- Палеогеография и история биоты четвертичного периода Западной Сибири

Содержание
- 2. Лаборатория филогенетики и биохронологии ИЭРиЖ УрО РАН Лаборатория Филогенетики и биохронологии образована 1 апреля 2012 года.
- 3. Лаборатория филогенетики и биохронологии ИЭРиЖ УрО РАН исследования в рамках четырех основных направлений: 1 - Исследование
- 4. Лаборатория филогенетики и биохронологии ИЭРиЖ УрО РАН исследования в рамках четырех основных направлений: 2 - Исследование
- 5. Лаборатория филогенетики и биохронологии ИЭРиЖ УрО РАН исследования в рамках четырех основных направлений: 3 - Исследование
- 6. Лаборатория филогенетики и биохронологии ИЭРиЖ УрО РАН исследования в рамках четырех основных направлений: 4 - Анализ
- 7. Геохронология. Возраст Земли 4,6 млрд лет
- 8. Эмпайр-стейт-билдинг 443,2 м Геохронология. Возраст Земли 4,6 млрд лет
- 9. Эмпайр-стейт-билдинг 443,2 м Геохронология. Возраст Земли 4,6 млрд лет
- 10. Зачем нужны архивы? - реконструировать историю Зачем необходимо знание истории? - для построения прогностической модели
- 11. Зачем изучать геологическую историю и историю биоты? "Тот, кто не помнит своего прошлого, обречен на то,
- 12. "Тот, кто не помнит своего прошлого, обречен на то, чтобы пережить его вновь". Джордж Сантаяна (1863
- 13. THE PRESENT IS THE KEY TO THE PAST IS THE KEY TO THE FUTURE (MATHIESON, Elizabeth
- 14. THE PRESENT IS THE KEY TO THE PAST IS THE KEY TO THE FUTURE НАСТОЯЩЕЕ ОТКРЫВАЕТ
- 15. Требования к архивам: Доступность Информативность Интерпретируемость
- 16. Тафоно́мия (от греч. τάφος — «могила, погребение», и νόμος — «закон») —раздел палеонтологии и археологии, изучающий
- 17. Мы будем говорить об изучении геологической истории и истории биоты четвертичного периода – это примерно 2,5
- 18. Архивы истории биоты четвертичного периода работают, прежде всего, на биостратиграфию и биохронологию
- 19. The West Siberian Plain is one of the largest plains in the World providing opportunity to
- 20. В настоящее время во всех научных науках, связанных с изучением четвертичного периода очевидна тенденция сопоставления региональных
- 21. Один из удачных примеров трансрегиональных стратиграфических корреляций:
- 22. Вот списоке регионов, включенных в это исследование, Западно-Сибирская равнина отсутствует
- 23. Причина кроется в неоднозначности интерпретаций палеогеографической истории и генезиса отложений, в частности, в северной части равнины
- 24. В настоящее время Западно-Сибирская равнина характеризуется ярко выраженной широтной зональностью, начиная от арктических пустынь на севере
- 25. В настоящее время Западно-Сибирская равнина характеризуется ярко выраженной широтной зональностью, начиная от арктических пустынь на севере
- 26. Но в этом регионе также существуют явно разные зоны формирования четвертичных отложений, которые традиционно подразделяются на
- 27. Когда корреляция, основанная на небиологических данных, кажется проблематичной, роль биостратиграфии значительно возрастает В то же время
- 28. В Северном полушарии полевки представляют группу млекопитающих, которая наиболее широко используется в биостратиграфических и биохронологических целях
- 29. Mammuth incessor Rodent incessor Для многих территорий основой биостратиграфических построений являются крупные млекопитающие. Для Западно-Сибирской равнины
- 30. Разрезы четвертичных отложений, доступные для исследования, часто дают богатые скопления макроостатков растений, жесткокрылых и мелких млекопитающих.
- 31. Объекты исследований В рамках палеонтологического подхода на первом этапе работ объектом исследований были местонахождения ископаемых остатков
- 32. Типы плейстоценовых и голоценовых местонахождений Урала и Западной Сибири Три аспекта, имеющие решающее значение при исследовании
- 33. Местонахождения в скальных массивах Одно из преимуществ пещерных отложений в том, что местонахождения, приуроченные к ним,
- 34. Cпелеогенные отложения карстовые полости К этому типу местонахождений относятся пещеры, гроты, скальные навесы в карстующихся скальных
- 35. Отложения некарстовых гротов К этому типу относятся неглубокие полости в скальных массивах с преобладанием базальтовых и
- 36. Аллювиально- спелеогенные отложения К этому типу местонахождений относятся пещеры, отложения которых формировались как за счет процессов
- 37. Факторы накопления и источников попадания костей в пещеры Накопление, распределение и консервация костей в отложениях пещер
- 38. Местонахождения равнинной части Зауралья и Западной Сибири Карьеры и береговые обрывы рек и озер – основные
- 39. Отложения, вскрытые по берегам рек и в карьерах являются основным источником информации о фаунах насекомых. Именно
- 40. Кроме того, есть несколько дополнительных источников материалов для голоценовых отложений : торфяники, донные отложения и норовища
- 41. Торфяники Озерно-болотные отложения, ручное бурение, послойное изучение кернов на фото: керн из скважины торфяника, голоцен, Пермское
- 42. Одним из источников информации для изучения четвертичных насекомых являются отложения погребенных торфяников плейстоценового и голоценового возраста
- 43. Голоценовые торфяники широко распространены на территории Урала и Западной Сибири. В них содержатся остатки насекомых, однако
- 44. Археологические памятники
- 45. Остатки насекомых из археологических памятников активно изучаются в Западной Европе и Северной Америке. Разработан даже специальный
- 46. Археологические памятники Культурный слой средневекового поселения, археологический раскоп жилища, зольник около очага на фото: Рождественское городище,
- 47. Реконструкция последней диеты животного, его пастбищних стаций, условий палеосреды обитания Туши крупных млекопитающих сохранившихся в условиях
- 48. На фото: казахский энтомолог-четвертичник Ф.Г.Бидашко (Уральск, Казахстан) исследует речной найлок в нижнем течении реки Тавда (август
- 49. Норы Дополнителные источники голоценового материала: местонахождения костных остатков в отложениях нор Cовременные норы таких животных как
- 50. Дополнителные источники голоценового материала: Болота и донные отложения современных озер
- 51. Четвертичные местонахождения Урала и Западной Сибири Четвертичные местонахождения фаун мелких млекопитающих Урала и Западно-Сибирской равнины в
- 52. Позднеплейстоценовые и голоценовые местонахождения мелких млекопитающих и жуков Урала и Западной Сибири с учетом типа фаун
- 53. Видовой состав фаун позднеплейстоценовых фаун мелких млекопитающих изменяется в градиенте север-юг
- 54. The structure of faunas reflect the latitudinal gradient. The three major groups of faunas could be
- 55. Although traditionally named after their relation to glacial processes, the periglacial and glaciomarine zones of the
- 56. Latitudinal zones A1-A5 To account for the latitudinal zonation of the West Siberia and Trans-Urals, we
- 57. The latitudinal biochronological zones are established based on the geographic occurrence of the key lineages of
- 58. An approach to unification of inter-biome and inter-regional correlations of micromammal faunal complexes across latitudinal zones
- 59. АI АIV-AV
- 60. Однако формализация подходов к межрегиональным корреляциям не должны упрощать биологический фон для биохронологических корреляций. При использовании
- 61. Если мы посмотрим на ареалы современных представителей таксонов используемых для биохронологических корреляций, мы можем наблюдать принципиальные
- 62. Хотя реакции на изменения окружающей среды специфичны для каждого таксона, разные виды часто демонстрируют сходные модели
- 63. … и 2) биогеографический фон отдельных регионов накладывает те же ограничения на модели биологического разнообразия. Например,
- 64. Так было в течение последних 2,6 миллионов лет, и фауна этой огромной территории представляет собой арену
- 65. С другой стороны, мы должны помнить, что морфологические различия, сопоставимые с эволюционно значимыми для ключевых таксонов
- 66. Во-первых, географическая изменчивость играет важную роль в формировании морфологического разнообразия: пример существующих видов Dicrostonyx Dicrostonyx species
- 67. Во-вторых, генетический дрейф в небольших изолированных популяциях приводит к морфологическим изменениям, которые противоположны эволюционным тенденциям, известным
- 68. Зная, что материковая часть Евразии в четвертичное время была изменчивой, мы не можем рассматривать эволюционные изменения
- 69. Зная, что материковая часть Евразии в четвертичное время была изменчивой, мы не можем рассматривать эволюционные изменения
- 70. Зная, что материковая часть Евразии в четвертичное время была изменчивой, мы не можем рассматривать эволюционные изменения
- 71. … трансгрессии морей Арктики,
- 72. и трансгрессии Понто-Каспийского на юге…
- 73. мы не можем рассматривать эволюционные изменения как плавные и постепенные преобразования с равными темпами на огромных
- 74. И даже больше: мы не должны забывать о неполноте палеонтологической летописи. В настоящее время нет остатков
- 75. And this is equally important for all index taxa in all biochronologically important lineages of micromammals:
- 76. В настоящее время растет количество доказательств того, что методы молекулярной генетики дают представление о прошлой истории
- 77. Вряд ли, мы когда-либо сможем реализовать молекулярно-генетические методы для создания биостратиграфических единиц для всего четвертичного периода.
- 78. Statistical parsimony networks showing phylogenetic relationships among mtDNA haplotypes from different regions adjacent to Pymva Shor.
- 79. Проведено сравнение видовых реакций на изменение климата в позднечетвертичное время двух важных для арктических экосистем видов
- 80. Рисунок 2– Результаты моделирования экологических ниш копытного лемминга (слева) и узкочерепной полевки (справа). Yellow dots indicate
- 81. Рисунок 1– Динамика генетического разнообразия копытного лемминга и узкочерепной полевки от позднего плейстоцена до современности Этот
- 82. Но что также важно, мы можем использовать молекулярную генетику для палеобиогеографических выводов, используя современные виды в
- 83. Неонтологические архивы
- 84. Pteromys volans L., 1785
- 86. Phylogeography of Carabus sibiricus (unpublished) Modern studied samples Pleistocene localities
- 88. Рис. 3. Филогенетическое дерево, реконструированное методом Байесова анализа на основании 69-ти гаплотипов контрольного региона мтДНК L.
- 90. Заключение В условиях неоднозначности в стратиграфических системах или в сценариях геологической истории палеонтологические материалы становятся особенно
- 91. Заключение При использовании биологических ключевых таксонов для корреляции следует учитывать, что изменчивость является неотъемлемой характеристикой живых
- 92. Заключение В течение четвертичного периода Урал и Западная Сибирь подвергались значительным изменениям фауны, определяемым трансформацией природных
- 93. THE PRESENT IS THE KEY TO THE PAST IS THE KEY TO THE FUTURE НАСТОЯЩЕЕ ОТКРЫВАЕТ
- 94. THE PRESENT IS THE KEY TO THE PAST IS THE KEY TO THE FUTURE НАСТОЯЩЕЕ ОТКРЫВАЕТ
- 95. THE PRESENT IS THE KEY TO THE PAST IS THE KEY TO THE FUTURE НАСТОЯЩЕЕ ОТКРЫВАЕТ
- 97. Скачать презентацию

Лаборатория
филогенетики и биохронологии ИЭРиЖ УрО РАН
Лаборатория Филогенетики и биохронологии образована
Лаборатория
филогенетики и биохронологии ИЭРиЖ УрО РАН
Лаборатория Филогенетики и биохронологии образована

Лаборатория
филогенетики и биохронологии ИЭРиЖ УрО РАН
исследования в рамках четырех основных
Лаборатория
филогенетики и биохронологии ИЭРиЖ УрО РАН
исследования в рамках четырех основных

Лаборатория
филогенетики и биохронологии ИЭРиЖ УрО РАН
исследования в рамках четырех основных
Лаборатория
филогенетики и биохронологии ИЭРиЖ УрО РАН
исследования в рамках четырех основных

Лаборатория
филогенетики и биохронологии ИЭРиЖ УрО РАН
исследования в рамках четырех основных
Лаборатория
филогенетики и биохронологии ИЭРиЖ УрО РАН
исследования в рамках четырех основных

Лаборатория
филогенетики и биохронологии ИЭРиЖ УрО РАН
исследования в рамках четырех основных
Лаборатория
филогенетики и биохронологии ИЭРиЖ УрО РАН
исследования в рамках четырех основных

Геохронология. Возраст Земли 4,6 млрд лет
Геохронология. Возраст Земли 4,6 млрд лет
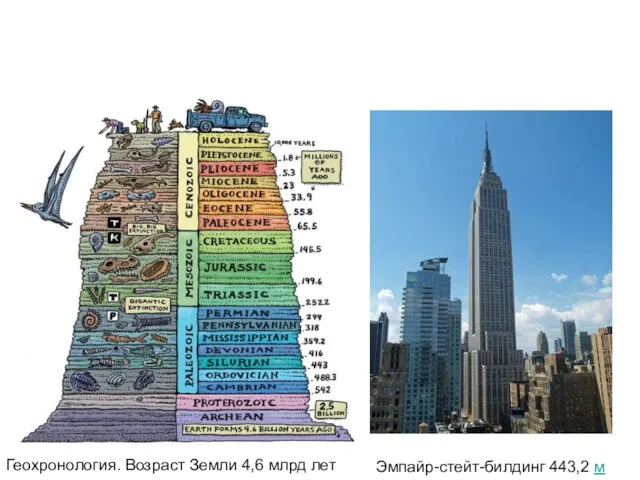
Эмпайр-стейт-билдинг 443,2 м
Геохронология. Возраст Земли 4,6 млрд лет
Эмпайр-стейт-билдинг 443,2 м
Геохронология. Возраст Земли 4,6 млрд лет
Эмпайр-стейт-билдинг 443,2 м
Геохронология. Возраст Земли 4,6 млрд лет
Эмпайр-стейт-билдинг 443,2 м
Геохронология. Возраст Земли 4,6 млрд лет

Зачем нужны архивы?
- реконструировать историю
Зачем необходимо знание истории?
- для
Зачем нужны архивы?
- реконструировать историю
Зачем необходимо знание истории?
- для

Зачем изучать геологическую историю и историю биоты?
"Тот, кто не помнит своего
Зачем изучать геологическую историю и историю биоты?
"Тот, кто не помнит своего

"Тот, кто не помнит своего прошлого, обречен на то, чтобы пережить
"Тот, кто не помнит своего прошлого, обречен на то, чтобы пережить

THE PRESENT IS THE KEY TO THE PAST IS THE KEY
THE PRESENT IS THE KEY TO THE PAST IS THE KEY

THE PRESENT IS THE KEY TO THE PAST IS THE KEY
THE PRESENT IS THE KEY TO THE PAST IS THE KEY

Требования к архивам:
Доступность
Информативность
Интерпретируемость
Требования к архивам:
Доступность
Информативность
Интерпретируемость

Тафоно́мия (от греч. τάφος — «могила, погребение», и νόμος — «закон»)
—раздел палеонтологии и археологии, изучающий закономерности процессов захоронения и образования
Тафоно́мия (от греч. τάφος — «могила, погребение», и νόμος — «закон»)
—раздел палеонтологии и археологии, изучающий закономерности процессов захоронения и образования

Мы будем говорить об изучении геологической истории и истории биоты четвертичного
Мы будем говорить об изучении геологической истории и истории биоты четвертичного

Архивы истории биоты четвертичного периода работают, прежде всего, на
биостратиграфию
Архивы истории биоты четвертичного периода работают, прежде всего, на
биостратиграфию
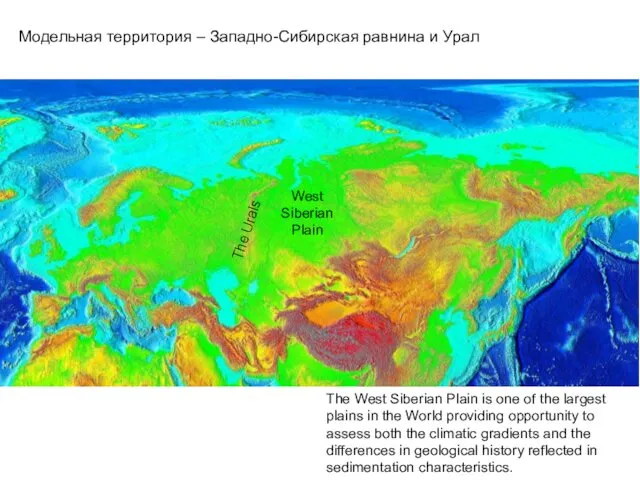
The West Siberian Plain is one of the largest plains in
The West Siberian Plain is one of the largest plains in

В настоящее время во всех научных науках, связанных с изучением четвертичного
В настоящее время во всех научных науках, связанных с изучением четвертичного

Один из удачных примеров трансрегиональных стратиграфических корреляций:
Один из удачных примеров трансрегиональных стратиграфических корреляций:

Вот списоке регионов, включенных в это исследование,
Западно-Сибирская равнина отсутствует
Вот списоке регионов, включенных в это исследование,
Западно-Сибирская равнина отсутствует
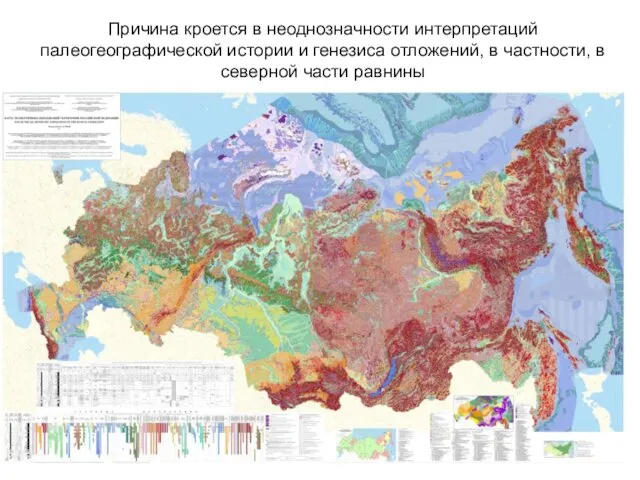
Причина кроется в неоднозначности интерпретаций палеогеографической истории и генезиса отложений,
Причина кроется в неоднозначности интерпретаций палеогеографической истории и генезиса отложений,
В настоящее время Западно-Сибирская равнина характеризуется ярко выраженной широтной зональностью, начиная
В настоящее время Западно-Сибирская равнина характеризуется ярко выраженной широтной зональностью, начиная
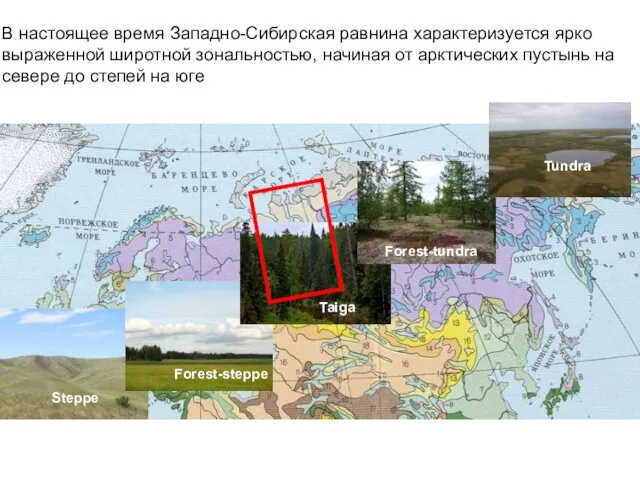
В настоящее время Западно-Сибирская равнина характеризуется ярко выраженной широтной зональностью, начиная
В настоящее время Западно-Сибирская равнина характеризуется ярко выраженной широтной зональностью, начиная
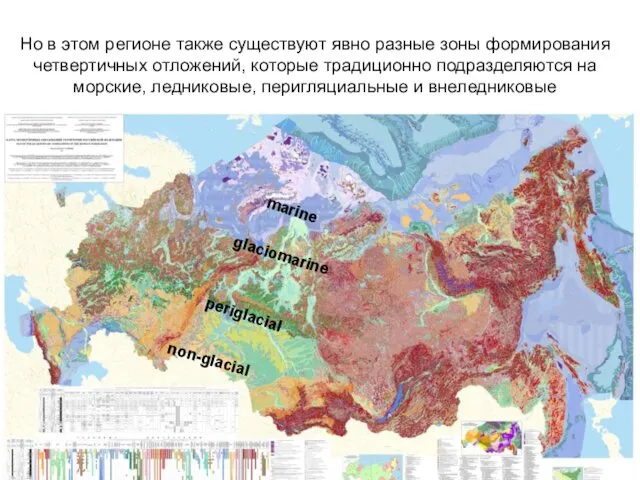
Но в этом регионе также существуют явно разные зоны формирования четвертичных
Но в этом регионе также существуют явно разные зоны формирования четвертичных

Когда корреляция, основанная на небиологических данных, кажется проблематичной, роль биостратиграфии значительно
Когда корреляция, основанная на небиологических данных, кажется проблематичной, роль биостратиграфии значительно

В Северном полушарии полевки представляют группу млекопитающих, которая наиболее широко используется
В Северном полушарии полевки представляют группу млекопитающих, которая наиболее широко используется

Mammuth incessor
Rodent incessor
Для многих территорий основой биостратиграфических построений являются крупные млекопитающие.
Mammuth incessor
Rodent incessor
Для многих территорий основой биостратиграфических построений являются крупные млекопитающие.

Разрезы четвертичных отложений, доступные для исследования, часто дают богатые скопления макроостатков
Разрезы четвертичных отложений, доступные для исследования, часто дают богатые скопления макроостатков

Объекты исследований
В рамках палеонтологического подхода на первом этапе работ объектом исследований
Объекты исследований
В рамках палеонтологического подхода на первом этапе работ объектом исследований

Типы плейстоценовых и голоценовых местонахождений Урала и Западной Сибири
Три аспекта, имеющие
Типы плейстоценовых и голоценовых местонахождений Урала и Западной Сибири
Три аспекта, имеющие

Местонахождения в скальных массивах
Одно из преимуществ пещерных отложений в том, что
Местонахождения в скальных массивах
Одно из преимуществ пещерных отложений в том, что

Cпелеогенные отложения карстовые полости
К этому типу местонахождений относятся пещеры, гроты, скальные
Cпелеогенные отложения карстовые полости
К этому типу местонахождений относятся пещеры, гроты, скальные

Отложения некарстовых гротов
К этому типу относятся неглубокие полости в скальных массивах
Отложения некарстовых гротов
К этому типу относятся неглубокие полости в скальных массивах

Аллювиально- спелеогенные отложения
К этому типу местонахождений относятся пещеры, отложения которых формировались
Аллювиально- спелеогенные отложения
К этому типу местонахождений относятся пещеры, отложения которых формировались

Факторы накопления и источников попадания костей в пещеры
Накопление, распределение и консервация
Факторы накопления и источников попадания костей в пещеры
Накопление, распределение и консервация

Местонахождения равнинной части Зауралья и Западной Сибири
Карьеры и береговые обрывы
Местонахождения равнинной части Зауралья и Западной Сибири
Карьеры и береговые обрывы

Отложения, вскрытые по берегам рек и в карьерах являются основным источником
Отложения, вскрытые по берегам рек и в карьерах являются основным источником

Кроме того, есть несколько дополнительных источников материалов для голоценовых отложений :
торфяники,
Кроме того, есть несколько дополнительных источников материалов для голоценовых отложений :
торфяники,
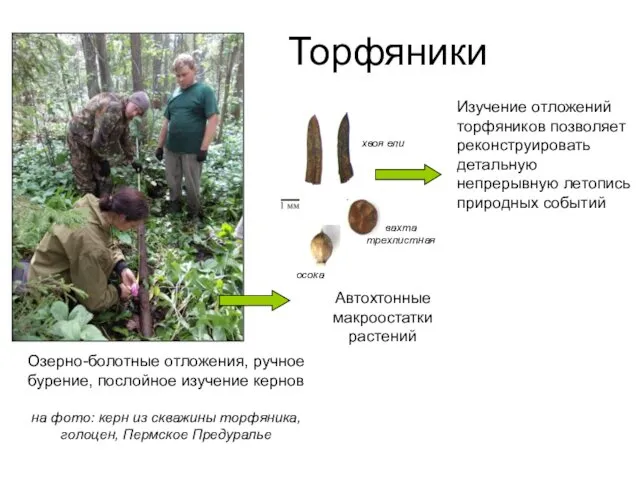
Торфяники
Озерно-болотные отложения, ручное бурение, послойное изучение кернов
на фото: керн из скважины
Торфяники
Озерно-болотные отложения, ручное бурение, послойное изучение кернов на фото: керн из скважины

Одним из источников информации для изучения четвертичных насекомых являются отложения погребенных
Одним из источников информации для изучения четвертичных насекомых являются отложения погребенных

Голоценовые торфяники широко распространены на территории Урала и Западной Сибири. В
Голоценовые торфяники широко распространены на территории Урала и Западной Сибири. В

Археологические памятники
Археологические памятники

Остатки насекомых из археологических памятников активно изучаются в Западной Европе и
Остатки насекомых из археологических памятников активно изучаются в Западной Европе и

Археологические памятники
Культурный слой средневекового поселения, археологический раскоп жилища, зольник около очага
на
Археологические памятники
Культурный слой средневекового поселения, археологический раскоп жилища, зольник около очага на
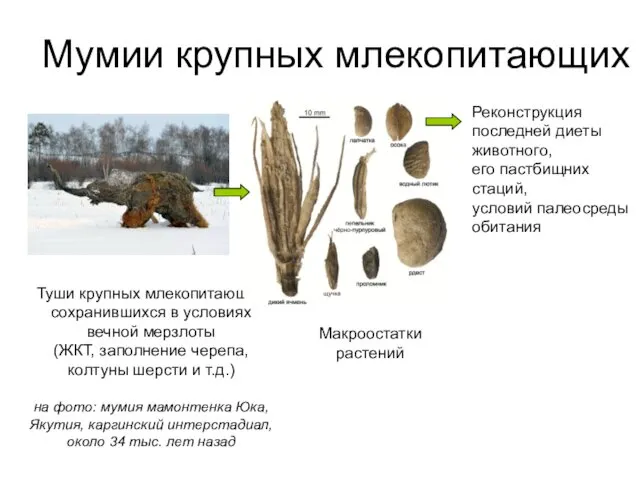
Реконструкция последней диеты животного,
его пастбищних стаций,
условий палеосреды обитания
Туши крупных
Реконструкция последней диеты животного,
его пастбищних стаций,
условий палеосреды обитания
Туши крупных

На фото: казахский энтомолог-четвертичник Ф.Г.Бидашко (Уральск, Казахстан) исследует речной найлок в
На фото: казахский энтомолог-четвертичник Ф.Г.Бидашко (Уральск, Казахстан) исследует речной найлок в

Норы
Дополнителные источники голоценового материала:
местонахождения костных остатков в отложениях нор
Cовременные норы
Норы
Дополнителные источники голоценового материала:
местонахождения костных остатков в отложениях нор
Cовременные норы

Дополнителные источники голоценового материала:
Болота и донные отложения современных озер
Болота и донные отложения современных озер
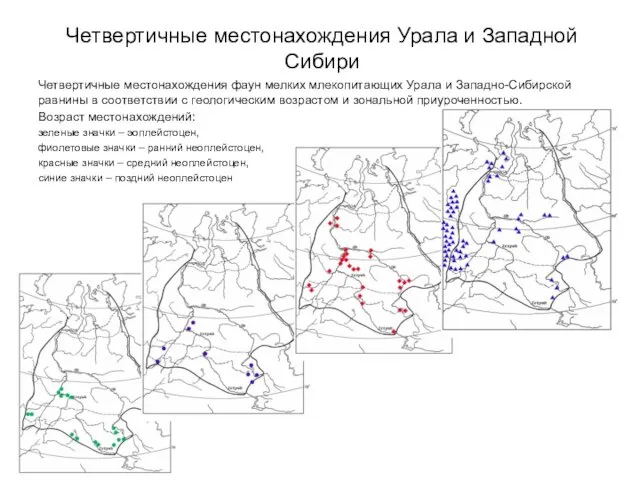
Четвертичные местонахождения Урала и Западной Сибири
Четвертичные местонахождения фаун мелких млекопитающих Урала
Четвертичные местонахождения Урала и Западной Сибири
Четвертичные местонахождения фаун мелких млекопитающих Урала
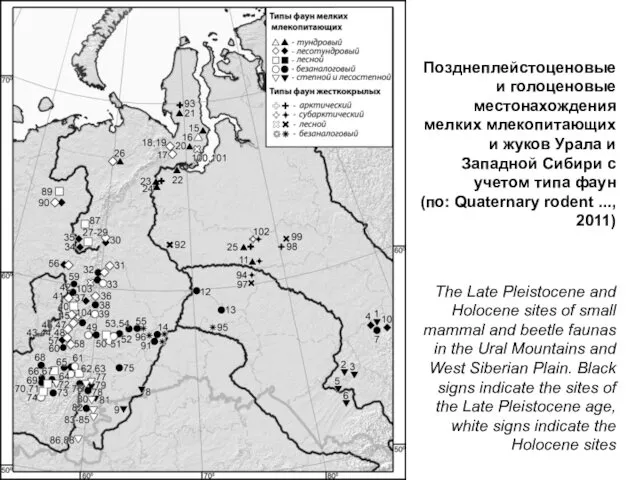
Позднеплейстоценовые и голоценовые местонахождения мелких млекопитающих и жуков Урала и Западной
Позднеплейстоценовые и голоценовые местонахождения мелких млекопитающих и жуков Урала и Западной

Видовой состав фаун позднеплейстоценовых фаун мелких млекопитающих изменяется в градиенте север-юг
Видовой состав фаун позднеплейстоценовых фаун мелких млекопитающих изменяется в градиенте север-юг
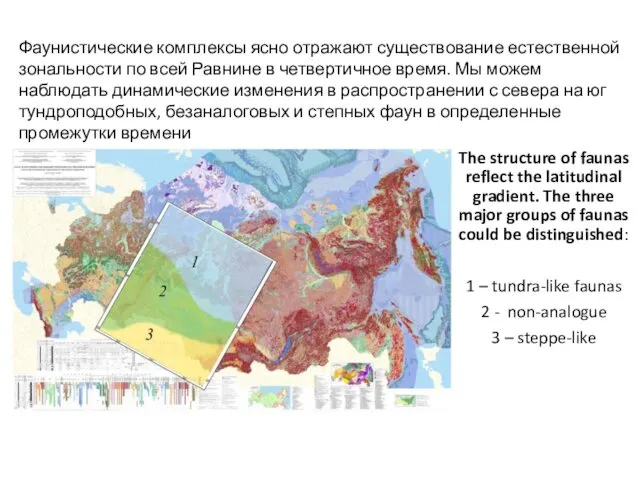
The structure of faunas reflect the latitudinal gradient. The three major
The structure of faunas reflect the latitudinal gradient. The three major
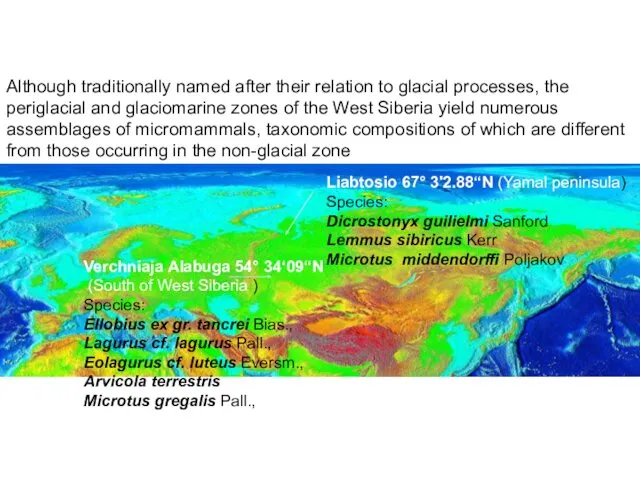
Although traditionally named after their relation to glacial processes, the periglacial
Although traditionally named after their relation to glacial processes, the periglacial
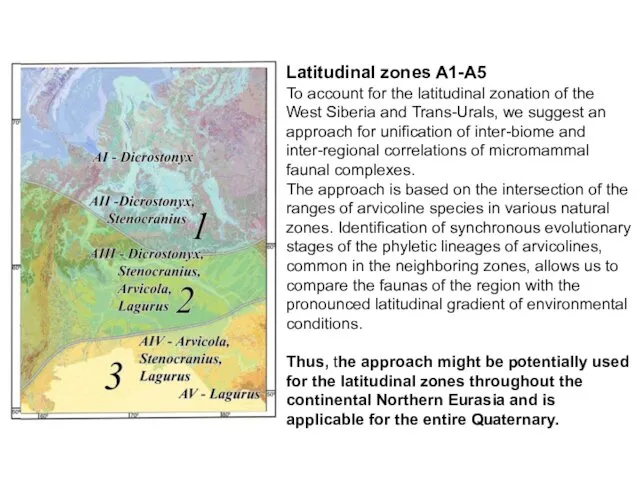
Latitudinal zones A1-A5
To account for the latitudinal zonation of the
Latitudinal zones A1-A5
To account for the latitudinal zonation of the
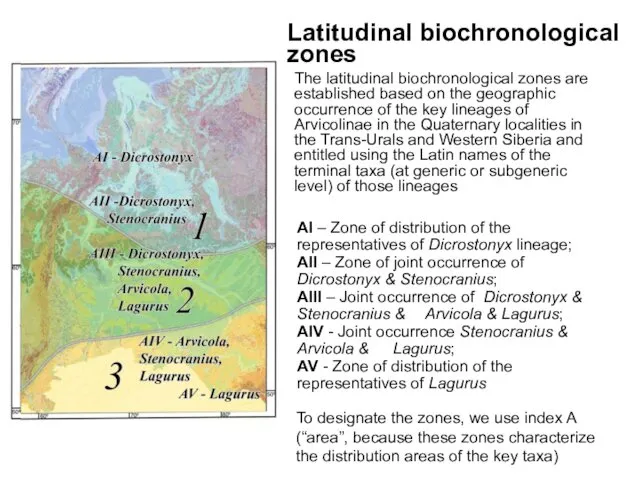
The latitudinal biochronological zones are established based on the geographic occurrence
The latitudinal biochronological zones are established based on the geographic occurrence

An approach to unification of inter-biome and inter-regional correlations of micromammal
An approach to unification of inter-biome and inter-regional correlations of micromammal
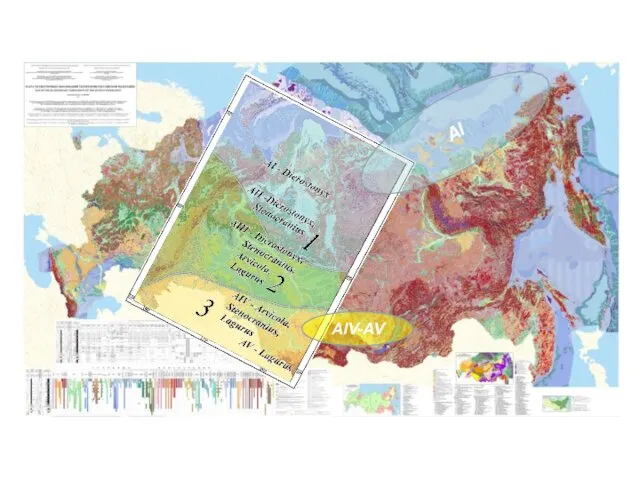
АI
АIV-AV
АI
АIV-AV

Однако формализация подходов к межрегиональным корреляциям не должны упрощать биологический фон
Однако формализация подходов к межрегиональным корреляциям не должны упрощать биологический фон
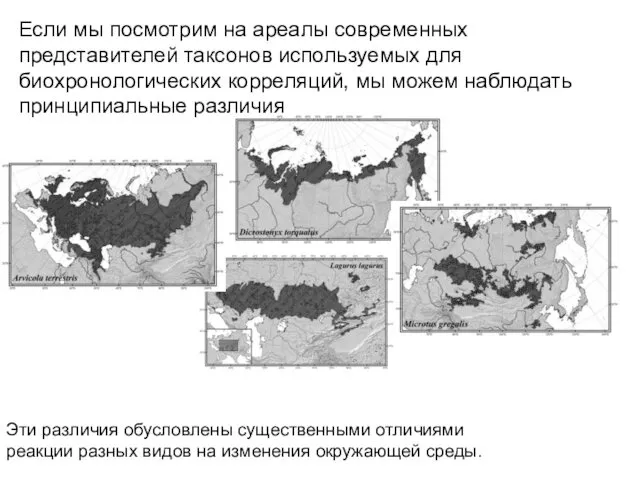
Если мы посмотрим на ареалы современных представителей таксонов используемых для биохронологических
Если мы посмотрим на ареалы современных представителей таксонов используемых для биохронологических

Хотя реакции на изменения окружающей среды специфичны для каждого таксона, разные
Хотя реакции на изменения окружающей среды специфичны для каждого таксона, разные

… и 2) биогеографический фон отдельных регионов накладывает те же ограничения
… и 2) биогеографический фон отдельных регионов накладывает те же ограничения
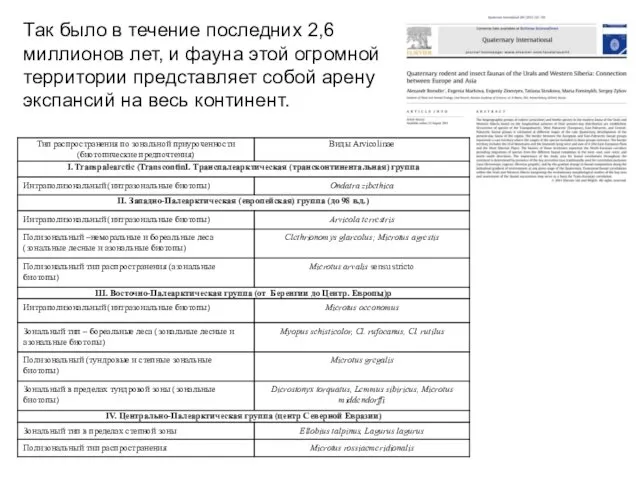
Так было в течение последних 2,6 миллионов лет, и фауна этой
Так было в течение последних 2,6 миллионов лет, и фауна этой

С другой стороны, мы должны помнить, что морфологические различия, сопоставимые с
С другой стороны, мы должны помнить, что морфологические различия, сопоставимые с
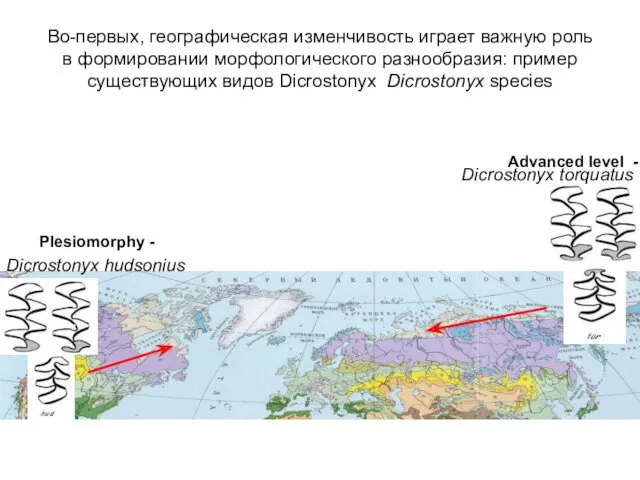
Во-первых, географическая изменчивость играет важную роль в формировании морфологического разнообразия: пример
Во-первых, географическая изменчивость играет важную роль в формировании морфологического разнообразия: пример

Во-вторых, генетический дрейф в небольших изолированных популяциях приводит к морфологическим изменениям,
Во-вторых, генетический дрейф в небольших изолированных популяциях приводит к морфологическим изменениям,

Зная, что материковая часть Евразии в четвертичное время была изменчивой, мы
Зная, что материковая часть Евразии в четвертичное время была изменчивой, мы

Зная, что материковая часть Евразии в четвертичное время была изменчивой, мы
Зная, что материковая часть Евразии в четвертичное время была изменчивой, мы

Зная, что материковая часть Евразии в четвертичное время была изменчивой, мы
Зная, что материковая часть Евразии в четвертичное время была изменчивой, мы

… трансгрессии морей Арктики,
… трансгрессии морей Арктики,
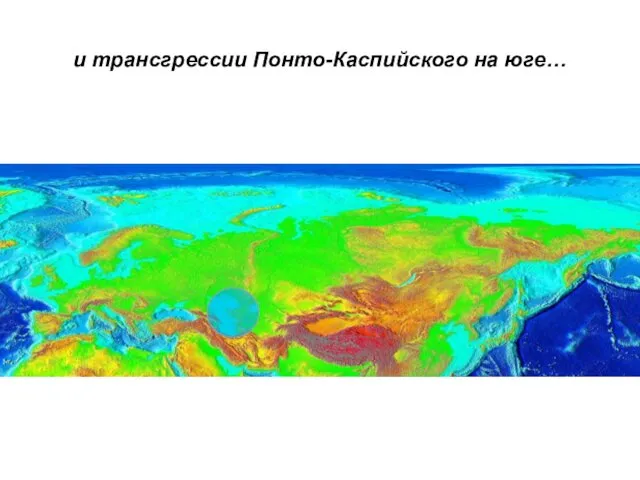
и трансгрессии Понто-Каспийского на юге…
и трансгрессии Понто-Каспийского на юге…
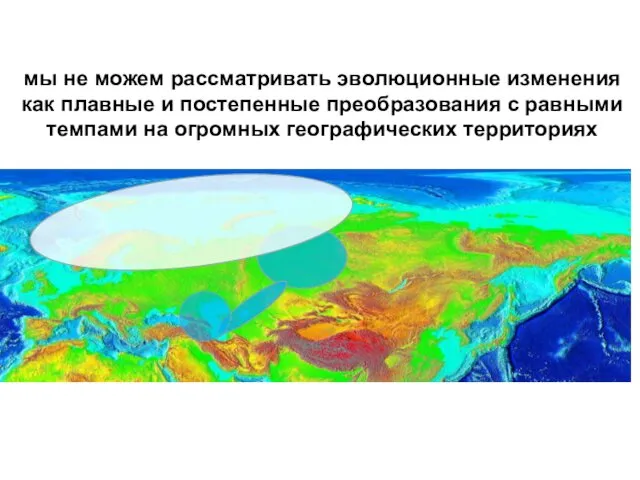
мы не можем рассматривать эволюционные изменения как плавные и постепенные преобразования
мы не можем рассматривать эволюционные изменения как плавные и постепенные преобразования

И даже больше: мы не должны забывать о неполноте палеонтологической летописи.
И даже больше: мы не должны забывать о неполноте палеонтологической летописи.

And this is equally important for all index taxa in all
And this is equally important for all index taxa in all

В настоящее время растет количество доказательств того, что методы молекулярной генетики
В настоящее время растет количество доказательств того, что методы молекулярной генетики

Вряд ли, мы когда-либо сможем реализовать молекулярно-генетические методы для создания биостратиграфических
Вряд ли, мы когда-либо сможем реализовать молекулярно-генетические методы для создания биостратиграфических
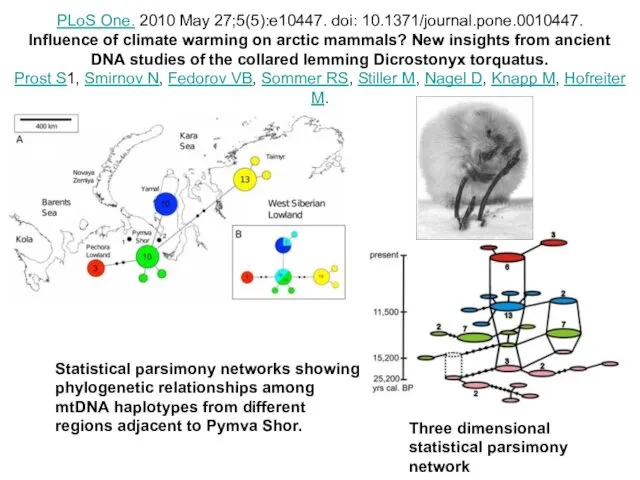
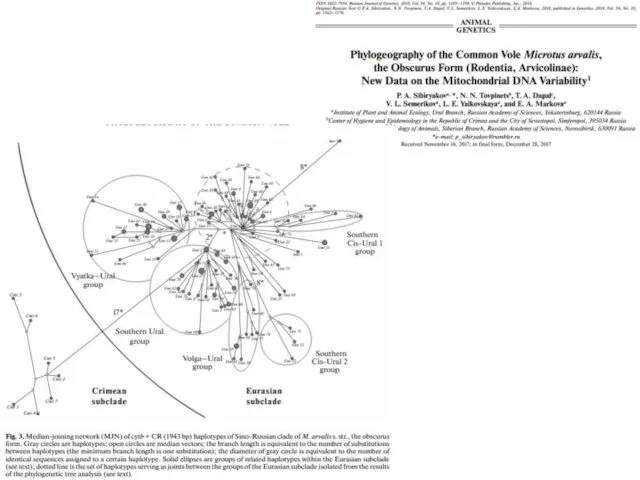
Statistical parsimony networks showing phylogenetic relationships among mtDNA haplotypes from different
Statistical parsimony networks showing phylogenetic relationships among mtDNA haplotypes from different

Проведено сравнение видовых реакций на изменение климата в позднечетвертичное время двух
Проведено сравнение видовых реакций на изменение климата в позднечетвертичное время двух

Рисунок 2– Результаты моделирования экологических ниш копытного лемминга (слева) и узкочерепной
Рисунок 2– Результаты моделирования экологических ниш копытного лемминга (слева) и узкочерепной
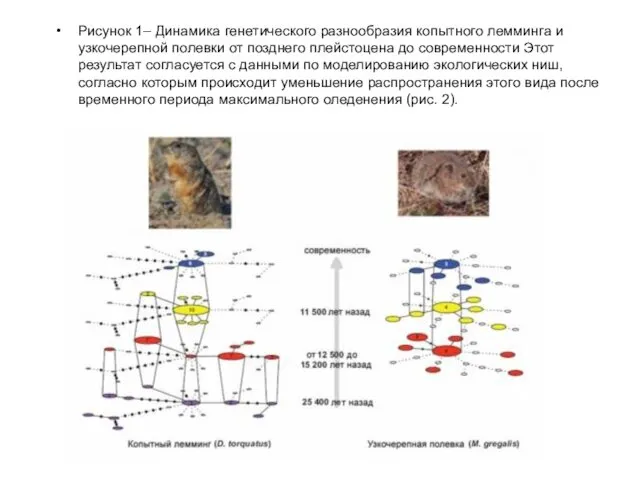
Рисунок 1– Динамика генетического разнообразия копытного лемминга и узкочерепной полевки от
Рисунок 1– Динамика генетического разнообразия копытного лемминга и узкочерепной полевки от

Но что также важно, мы можем использовать молекулярную генетику для палеобиогеографических
Но что также важно, мы можем использовать молекулярную генетику для палеобиогеографических

Неонтологические архивы
Неонтологические архивы

Pteromys volans L., 1785
Pteromys volans L., 1785

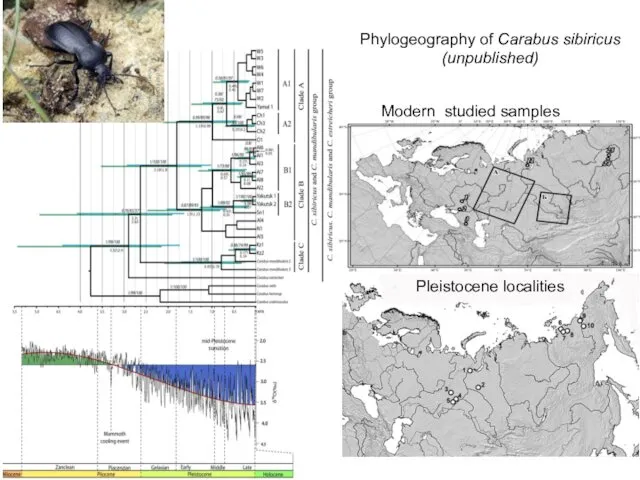
Phylogeography of Carabus sibiricus
(unpublished)
Modern studied samples
Pleistocene localities
Phylogeography of Carabus sibiricus
(unpublished)
Modern studied samples
Pleistocene localities
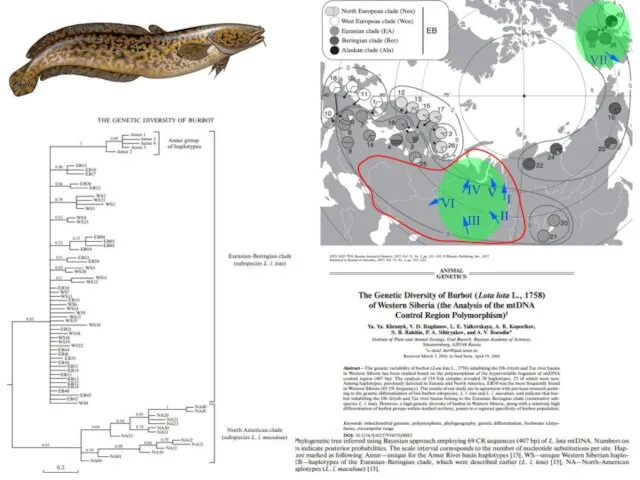
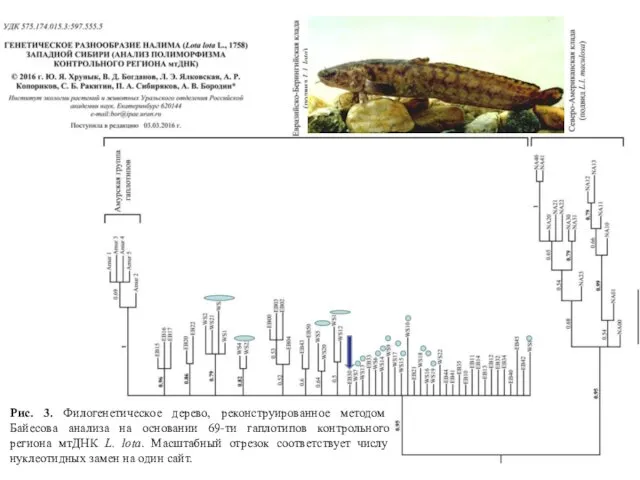
Рис. 3. Филогенетическое дерево, реконструированное методом Байесова анализа на основании 69-ти
Рис. 3. Филогенетическое дерево, реконструированное методом Байесова анализа на основании 69-ти

Заключение
В условиях неоднозначности в стратиграфических системах или в сценариях геологической истории
Заключение
В условиях неоднозначности в стратиграфических системах или в сценариях геологической истории

Заключение
При использовании биологических ключевых таксонов для корреляции следует учитывать, что изменчивость
Заключение
При использовании биологических ключевых таксонов для корреляции следует учитывать, что изменчивость

Заключение
В течение четвертичного периода Урал и Западная Сибирь подвергались значительным изменениям
Заключение
В течение четвертичного периода Урал и Западная Сибирь подвергались значительным изменениям

THE PRESENT IS THE KEY TO THE PAST IS THE KEY
THE PRESENT IS THE KEY TO THE PAST IS THE KEY

THE PRESENT IS THE KEY TO THE PAST IS THE KEY
THE PRESENT IS THE KEY TO THE PAST IS THE KEY

THE PRESENT IS THE KEY TO THE PAST IS THE KEY
THE PRESENT IS THE KEY TO THE PAST IS THE KEY
 Компания Сибирское здоровье. Продукт для профилактики хронических дегенеративных поражений суставов
Компания Сибирское здоровье. Продукт для профилактики хронических дегенеративных поражений суставов Презентация С единым днем защиты интернета
Презентация С единым днем защиты интернета Презентация к конспекту образовательной деятельности.Тема: Мы веселые зайчата
Презентация к конспекту образовательной деятельности.Тема: Мы веселые зайчата Презентация кружковой работы по бисероплетению
Презентация кружковой работы по бисероплетению фото кабинета этнокультуры Диск
фото кабинета этнокультуры Диск Алгоритм работы вожатого в организационный период
Алгоритм работы вожатого в организационный период Ежемесячная выплата в связи с рождением второго ребенка
Ежемесячная выплата в связи с рождением второго ребенка Кто такие насекомые
Кто такие насекомые Изменения в деятельности учителя при переходе на ФГОС ООО
Изменения в деятельности учителя при переходе на ФГОС ООО История становления специальной педагогики в России
История становления специальной педагогики в России Консервовані зелені корми: сіно, трав’яне борошно
Консервовані зелені корми: сіно, трав’яне борошно РАЗВИТИЕ МЕЛКОЙ МОТОРИКИ РУК В ИГРАХ-ЗАНЯТИЯХ С ДЕТЬМИ С ОГРАНИЧЕННЫМИ ВОЗМОЖНОСТЯМИ ЗДОРОВЬЯ
РАЗВИТИЕ МЕЛКОЙ МОТОРИКИ РУК В ИГРАХ-ЗАНЯТИЯХ С ДЕТЬМИ С ОГРАНИЧЕННЫМИ ВОЗМОЖНОСТЯМИ ЗДОРОВЬЯ Васильев Михаил 9а
Васильев Михаил 9а Воспитание у дошкольников основ экологической культуры
Воспитание у дошкольников основ экологической культуры Фигуры на клетчатой бумаге
Фигуры на клетчатой бумаге Желудочно-кишечные кровотечения: современное состояние и перспективы проблемы
Желудочно-кишечные кровотечения: современное состояние и перспективы проблемы Россия в первой половине XVII века
Россия в первой половине XVII века Анемии у детей и подростков: другие виды
Анемии у детей и подростков: другие виды Решение конфликта.
Решение конфликта. Музыкальный образ и мастерство исполнителя
Музыкальный образ и мастерство исполнителя Россия в первой половине XVIII века
Россия в первой половине XVIII века Предметно-развивающая среда как средство построения здоровьесберегающего пространства в ДОУ
Предметно-развивающая среда как средство построения здоровьесберегающего пространства в ДОУ Примерная структура планирования воспитательно-образовательной работы на 1 день
Примерная структура планирования воспитательно-образовательной работы на 1 день Теория государства и права, как наука и учебная дисциплина
Теория государства и права, как наука и учебная дисциплина Презентация классного часа на тему Люблю березку русскую
Презентация классного часа на тему Люблю березку русскую Информационно-аналитический журнал Агровыбор Чувашии
Информационно-аналитический журнал Агровыбор Чувашии Внеклассное занятие Семь чудес Санкт-Петербурга
Внеклассное занятие Семь чудес Санкт-Петербурга Использование современных технологий и методик в педагогической деятельности воспитателя
Использование современных технологий и методик в педагогической деятельности воспитателя