импульс).
2.Происходит открытие

ПРОДОЛЖЕНИЕ
5.Киназа легких цепей миозина фосфорилирует
легкие цепи головок миозина и в
ПРОДОЛЖЕНИЕ
5.Киназа легких цепей миозина фосфорилирует
легкие цепи головок миозина и в

ПРОДОЛЖЕНИЕ
6.Прекращает сокращение фермент фосфатаза
миозина,вызывающая дефосфорилирования
легких цепей миозина.Особенность гладких
мышц
ПРОДОЛЖЕНИЕ
6.Прекращает сокращение фермент фосфатаза
миозина,вызывающая дефосфорилирования
легких цепей миозина.Особенность гладких
мышц

ГИСТОГЕНЕЗ СКЕЛЕТНОЙ МЫШЕЧНОЙ ТКАНИ
Источником развития являются миотомы
сомитов.Клетки миотомов мигрируют в места
закладки
ГИСТОГЕНЕЗ СКЕЛЕТНОЙ МЫШЕЧНОЙ ТКАНИ
Источником развития являются миотомы
сомитов.Клетки миотомов мигрируют в места
закладки

продолжение
миосимпласт, окруженный базальной
мембраной.Начинается синтез миофибрилл.Часть
миобластов превращаются в миосателиоциты.
3.Стадия миотубул. Миосимпласты
продолжение
миосимпласт, окруженный базальной
мембраной.Начинается синтез миофибрилл.Часть
миобластов превращаются в миосателиоциты.
3.Стадия миотубул. Миосимпласты

СТРОЕНИЕ ПОПЕРЕЧНО-ПОЛОСАТОГО МЫШЕЧНОГО ВОЛОКНА
Структурно-функциональными элементами
скелетной мышечной ткани являются мышечные
волокна,состоящие из 2-х
СТРОЕНИЕ ПОПЕРЕЧНО-ПОЛОСАТОГО МЫШЕЧНОГО ВОЛОКНА
Структурно-функциональными элементами
скелетной мышечной ткани являются мышечные
волокна,состоящие из 2-х

ПРОДОЛЖЕНИЕ
Толщина волокна составляет 50-100 мкм,
длина 20 – 30 см.Снаружи симпласт покрыт
сарколеммой,состоящей
ПРОДОЛЖЕНИЕ
Толщина волокна составляет 50-100 мкм,
длина 20 – 30 см.Снаружи симпласт покрыт
сарколеммой,состоящей

МИОФИБРИЛЛЫ
Это специальные органеллы,занимающие
основную часть волокна.В одном волокне их до
2000 .
МИОФИБРИЛЛЫ
Это специальные органеллы,занимающие
основную часть волокна.В одном волокне их до
2000 .

ПРОДОЛЖЕНИЕ
В саркоплазме мышечного волокна находятся
большое количество органелл общего значения
(За исключением
ПРОДОЛЖЕНИЕ
В саркоплазме мышечного волокна находятся
большое количество органелл общего значения
(За исключением

ПРОДОЛЖЕНИЕ
Посередине изотропного диска проходит
Z –линия ( на уровне Т- трубочки ),где
закрепляются
ПРОДОЛЖЕНИЕ
Посередине изотропного диска проходит
Z –линия ( на уровне Т- трубочки ),где
закрепляются

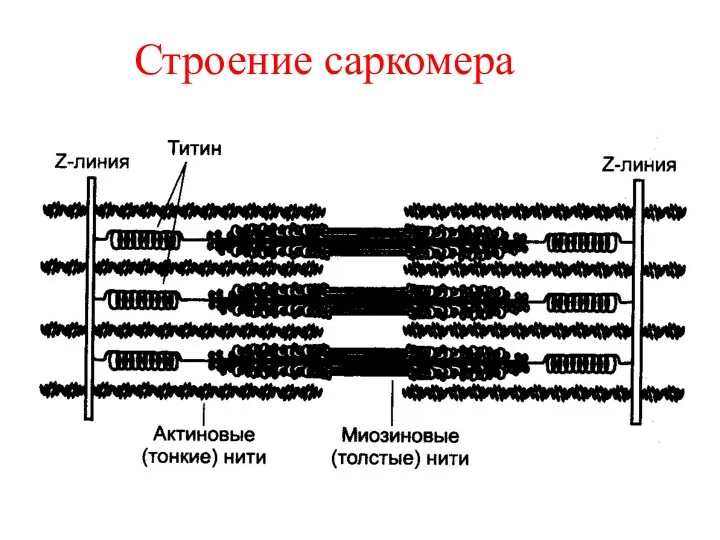
САРКОМЕР
Саркомер - это структурно-функциональная
единица миофибриллы,длиной 2,5 - 3 мкм.
Саркомер охватывает
САРКОМЕР
Саркомер - это структурно-функциональная
единица миофибриллы,длиной 2,5 - 3 мкм.
Саркомер охватывает

ПРОДОЛЖЕНИЕ
2.Миозиновые филаменты располагаются
только в анизотропном диске.
Светлая Н -полоска состоит только
ПРОДОЛЖЕНИЕ
2.Миозиновые филаменты располагаются
только в анизотропном диске.
Светлая Н -полоска состоит только

Скелетная мышечная ткань
Скелетная мышечная ткань

Схема строения поперечнополосатого мышечного волокна
Схема строения поперечнополосатого мышечного волокна

Актиновые микрофиламенты
Диаметр = 5 нм, длина – 1 мкм
В их состав
Актиновые микрофиламенты
Диаметр = 5 нм, длина – 1 мкм
В их состав

АКТИНОВЫЕ МИКРОФИЛАИЕНТЫ
Построены следующим образом.
Цепочки глобулярных молекул актина образуют
двойную спираль, а в
АКТИНОВЫЕ МИКРОФИЛАИЕНТЫ
Построены следующим образом.
Цепочки глобулярных молекул актина образуют
двойную спираль, а в
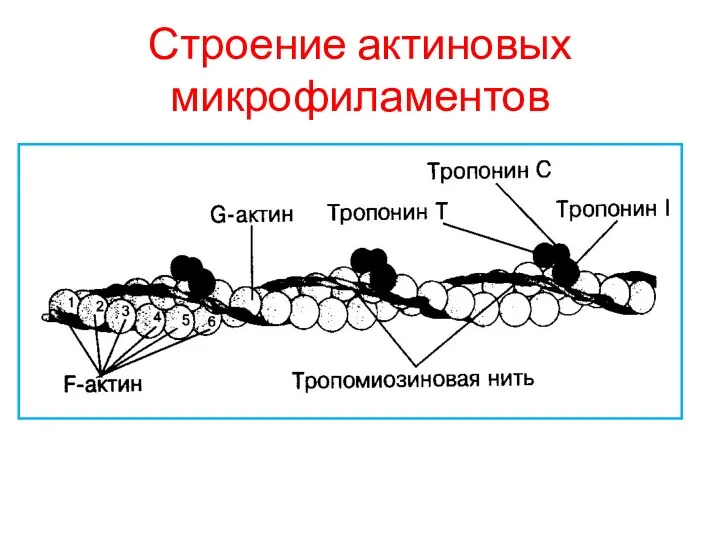
Строение актиновых микрофиламентов
Строение актиновых микрофиламентов

Тропониновый комплекс состоит из:
Субъединицы (ТпТ) , осуществляющая прикрепление тропонинового комплекса к
Тропониновый комплекс состоит из:
Субъединицы (ТпТ) , осуществляющая прикрепление тропонинового комплекса к
Строение саркомера
Строение саркомера

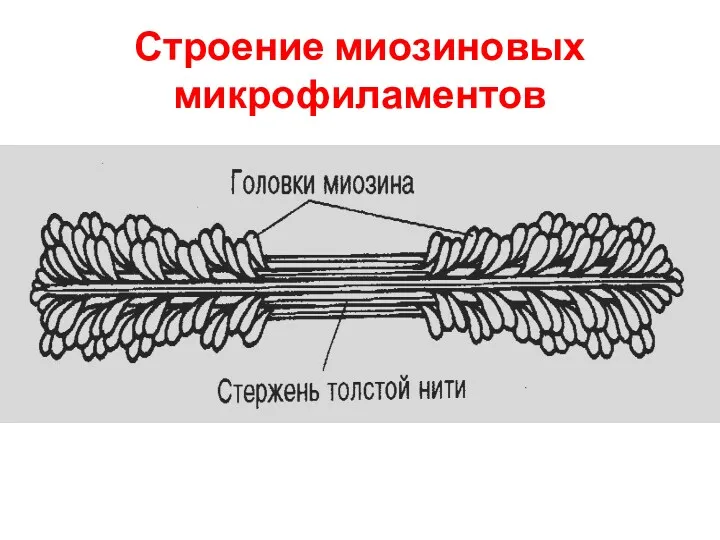
Миозиновые микрофиламенты включают:
Миозин – составляет основную массу
Белок титин – наподобие пружины
Миозиновые микрофиламенты включают:
Миозин – составляет основную массу
Белок титин – наподобие пружины
Строение миозиновых микрофиламентов
Строение миозиновых микрофиламентов

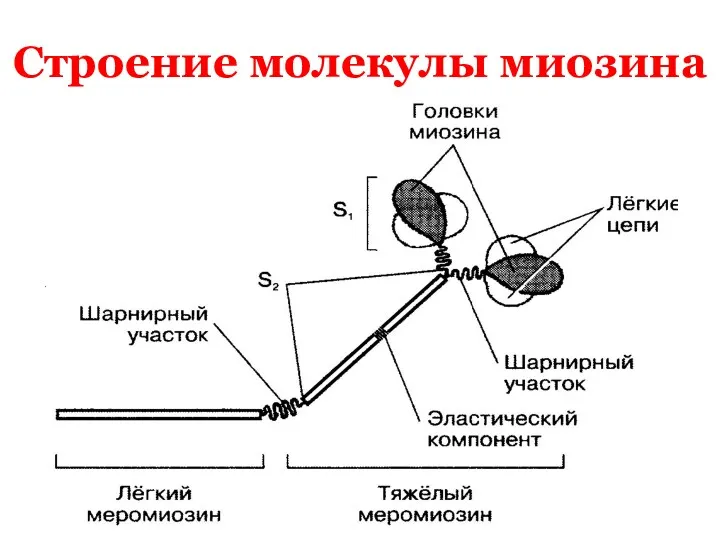
В молекуле миозина различают
1.Каждая миозиновая нить состоит из 300 -400
молекул
В молекуле миозина различают
1.Каждая миозиновая нить состоит из 300 -400
молекул

ПРОДОЛЖЕНИЕ
&легкий меромиозин,который обеспечивает
агрегацию молекул миозина
ТЯЖЕЛЫЙ МЕРОМИОЗИН
имеет два
ПРОДОЛЖЕНИЕ
&легкий меромиозин,который обеспечивает
агрегацию молекул миозина
ТЯЖЕЛЫЙ МЕРОМИОЗИН
имеет два

ПРОДОЛЖЕНИЕ
МОЛЕКУЛА МИОЗИНА ИМЕЕТ ДВА
шарнирных участка ,которые позволяют
молекуле
ПРОДОЛЖЕНИЕ
МОЛЕКУЛА МИОЗИНА ИМЕЕТ ДВА
шарнирных участка ,которые позволяют
молекуле
Строение молекулы миозина
Строение молекулы миозина

Типы мышечных волокон
Красные мышечные волокна.
Имеют небольшой диаметр
Содержат много миоглобина
Содержат много
Типы мышечных волокон
Красные мышечные волокна.
Имеют небольшой диаметр
Содержат много миоглобина
Содержат много

Типы мышечных волокон
2. Белые мышечные волокна
Имеют большой диаметр
Содержат мало миоглобина
Меньшее содержание
Типы мышечных волокон
2. Белые мышечные волокна
Имеют большой диаметр
Содержат мало миоглобина
Меньшее содержание

МЕХАНИЗМ СОКРАЩЕНИЯ ПОПЕРЕЧНО-
ПОЛОСАТОГО МЫШЕЧНОГО ВОЛОКНА
Важное функциональное значение имеют
1.Т-трубочки,которые образованы
МЕХАНИЗМ СОКРАЩЕНИЯ ПОПЕРЕЧНО-
ПОЛОСАТОГО МЫШЕЧНОГО ВОЛОКНА
Важное функциональное значение имеют
1.Т-трубочки,которые образованы

БОЛЬШИЕ ТЕРМИНАЛЬНЫЕ ЦИСТЕРНЫ ГЛАДКОЙ ЭПС
Здесь депонируются ионы Са,мембрана
имеет рецепторы
БОЛЬШИЕ ТЕРМИНАЛЬНЫЕ ЦИСТЕРНЫ ГЛАДКОЙ ЭПС
Здесь депонируются ионы Са,мембрана
имеет рецепторы

КАЛЬЦИЙ-ЗАВИСИМАЯ ЧАСТЬ СОКРАЩЕНИЯ
СМЫСЛ ПРОЦЕССА
На актине должны открыться участки для
КАЛЬЦИЙ-ЗАВИСИМАЯ ЧАСТЬ СОКРАЩЕНИЯ
СМЫСЛ ПРОЦЕССА
На актине должны открыться участки для

ПРОДОЛЖЕНИЕ
1.Поступление сократительного импульса.
2.Прохождение импульса по цитомембране.
3.Прохождение импульса по
ПРОДОЛЖЕНИЕ
1.Поступление сократительного импульса.
2.Прохождение импульса по цитомембране.
3.Прохождение импульса по

ПРОДОЛЖЕНИЕ
7.Открытие кальциевых каналов в мембранах
ретикулума.
8.Выход ионов кальция из ретикулума в
ПРОДОЛЖЕНИЕ
7.Открытие кальциевых каналов в мембранах
ретикулума.
8.Выход ионов кальция из ретикулума в

АТФ-ЗАВИСИМАЯ ЧАСТЬ СОКРАЩЕНИЯ
1.Головка миозина присоединяет молекулу АТФ.
2.Головка миозина расщепляет молекулу АТФ
АТФ-ЗАВИСИМАЯ ЧАСТЬ СОКРАЩЕНИЯ
1.Головка миозина присоединяет молекулу АТФ.
2.Головка миозина расщепляет молекулу АТФ

ПРОДОЛЖЕНИЕ
молекулы актина(молекула миозина тянет
на себя актин).
4.Головка миозина присоединяет новую
ПРОДОЛЖЕНИЕ
молекулы актина(молекула миозина тянет
на себя актин).
4.Головка миозина присоединяет новую

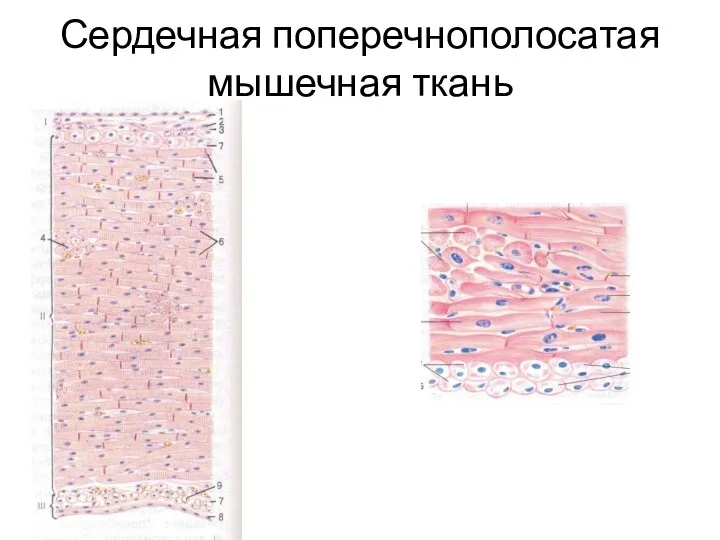
Сердечная мышечная ткань
Структурно-функциональной единицей являются поперечнополосатые кардиомиоциты
Границами клеток являются вставочные пластинки,
Сердечная мышечная ткань
Структурно-функциональной единицей являются поперечнополосатые кардиомиоциты
Границами клеток являются вставочные пластинки,

Сердечная мышечная ткань
Цепочка кардиомиоцитов образует мышечные волокна
Волокна соединяются анастомозами (приобретают сетчатую
Сердечная мышечная ткань
Цепочка кардиомиоцитов образует мышечные волокна
Волокна соединяются анастомозами (приобретают сетчатую
Сердечная поперечнополосатая мышечная ткань
Сердечная поперечнополосатая мышечная ткань
 Обогащение словаря детей в процессе расширения представлений о разных способах передачи сообщений
Обогащение словаря детей в процессе расширения представлений о разных способах передачи сообщений Психолого-педагогическое сопровождение старших дошкольников при подготовке их к школе
Психолого-педагогическое сопровождение старших дошкольников при подготовке их к школе Техническое обслуживание магистральных газопроводов
Техническое обслуживание магистральных газопроводов Интегративный подход к организации развивающих центров активности детей
Интегративный подход к организации развивающих центров активности детей Формирование у детей основы правосознания и культуры в области дорожного движения через использование разных форм взаимовоздействия педагогов с дошкольниками.
Формирование у детей основы правосознания и культуры в области дорожного движения через использование разных форм взаимовоздействия педагогов с дошкольниками. Силачи и слабаки в мире кислот и оснований.
Силачи и слабаки в мире кислот и оснований. Заочное производство. Понятие и порядок проведения
Заочное производство. Понятие и порядок проведения Классный час-презентация Моя Родина.
Классный час-презентация Моя Родина. презентация Мини - музей средних и младших группах детского сада № 26 Сказка
презентация Мини - музей средних и младших группах детского сада № 26 Сказка Алгоритм и его формальное исполнение
Алгоритм и его формальное исполнение Гидросфера. Свойства воды. Три состояния воды
Гидросфера. Свойства воды. Три состояния воды Classroom language. Instructions
Classroom language. Instructions 11класс. Германия. Диск
11класс. Германия. Диск Наша Тула.
Наша Тула. Почему дети лгут?
Почему дети лгут? Кроссворд с применением национально-регионального компонента
Кроссворд с применением национально-регионального компонента Предмет и задачи регионоведения
Предмет и задачи регионоведения Корпоративные стратегии диверсификации
Корпоративные стратегии диверсификации Памятка православного христианина. О некоторых основах и правилах церковной жизни
Памятка православного христианина. О некоторых основах и правилах церковной жизни Ударные и безударные гласные звуки
Ударные и безударные гласные звуки презентация обобщение знаний по теме Вселенная
презентация обобщение знаний по теме Вселенная Презентация к уроку Роль гидросферы в жизни Земли часть1 7 класс
Презентация к уроку Роль гидросферы в жизни Земли часть1 7 класс Презентация Портфолио воспитателя
Презентация Портфолио воспитателя Человек и природа
Человек и природа Линейная алгебра
Линейная алгебра Многоугольники. Четырехугольники. 8 класс
Многоугольники. Четырехугольники. 8 класс Организация работы ПМПк и ППк в дошкольных образовательных учреждениях
Организация работы ПМПк и ППк в дошкольных образовательных учреждениях Физиология больного организма
Физиология больного организма