- Ощущения с поверхности тела

Содержание
- 2. механорецепция, терморецепция и ноцицепция ( болевая чувствительность) Соматовисцеральная чувствительность Типы рецепторов кожи: А — тельце Пачини;
- 3. Рецепторный аппарат кожи 1 – нервное сплетение луковицы волоса; 2 – свободные нервные окончания; 3 –
- 4. Механорецепторы голой (А) и волосистой (В) кожи человека 1 — меланоцит, 2 — герминативный слой, 3
- 5. Строение тельца Пачини (А) и его электрические реакции на механическое раздражение (Б) I — без поляризации
- 6. Схема пластинчатого тельца Фатера - Пачини: А - продольный срез: 1 - соединительнотканная капсула; 2, 5,
- 7. В ответ на механический стимул в тельце Пачини возникает РП (рецепторный потенциал), который распространяется вдоль нервного
- 8. диск Меркеля тельце Мейсснера
- 10. Свободные нервные окончания в соединительной ткани. Осевой цилиндр контактирует с вспомогательными клетками на значительном протяжении или
- 11. Мышечная и суставная рецепция (проприорецепция). В мышцах млекопитающих животных и человека содержится три типа специализированных рецепторов:
- 12. Мышечные веретена. Впервые в ряду позвоночных мышечные веретена появляются у амфибий. Предполагают, что они возникли для
- 13. Строение мышечного веретена лягушки (А) и средней части мышечного веретена млекопитающих (Б) 1 — моторный аксон,
- 14. У птиц и млекопитающих в строении мышечных рецепторов наблюдается ряд прогрессивных черт. Интрафузальные волокна в мышечных
- 15. Мышечное веретено. 1 — проксимальный конец интрафузального мышечного волокна, прикрепленного к волокну скелетной мышцы; 2 —
- 16. Эфферентная иннервация скелетной мышцы: 1 — сухожилие мышцы; 2 — сухожильные рецепторы Гольджи; 3 — свободные
- 18. Скачать презентацию

механорецепция,
терморецепция и
ноцицепция ( болевая чувствительность)
Соматовисцеральная чувствительность
Типы рецепторов кожи: А
механорецепция,
терморецепция и
ноцицепция ( болевая чувствительность)
Соматовисцеральная чувствительность
Типы рецепторов кожи: А
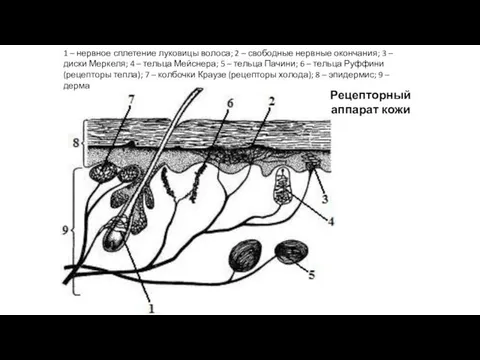
Рецепторный аппарат кожи
1 – нервное сплетение луковицы волоса; 2 – свободные
Рецепторный аппарат кожи
1 – нервное сплетение луковицы волоса; 2 – свободные
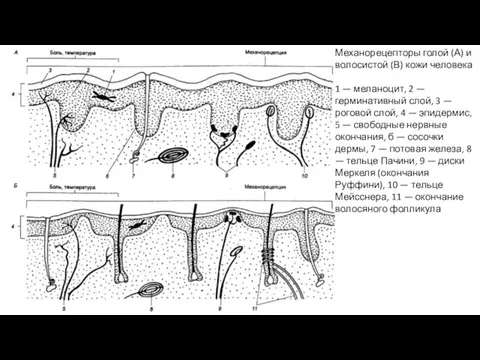
Механорецепторы голой (А) и волосистой (В) кожи человека
1 — меланоцит, 2
Механорецепторы голой (А) и волосистой (В) кожи человека
1 — меланоцит, 2

Строение тельца Пачини (А) и его электрические реакции на механическое
Строение тельца Пачини (А) и его электрические реакции на механическое
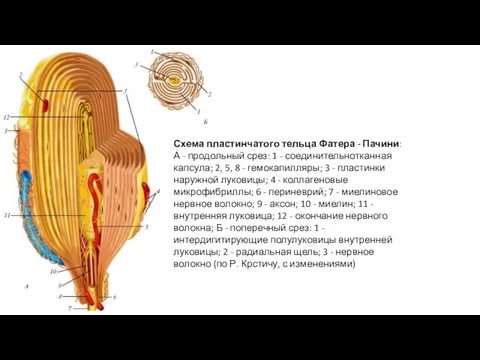
Схема пластинчатого тельца Фатера - Пачини:
А - продольный срез: 1
Схема пластинчатого тельца Фатера - Пачини:
А - продольный срез: 1

В ответ на механический стимул в тельце Пачини возникает РП (рецепторный
В ответ на механический стимул в тельце Пачини возникает РП (рецепторный
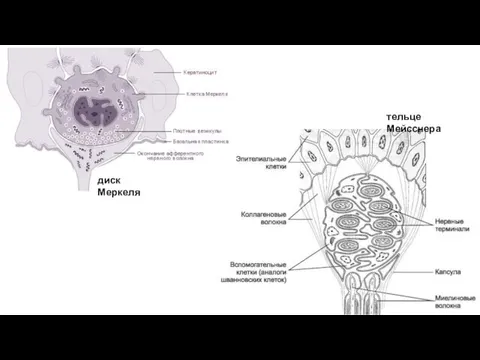
диск Меркеля
тельце Мейсснера
диск Меркеля
тельце Мейсснера
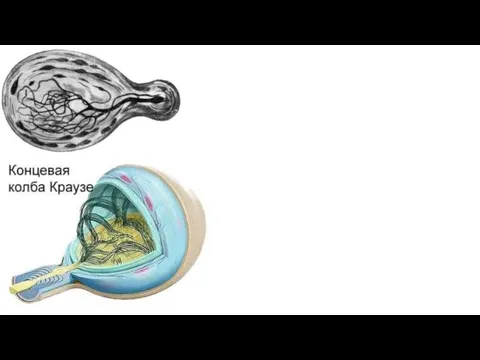
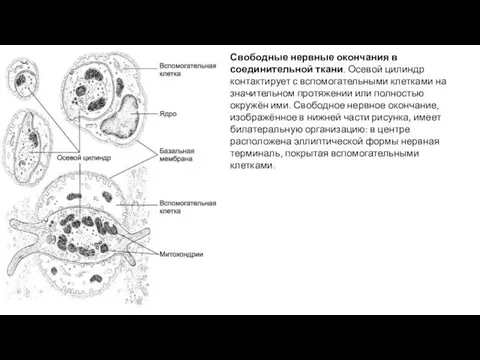
Свободные нервные окончания в соединительной ткани. Осевой цилиндр контактирует с вспомогательными
Свободные нервные окончания в соединительной ткани. Осевой цилиндр контактирует с вспомогательными

Мышечная и суставная рецепция (проприорецепция). В мышцах млекопитающих животных и человека содержится
Мышечная и суставная рецепция (проприорецепция). В мышцах млекопитающих животных и человека содержится

Мышечные веретена. Впервые в ряду позвоночных мышечные веретена появляются у амфибий.
Мышечные веретена. Впервые в ряду позвоночных мышечные веретена появляются у амфибий.

Строение мышечного веретена лягушки (А) и средней части мышечного веретена млекопитающих
Строение мышечного веретена лягушки (А) и средней части мышечного веретена млекопитающих
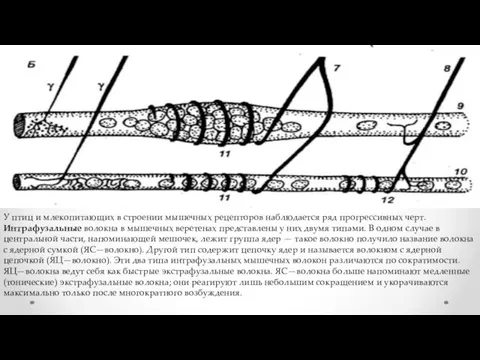
У птиц и млекопитающих в строении мышечных рецепторов наблюдается ряд прогрессивных
У птиц и млекопитающих в строении мышечных рецепторов наблюдается ряд прогрессивных
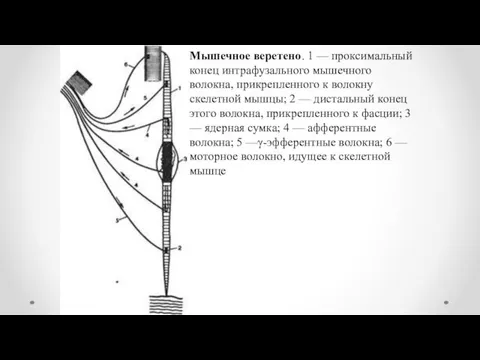
Мышечное веретено. 1 — проксимальный конец интрафузального мышечного волокна, прикрепленного к
Мышечное веретено. 1 — проксимальный конец интрафузального мышечного волокна, прикрепленного к

Эфферентная иннервация скелетной мышцы:
1 — сухожилие мышцы; 2 — сухожильные рецепторы
Эфферентная иннервация скелетной мышцы:
1 — сухожилие мышцы; 2 — сухожильные рецепторы
 Советы по внешнему оформлению презентации
Советы по внешнему оформлению презентации День из жизни водохранилища
День из жизни водохранилища Целевой рынок. Тема 9
Целевой рынок. Тема 9 Базовый курс по напиткам
Базовый курс по напиткам Дидактическая игра для детей 3-4 лет Помоги зайке
Дидактическая игра для детей 3-4 лет Помоги зайке Шаблоны презентаций Синие
Шаблоны презентаций Синие Первая русская революция. 9 класс
Первая русская революция. 9 класс Европейский Север
Европейский Север Особенности и правила приёмной кампании - 2020 в вятское художественное училище имени А.А. Рылова
Особенности и правила приёмной кампании - 2020 в вятское художественное училище имени А.А. Рылова Служение Отечеству императора Николая II: хозяин Земли Русской
Служение Отечеству императора Николая II: хозяин Земли Русской Использование ИКТ на уроках географии линии Сфера как средство формирования информационной компетентности учащихся
Использование ИКТ на уроках географии линии Сфера как средство формирования информационной компетентности учащихся Регистрация на сайте Госуслуги.ру или Единая система идентификации и аутентификации (ЕСИА)
Регистрация на сайте Госуслуги.ру или Единая система идентификации и аутентификации (ЕСИА) Семинар - теоретический Контроль в дошкольном образовательном учреждении.
Семинар - теоретический Контроль в дошкольном образовательном учреждении. Пісіру, дәнекерлеу, желімдеу
Пісіру, дәнекерлеу, желімдеу Крапивин Владислав Петрович
Крапивин Владислав Петрович Морфологическая характеристика лекарственных и ядовитых растений
Морфологическая характеристика лекарственных и ядовитых растений Формирование нравственно-патриотических качеств у младших школьников.
Формирование нравственно-патриотических качеств у младших школьников. Екатерина Хохрина искитимское озеро 26_02_2022
Екатерина Хохрина искитимское озеро 26_02_2022 Язык программирования Pascal
Язык программирования Pascal Интернет - ресурсы в работе учителя как один из способов повышения педагогического мастерства
Интернет - ресурсы в работе учителя как один из способов повышения педагогического мастерства Подготовка к аккредитации
Подготовка к аккредитации Componentele hardware ale unui calculator
Componentele hardware ale unui calculator Презентация к уроку химии по теме: Соединения железа
Презентация к уроку химии по теме: Соединения железа Интеллектуальная викторина Своя игра
Интеллектуальная викторина Своя игра Форма нігтя, розповість все про твій характер
Форма нігтя, розповість все про твій характер Перспективные направления биотехнологии
Перспективные направления биотехнологии Еще одна прекрасная церковь_БОУ СОШ6
Еще одна прекрасная церковь_БОУ СОШ6 Презентация по технологии
Презентация по технологии