- Образование микроспоры и микрогаметофита

Содержание
- 2. Смена поколений Начало гаплоидному поколению растений дает мейотическое деление диплоидных спорогенных клеток. При этом образуются споры,
- 3. Особенности физиологии гаметофитного поколения определяется его местом в жизненном цикле растений. Гаметофиты у семенных растений зависят
- 4. Образование микрогаметофита
- 5. Аспекты физиологии мужского гаметофита Пыльник. Формирование, поляризация и деление микроспоры, формирование оболочки Метаболизм, транскриптом и протеом
- 6. Пыльник. Тапетум. У цветковых растений образование гаплоидных микроспор и развитие пыльцевых зерен происходит в пыльнике. В
- 7. Водный статус пыльника В качестве одного из главных механизмов обезвоживания рассматривают перераспределение воды в пыльнике и
- 8. Гормональный контроль Для развития тычинки и раскрытия пыльников нужны фитогормоны. Наиболее изучен ауксин. Его концентрация в
- 9. Одновременно увеличивается экспрессия генов, контролирующих синтез жасмоновой кислоты. Последующая дегидратация эндотеция приводит к деформации пыльникаиз-за того,
- 10. Тем временем в гнезде пыльника…
- 11. Формирование тетрады После мейоза микроспороцита четыре гаплоидные клетки вначале остаются под общей каллозной оболочкой, образуя тетрады
- 12. Особенности и синтез каллозы Характерная особенность каллозы – быстрый синтез и быстрая деградация Этот полисахарид играет
- 13. Зачем же нужна каллоза? В ранних работах обсуждали две главные функции каллозной оболочки: во-первых, физическая изоляция
- 14. Экзина и апертуры Экзина, наружная оболочка пыльцевого зерна, защищает гаметофит от воздействия неблагоприятных факторов внешней среды
- 15. Строение экзины Экзина у многих растений имеет уникальный рельеф. Обычно она состоит из двух слоев: наружный
- 16. Спорополленин Спорополленин обеспечивает не только механическую прочность экзины, но и ее высокий антиоксидантный потенциал. Это свойство
- 17. Формирование экзины Образование экзины начинается, когда микроспора еще находится в тетраде под толстой каллозной оболочкой и
- 18. Секреция предшественников СП Для секреции клетками тапетума нужны белки, связанные с плазмалеммой. Эту роль отводят ABC
- 19. Общая схема строительства экзины Proposed Model for the Localization of Pollen Wall Components within Anther Tissues.
- 20. Тельца Убиша Они были открыты около 150 лет назад и обнаружены в пыльниках более 300 видов
- 21. Накопление компонентов СП
- 22. Полимеризация спорополленина Пока обнаружен лишь один из возможных механизмов полимеризации, в основе которого лежат использование активных
- 23. Формирование уникального рельефа экзины Согласно актуальной модели, матрикс примэкзины гидратируется в периплазматическом пространстве микроспоры и, с
- 24. Самосборка экзины Другие авторы рассматривают процесс самосборки экзины с позиций коллоидной химии. В рамках этой концепции
- 25. Растворение и выход из тетрады В растворении каллозной оболочки тетрады участвует каллаза – комплекс β-1,3-глюканаз (экзо-
- 26. Созревание микроспоры В микроспоре растет крупная вакуоль, а ядро перемещается на периферию. Микроспора растет практически без
- 27. Миграция ядра Ядро в премитозной микроспоре прижато к оболочке. Миграция ядра начинается раньше, чем вакуолизация, поэтому
- 28. Механизмы поляризации Механизмы поляризации микроспоры все еще мало изучены, однако анализ немногочисленных мутантов показал, что важную
- 29. Деление микроспоры Поляризованная микроспора вступает в асимметричный митоз (I пыльцевой митоз). Он приводит к образованию пыльцевого
- 30. Формирование интины Отложение интины начинается до или после митоза микроспоры, как правило, сначала вблизи апертуры, а
- 31. Нарушения формирования интины Число мутантов с нарушениями развития интины сравнительно невелико. Как правило, они затрагивают гены
- 32. Пыльцевое зерно Двухклеточное пыльцевое зерно цветковых растений – это уникальная конструкция, построенная по типу «клетка-в-клетке». Почти
- 33. Деления при созревании гаметофита Покрытосеменные: 25% 3-клеточных (арабидопсис), 75% 2-клеточных (лилия, табак). Микрогаметофит голосеменных претерпевает 3-5
- 34. «Ранние» и «поздние» Экспрессия «ранних» генов начинается в микроспоре и заметно снижается перед созреванием пыльцевого зерна.
- 35. Какие мРНК и белки запасаются После деления микроспоры идет накопление пула долгоживущих мРНК, которые не транслируются
- 37. Скачать презентацию

Смена поколений
Начало гаплоидному поколению растений дает мейотическое деление диплоидных спорогенных клеток.
Смена поколений
Начало гаплоидному поколению растений дает мейотическое деление диплоидных спорогенных клеток.

Особенности физиологии гаметофитного поколения определяется его местом в жизненном цикле растений.
Гаметофиты
Особенности физиологии гаметофитного поколения определяется его местом в жизненном цикле растений.
Гаметофиты
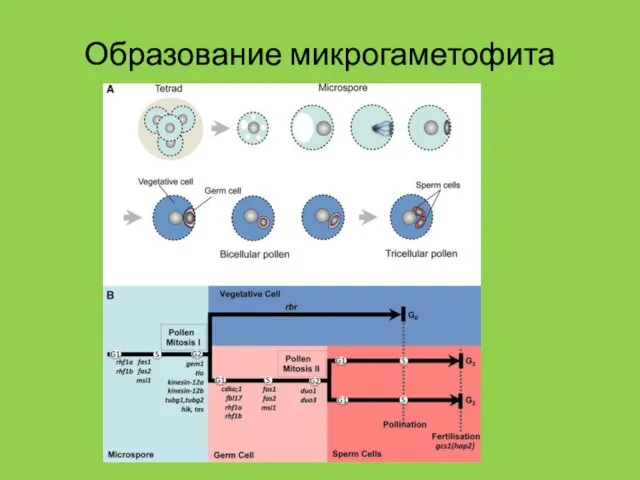
Образование микрогаметофита
Образование микрогаметофита

Аспекты физиологии мужского гаметофита
Пыльник. Формирование, поляризация и деление микроспоры, формирование оболочки
Метаболизм,
Аспекты физиологии мужского гаметофита
Пыльник. Формирование, поляризация и деление микроспоры, формирование оболочки
Метаболизм,
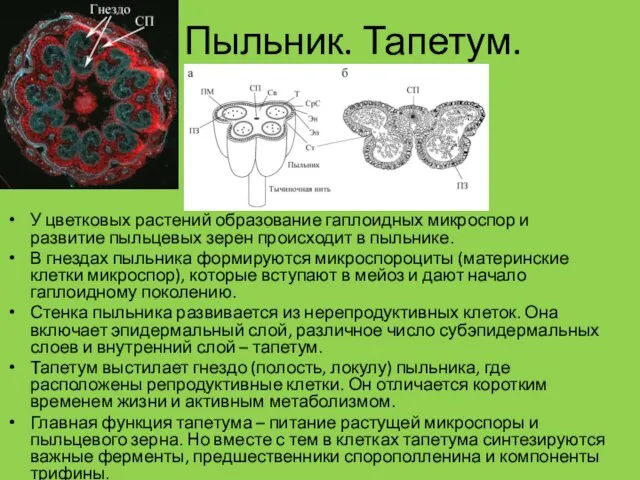
Пыльник. Тапетум.
У цветковых растений образование гаплоидных микроспор и развитие пыльцевых зерен
Пыльник. Тапетум.
У цветковых растений образование гаплоидных микроспор и развитие пыльцевых зерен

Водный статус пыльника
В качестве одного из главных механизмов обезвоживания рассматривают перераспределение
Водный статус пыльника
В качестве одного из главных механизмов обезвоживания рассматривают перераспределение
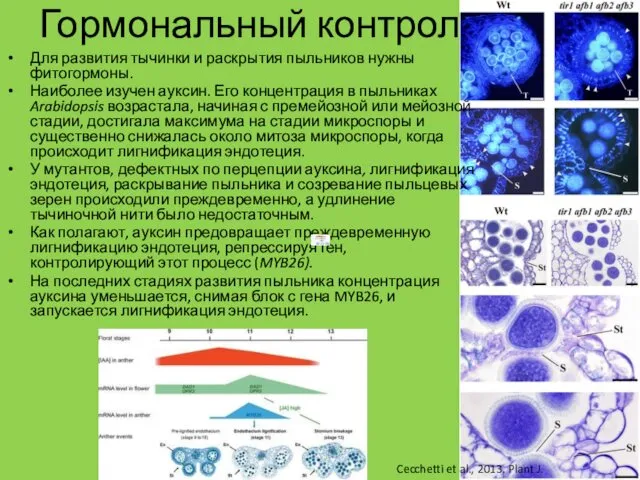
Гормональный контроль
Для развития тычинки и раскрытия пыльников нужны фитогормоны.
Наиболее изучен
Гормональный контроль
Для развития тычинки и раскрытия пыльников нужны фитогормоны.
Наиболее изучен
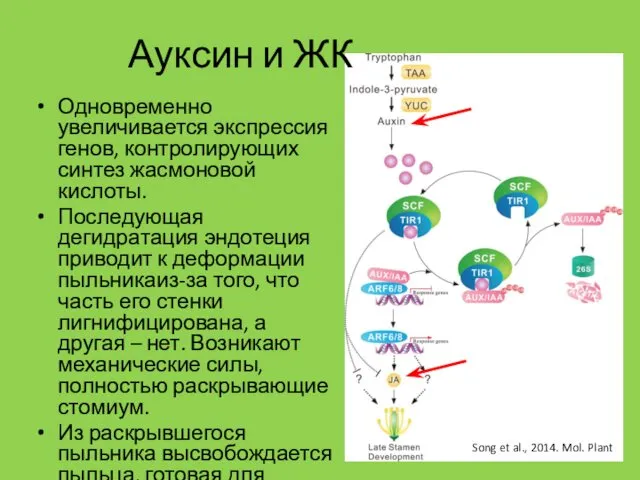
Одновременно увеличивается экспрессия генов, контролирующих синтез жасмоновой кислоты.
Последующая дегидратация эндотеция
Одновременно увеличивается экспрессия генов, контролирующих синтез жасмоновой кислоты.
Последующая дегидратация эндотеция

Тем временем в гнезде пыльника…
Тем временем в гнезде пыльника…
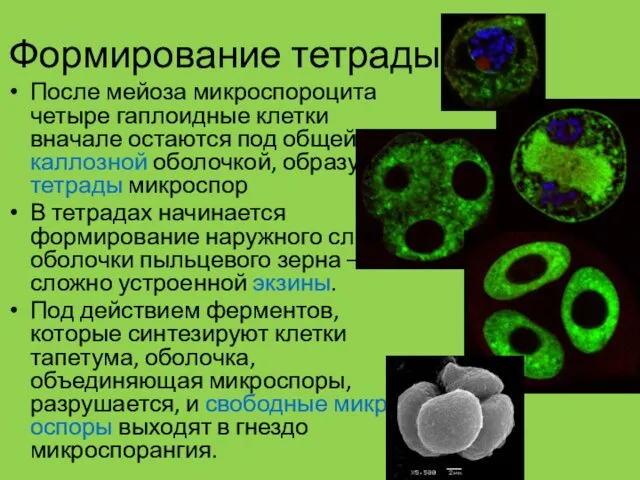
Формирование тетрады
После мейоза микроспороцита четыре гаплоидные клетки вначале остаются под общей
Формирование тетрады
После мейоза микроспороцита четыре гаплоидные клетки вначале остаются под общей
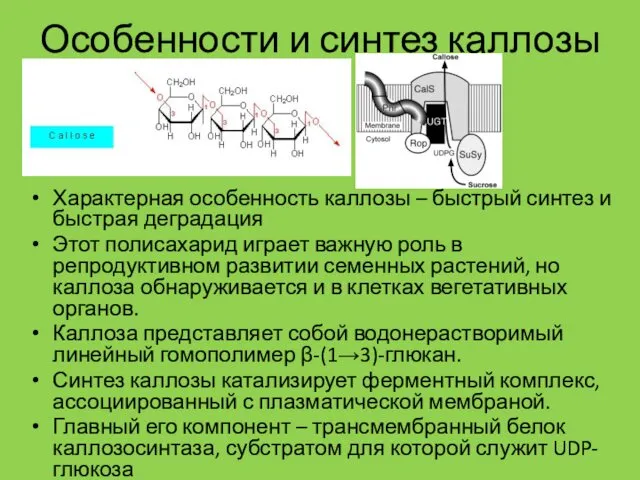
Особенности и синтез каллозы
Характерная особенность каллозы – быстрый синтез и быстрая
Особенности и синтез каллозы
Характерная особенность каллозы – быстрый синтез и быстрая

Зачем же нужна каллоза?
В ранних работах обсуждали две главные функции каллозной
Зачем же нужна каллоза?
В ранних работах обсуждали две главные функции каллозной
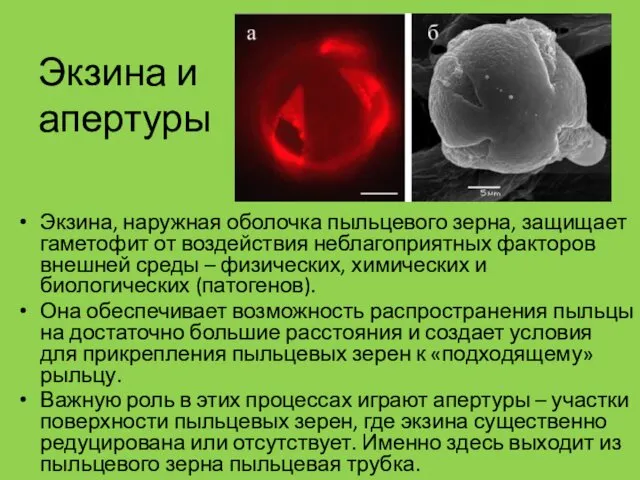
Экзина и апертуры
Экзина, наружная оболочка пыльцевого зерна, защищает гаметофит от воздействия
Экзина и апертуры
Экзина, наружная оболочка пыльцевого зерна, защищает гаметофит от воздействия
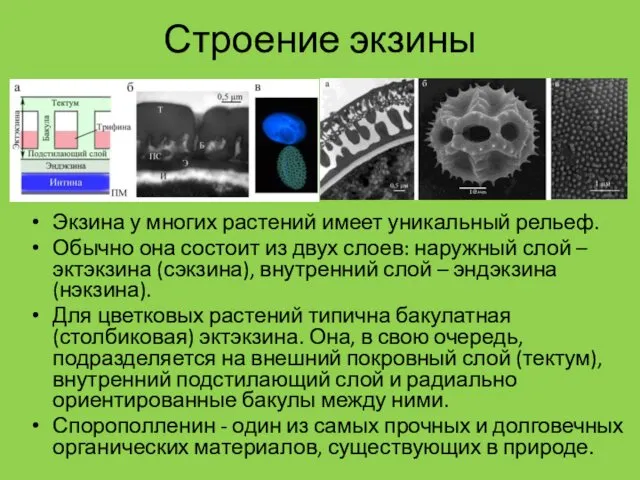
Строение экзины
Экзина у многих растений имеет уникальный рельеф.
Обычно она состоит из
Строение экзины
Экзина у многих растений имеет уникальный рельеф.
Обычно она состоит из
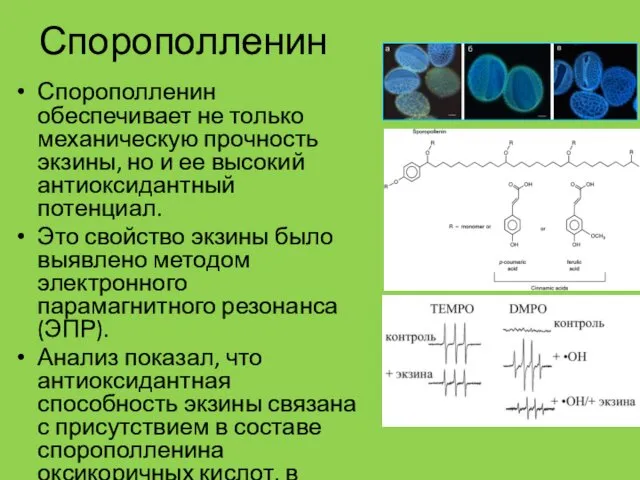
Спорополленин
Спорополленин обеспечивает не только механическую прочность экзины, но и ее высокий
Спорополленин
Спорополленин обеспечивает не только механическую прочность экзины, но и ее высокий

Формирование экзины
Образование экзины начинается, когда микроспора еще находится в тетраде под
Формирование экзины
Образование экзины начинается, когда микроспора еще находится в тетраде под
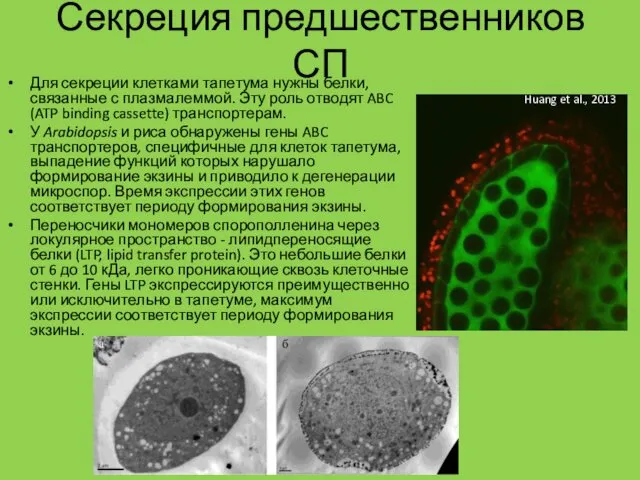
Секреция предшественников СП
Для секреции клетками тапетума нужны белки, связанные с плазмалеммой.
Секреция предшественников СП
Для секреции клетками тапетума нужны белки, связанные с плазмалеммой.
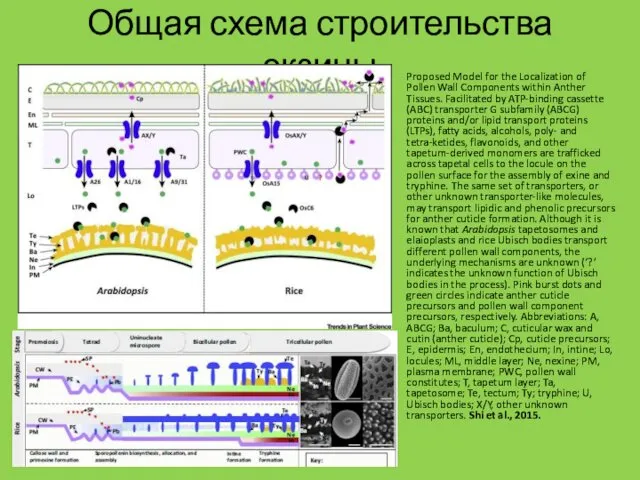
Общая схема строительства экзины
Proposed Model for the Localization of Pollen Wall
Общая схема строительства экзины
Proposed Model for the Localization of Pollen Wall
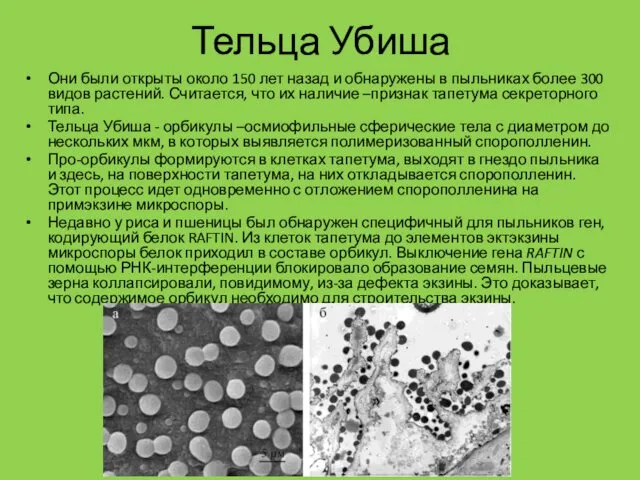
Тельца Убиша
Они были открыты около 150 лет назад и обнаружены в
Тельца Убиша
Они были открыты около 150 лет назад и обнаружены в
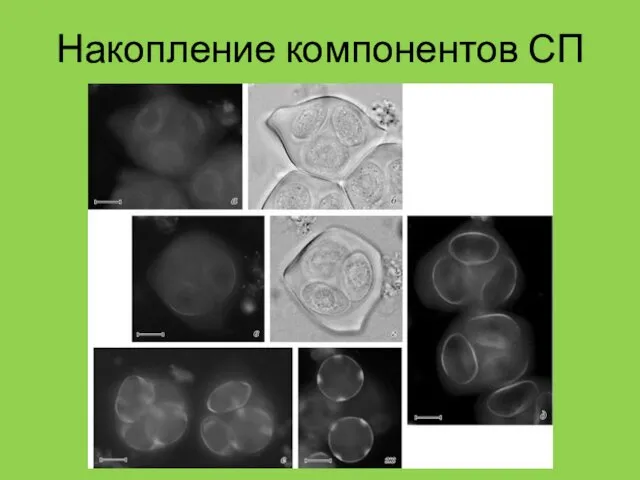
Накопление компонентов СП
Накопление компонентов СП
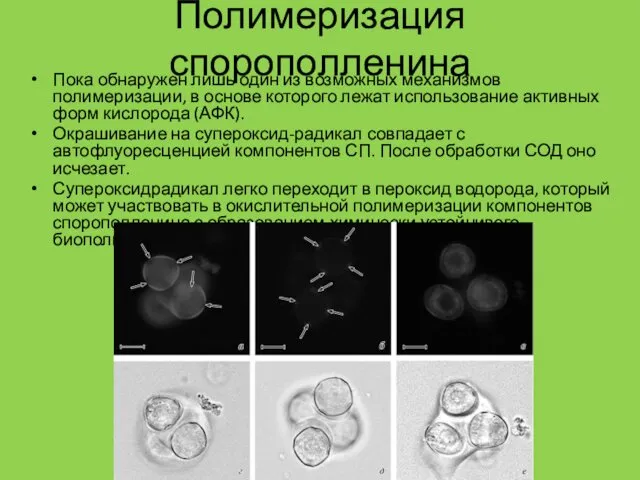
Полимеризация спорополленина
Пока обнаружен лишь один из возможных механизмов полимеризации, в основе
Полимеризация спорополленина
Пока обнаружен лишь один из возможных механизмов полимеризации, в основе

Формирование уникального рельефа экзины
Согласно актуальной модели, матрикс примэкзины гидратируется в периплазматическом
Формирование уникального рельефа экзины
Согласно актуальной модели, матрикс примэкзины гидратируется в периплазматическом

Самосборка экзины
Другие авторы рассматривают процесс самосборки экзины с позиций коллоидной химии.
Самосборка экзины
Другие авторы рассматривают процесс самосборки экзины с позиций коллоидной химии.
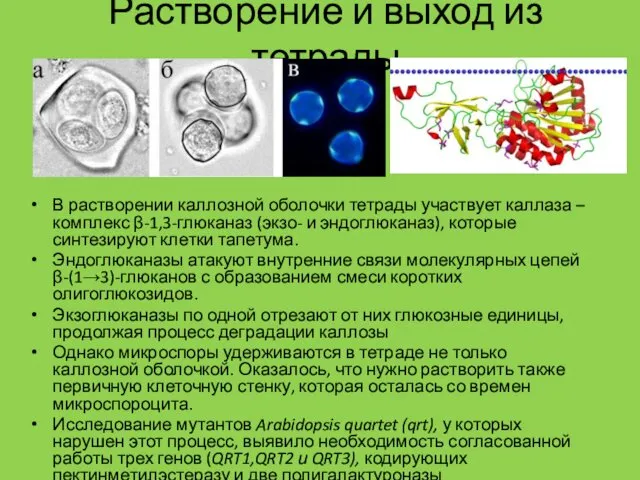
Растворение и выход из тетрады
В растворении каллозной оболочки тетрады участвует каллаза
Растворение и выход из тетрады
В растворении каллозной оболочки тетрады участвует каллаза
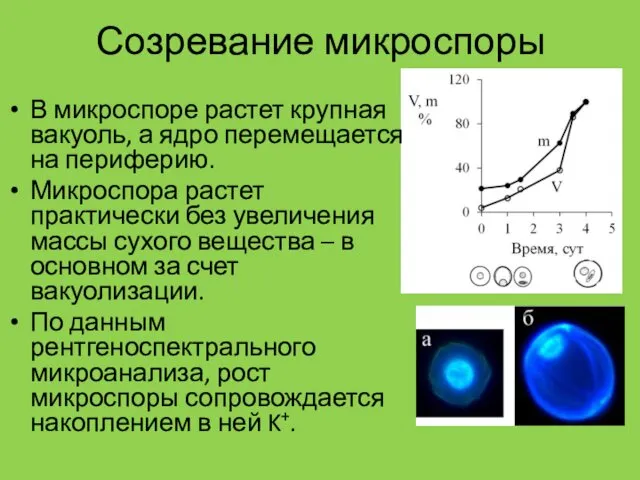
Созревание микроспоры
В микроспоре растет крупная вакуоль, а ядро перемещается на периферию.
Микроспора
Созревание микроспоры
В микроспоре растет крупная вакуоль, а ядро перемещается на периферию.
Микроспора
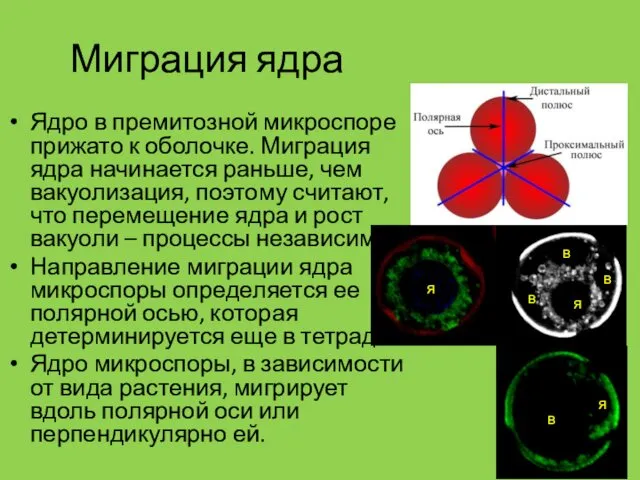
Миграция ядра
Ядро в премитозной микроспоре прижато к оболочке. Миграция ядра начинается
Миграция ядра
Ядро в премитозной микроспоре прижато к оболочке. Миграция ядра начинается
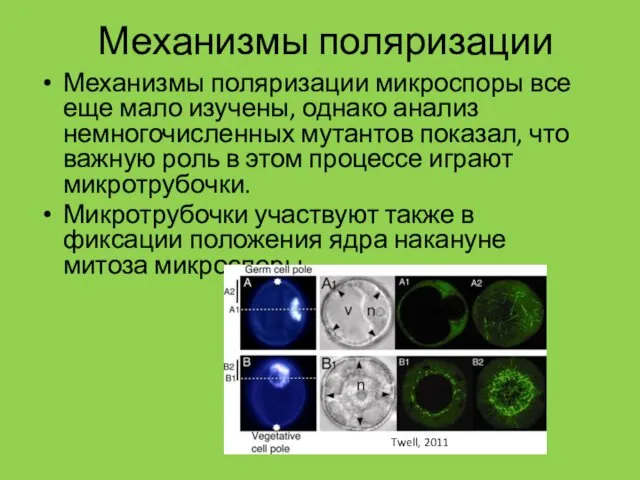
Механизмы поляризации
Механизмы поляризации микроспоры все еще мало изучены, однако анализ немногочисленных
Механизмы поляризации
Механизмы поляризации микроспоры все еще мало изучены, однако анализ немногочисленных

Деление микроспоры
Поляризованная микроспора вступает в асимметричный митоз (I пыльцевой митоз). Он
Деление микроспоры
Поляризованная микроспора вступает в асимметричный митоз (I пыльцевой митоз). Он
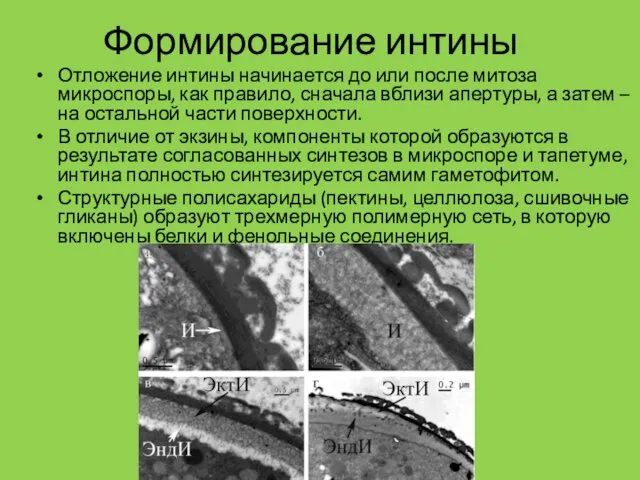
Формирование интины
Отложение интины начинается до или после митоза микроспоры, как правило,
Формирование интины
Отложение интины начинается до или после митоза микроспоры, как правило,

Нарушения формирования интины
Число мутантов с нарушениями развития интины сравнительно невелико. Как
Нарушения формирования интины
Число мутантов с нарушениями развития интины сравнительно невелико. Как
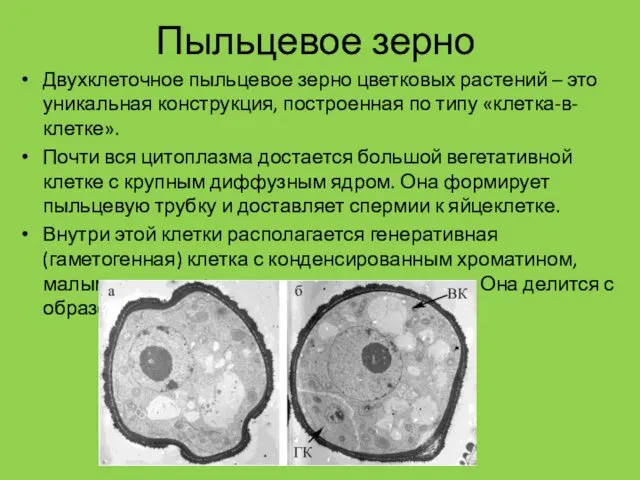
Пыльцевое зерно
Двухклеточное пыльцевое зерно цветковых растений – это уникальная конструкция, построенная
Пыльцевое зерно
Двухклеточное пыльцевое зерно цветковых растений – это уникальная конструкция, построенная
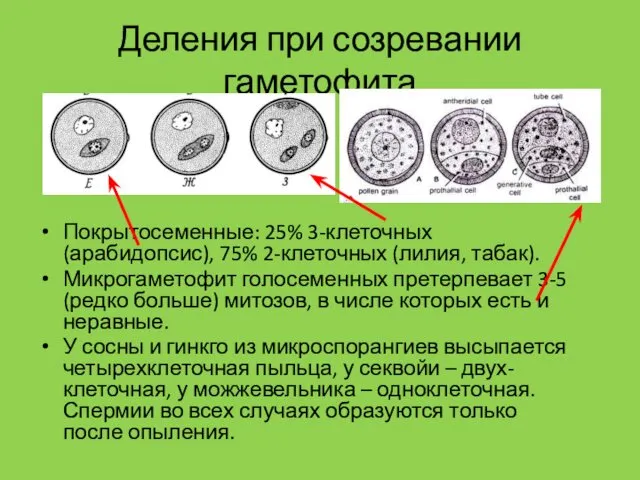
Деления при созревании гаметофита
Покрытосеменные: 25% 3-клеточных (арабидопсис), 75% 2-клеточных (лилия, табак).
Микрогаметофит
Деления при созревании гаметофита
Покрытосеменные: 25% 3-клеточных (арабидопсис), 75% 2-клеточных (лилия, табак).
Микрогаметофит

«Ранние» и «поздние»
Экспрессия «ранних» генов начинается в микроспоре и заметно снижается
«Ранние» и «поздние»
Экспрессия «ранних» генов начинается в микроспоре и заметно снижается

Какие мРНК и белки запасаются
После деления микроспоры идет накопление пула долгоживущих
Какие мРНК и белки запасаются
После деления микроспоры идет накопление пула долгоживущих
 Культура Эллинизма
Культура Эллинизма 1С Парфюм Логистик
1С Парфюм Логистик Применение нетрадиционных источников энергии в Чувашской Республике
Применение нетрадиционных источников энергии в Чувашской Республике zanyatie_10_pro_zaychat_i_zaychihu
zanyatie_10_pro_zaychat_i_zaychihu Джон Эверетт Милле (John Everett Millais)
Джон Эверетт Милле (John Everett Millais) Мои земляки - труженики полей
Мои земляки - труженики полей Перу. План описания страны Латинской Америки
Перу. План описания страны Латинской Америки Имя прилагательное.Суффиксы прилагательных
Имя прилагательное.Суффиксы прилагательных Культура ислама. Основы духовно – нравственной культуры народов России
Культура ислама. Основы духовно – нравственной культуры народов России Что такое дисграфия и как с ней бороться?
Что такое дисграфия и как с ней бороться? Секреты семейного воспитания
Секреты семейного воспитания Презентация Игра в жизни ребенка
Презентация Игра в жизни ребенка E-learning и дистанционные технологии
E-learning и дистанционные технологии Microsoft Office Power Point
Microsoft Office Power Point Луч и угол
Луч и угол Глобальная энергетическая и сырьевая проблема
Глобальная энергетическая и сырьевая проблема Система мотивации за инновационные проекты
Система мотивации за инновационные проекты Тема поэта и поэзии в лирике А.С. Пушкина
Тема поэта и поэзии в лирике А.С. Пушкина Шляпный этикет, как правильно вписать аксессуар из прошлого в современный образ
Шляпный этикет, как правильно вписать аксессуар из прошлого в современный образ Кабинет географии
Кабинет географии Кроссворд Что мы знаем о пчёлах
Кроссворд Что мы знаем о пчёлах Органы растений. Побег
Органы растений. Побег Газообразное состояние вещества
Газообразное состояние вещества Гидроэлектростанции. Типы ГЭС
Гидроэлектростанции. Типы ГЭС Жигули. ОАО АвтоВАЗ
Жигули. ОАО АвтоВАЗ Отчет по искусствоведческой практике. Шаблон
Отчет по искусствоведческой практике. Шаблон Презентация Интеллектуальное развитие дошкольников
Презентация Интеллектуальное развитие дошкольников Винсент Ван Гог - краткая биография и картины художника
Винсент Ван Гог - краткая биография и картины художника