- Палиндромы и рестриктазы

Содержание
- 2. Определение палиндрома. Палиндром (palindrome): последовательность ДНК, состоящая из смежных инвертированных повторов, одинаково считывающихся и в левом
- 3. Крестообразная форма палиндрома. В природных двунитевых ДНК часто встречаются палиндромные последовательности, обладающие осью симметрии второго порядка
- 4. Линейная форма палиндрома. В линейной ДНК крестообразная структура палиндромных областей является термодинамически невыгодной по сравнению с
- 5. Открытие крестообразных структур в природных кольцевых ДНК. Образование крестообразных структур в природных кольцевых ДНК было открыто
- 6. Эндонуклеазы как часть системы рестрикции-модификации. В ходе эволюции бактерии развили способность синтезировать так называемые рестрицирующие ферменты
- 7. Способ защиты бактериальных клеток от чужеродных ДНК. С целью защиты бактерии в ходе эволюции развили также
- 8. Эндонуклеазы, осуществляющие рестрикцию молекул ДНК. Рестриктазы впервые были выделены из Е. coli в 1968 г. Оказалось,
- 9. Палиндромы как сайты рестрикции ДНК. Действие системы рестрикции-модификации «рационализуется» так называемыми палиндромными (распознающими последовательностями) азотистых оснований,
- 10. Характерные особенности рестриктаз. Рестриктазы — это абсолютно необходимый инструмент в генной инженерии для вырезания интересующих фрагментов
- 11. Результаты рестрикции. Так, известная рестриктаза – фермент из системы рестрикция—модификация Е. coli вместо того, чтобы плавить
- 12. Достижение эффективности смыкания ДНК-фрагментов. «Липкие» концы в генно-инженерных опытах полезны по той причине, что они могут
- 13. Электрофорез как метод фракционирования рестрикцированной ДНК. Сайты распознания и сайты плавления в случае других рестриктаз имеют
- 14. Механизм проведения электрофореза. Фрагменты в электрическом поле мигрируют в ходе электрофореза при частоте, зависимой от их
- 15. Построение рестрикционных карт. После электрофореза фрагменты из агарозы выделяют с помощью разных методов. На основании результатов
- 16. Методы сенквенирования. Выделенные после электрофореза из агарозных гелей фрагменты ДНК (рестрикты) можно предварительно подвергнуть секвенированию, т.
- 17. Использование комплементарной ДНК в процессах рестрикции у высших организмов. Поскольку у высших организмов в ходе транскрипции
- 18. Список литературы. А.С. Коничев, Г.А. Севастьянова. Молекулярная биология, Москва, издательский центр «Академия», 2012 г., 395 стр.
- 20. Скачать презентацию

Определение палиндрома.
Палиндром (palindrome): последовательность ДНК, состоящая из смежных инвертированных повторов, одинаково
Определение палиндрома.
Палиндром (palindrome): последовательность ДНК, состоящая из смежных инвертированных повторов, одинаково
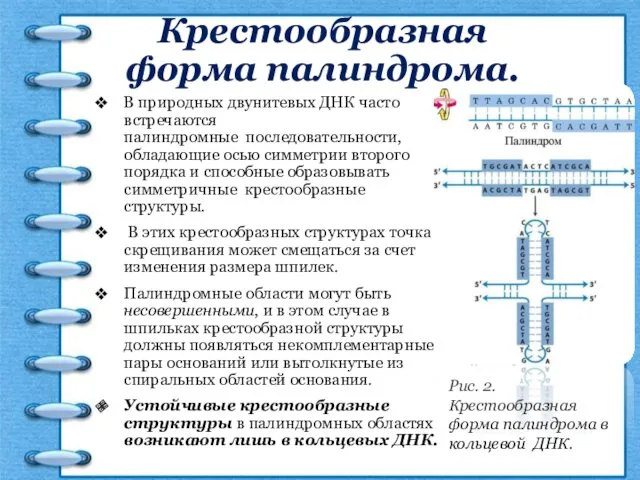
Крестообразная форма палиндрома.
В природных двунитевых ДНК часто встречаются палиндромные последовательности, обладающие
Крестообразная форма палиндрома.
В природных двунитевых ДНК часто встречаются палиндромные последовательности, обладающие

Линейная форма палиндрома.
В линейной ДНК крестообразная структура палиндромных областей является термодинамически
Линейная форма палиндрома.
В линейной ДНК крестообразная структура палиндромных областей является термодинамически
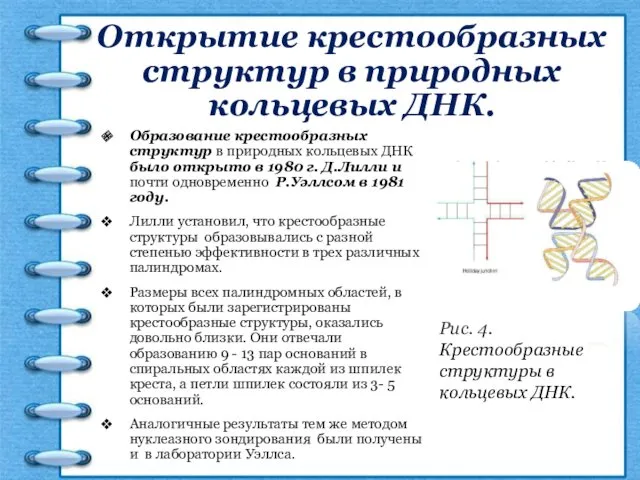
Открытие крестообразных структур в природных кольцевых ДНК.
Образование крестообразных структур в природных
Открытие крестообразных структур в природных кольцевых ДНК.
Образование крестообразных структур в природных

Эндонуклеазы как часть системы рестрикции-модификации.
В ходе эволюции бактерии развили способность синтезировать
Эндонуклеазы как часть системы рестрикции-модификации.
В ходе эволюции бактерии развили способность синтезировать

Способ защиты
бактериальных клеток
от чужеродных ДНК.
С целью защиты бактерии в
Способ защиты
бактериальных клеток
от чужеродных ДНК.
С целью защиты бактерии в

Эндонуклеазы, осуществляющие
рестрикцию молекул ДНК.
Рестриктазы впервые были выделены из Е. coli
Эндонуклеазы, осуществляющие
рестрикцию молекул ДНК.
Рестриктазы впервые были выделены из Е. coli
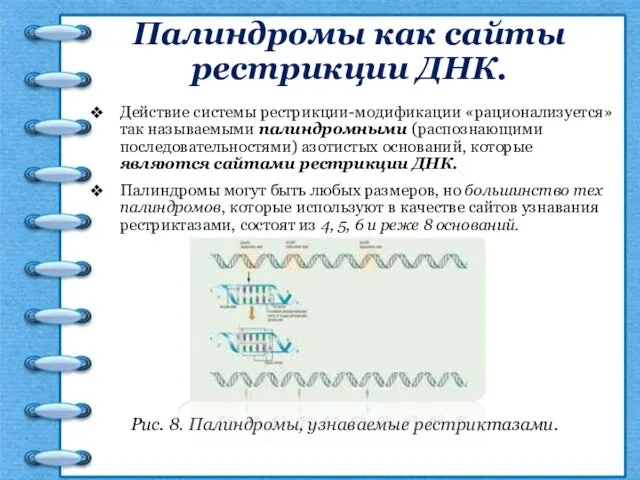
Палиндромы как сайты рестрикции ДНК.
Действие системы рестрикции-модификации «рационализуется» так называемыми палиндромными
Палиндромы как сайты рестрикции ДНК.
Действие системы рестрикции-модификации «рационализуется» так называемыми палиндромными
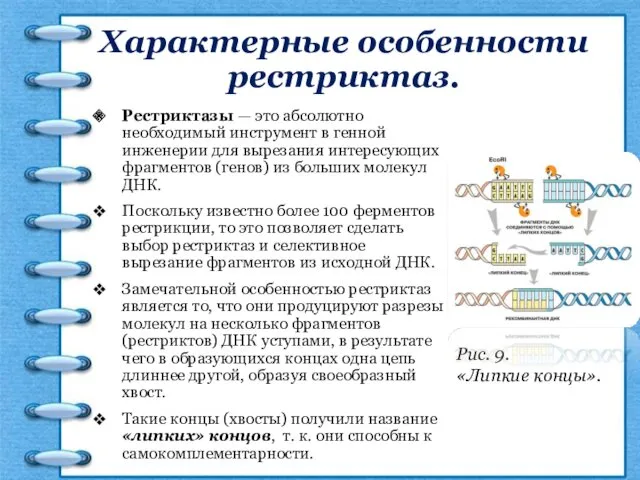
Характерные особенности рестриктаз.
Рестриктазы — это абсолютно необходимый инструмент в генной инженерии
Характерные особенности рестриктаз.
Рестриктазы — это абсолютно необходимый инструмент в генной инженерии
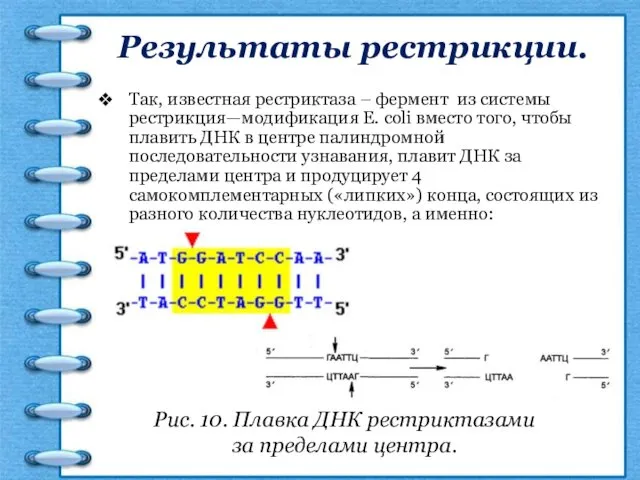
Результаты рестрикции.
Так, известная рестриктаза – фермент из системы рестрикция—модификация Е. coli
Результаты рестрикции.
Так, известная рестриктаза – фермент из системы рестрикция—модификация Е. coli

Достижение эффективности
смыкания ДНК-фрагментов.
«Липкие» концы в генно-инженерных опытах полезны по той
Достижение эффективности
смыкания ДНК-фрагментов.
«Липкие» концы в генно-инженерных опытах полезны по той

Электрофорез как метод фракционирования рестрикцированной ДНК.
Сайты распознания и сайты плавления в
Электрофорез как метод фракционирования рестрикцированной ДНК.
Сайты распознания и сайты плавления в
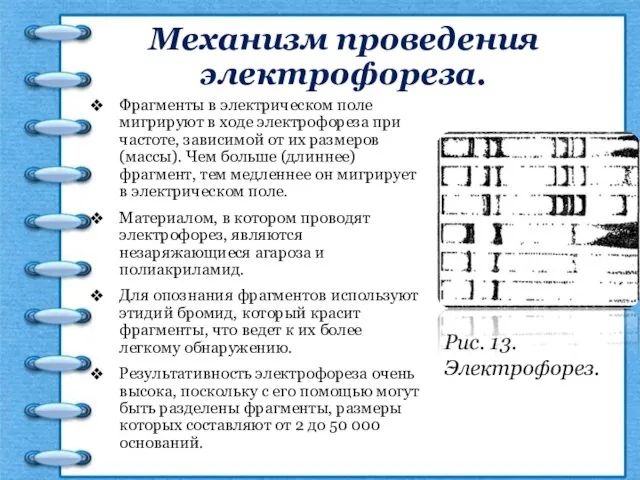
Механизм проведения электрофореза.
Фрагменты в электрическом поле мигрируют в ходе электрофореза при
Механизм проведения электрофореза.
Фрагменты в электрическом поле мигрируют в ходе электрофореза при

Построение
рестрикционных карт.
После электрофореза фрагменты из агарозы выделяют с помощью разных
Построение
рестрикционных карт.
После электрофореза фрагменты из агарозы выделяют с помощью разных

Методы сенквенирования.
Выделенные после электрофореза из агарозных гелей фрагменты ДНК (рестрикты) можно
Методы сенквенирования.
Выделенные после электрофореза из агарозных гелей фрагменты ДНК (рестрикты) можно

Использование комплементарной ДНК в процессах рестрикции у высших организмов.
Поскольку у высших
Использование комплементарной ДНК в процессах рестрикции у высших организмов.
Поскольку у высших

Список литературы.
А.С. Коничев, Г.А. Севастьянова. Молекулярная биология, Москва, издательский центр «Академия»,
Список литературы.
А.С. Коничев, Г.А. Севастьянова. Молекулярная биология, Москва, издательский центр «Академия»,
 300 лет Ломоносову
300 лет Ломоносову Дети войны
Дети войны Родительское собрание Добрые трации семьи
Родительское собрание Добрые трации семьи Етапи розробки презентації. Елементи дизайну презентації
Етапи розробки презентації. Елементи дизайну презентації Отчет о работе т/о Хореография за учебный год
Отчет о работе т/о Хореография за учебный год Усиление деревянных конструкций (2)
Усиление деревянных конструкций (2) Мишка и дни недели
Мишка и дни недели Классический стиль
Классический стиль Дизайн штор.
Дизайн штор. Музейная антресоль
Музейная антресоль Буровые растворы
Буровые растворы Технологическая карта изготовления Кактуса
Технологическая карта изготовления Кактуса Безопасность студентов в стенах образовательного учреждения
Безопасность студентов в стенах образовательного учреждения Природная зональность
Природная зональность Национальная японская кухня
Национальная японская кухня Ақмола облысы, Целиноград ауданы жағдайында қиярды көшетпен өсіру технологиясы
Ақмола облысы, Целиноград ауданы жағдайында қиярды көшетпен өсіру технологиясы Печиво
Печиво Рейтинг подразделений по подаче инициатив ДЦ, ЖДЦ
Рейтинг подразделений по подаче инициатив ДЦ, ЖДЦ Alternative energy
Alternative energy Объекты управления и их основные свойства. Автоматизированное управление ХТС Лекция 4
Объекты управления и их основные свойства. Автоматизированное управление ХТС Лекция 4 Основные компоненты материнской платы
Основные компоненты материнской платы Питание в походе
Питание в походе Мотивационные теории
Мотивационные теории Для родителей об адаптации первоклассников
Для родителей об адаптации первоклассников Введение прикорма
Введение прикорма Мышцы. Мышечная система
Мышцы. Мышечная система Учебно - методическое пособие:Пресноводные рыбы. Игры и упражнения.
Учебно - методическое пособие:Пресноводные рыбы. Игры и упражнения. Производственный менеджмент часть 1 (1)
Производственный менеджмент часть 1 (1)