- Перестройки генома в онтогенезе. 10 класс

Содержание
- 2. План Перестройки генома у прокариот Перестройки генома у эукариот Иммуноглобулиновые гены у млекопитающих Незапрограммированные перестройки генома
- 3. Строение прокариот
- 4. Перестройка генома у прокариот Геном прокариот проще, т.к. у них меньше генов У прокариот нет некоторых
- 5. У бактерий в геноме присутствуют гены, ненужные в нормальных условиях, но необходимые для образование спор. Эти
- 6. Например, ген фермента нитрогеназы у азотфиксирующих цианобактерий. Этот фермент восстанавливает атмосферный азот до аммиака. Но кислород
- 7. Восстановление рамки считывания в геноме цианобактерии с помощью вырезания участков ДНК
- 8. Перестройка генома у эукариот Геномные перестройки, значительно более сложные, чем у прокариот: дифференцированные клетки используют только
- 9. Перестройка генома у эукариот у некоторых червей и членистоногих генетический материал полностью сохраняется только в генеративных
- 10. У человеческой аскариды зигота и клетки –предшественники половых, имеют всего две крупные хромосомы. В ходе дробления
- 11. Удаление части ДНК во время дробления зародыша лошадиной аскариды: А — стадия двух бластомеров; Б —
- 12. Перестройки у инфузорий туфелек У одноклеточных двухядерных эукариот-инфузорий на соматические и генеративные дифференцируются не клетки, а
- 13. Микронуклеус содержит полный геном, с его генов почти не считываются м-РНК и, следовательно, его гены не
- 14. Удаление ДНК иногда происходит и у млекопитающих, хотя и в меньших масштабах. Например, при дифференцировке кроветворных
- 16. Перестройки генома могут быть использованы клетками не только для необратимого выключения части генома, но и, наоборот,
- 17. Наиболее сложные изменения генома происходят у млекопитающих при формировании механизмов иммунитета (образование антител).
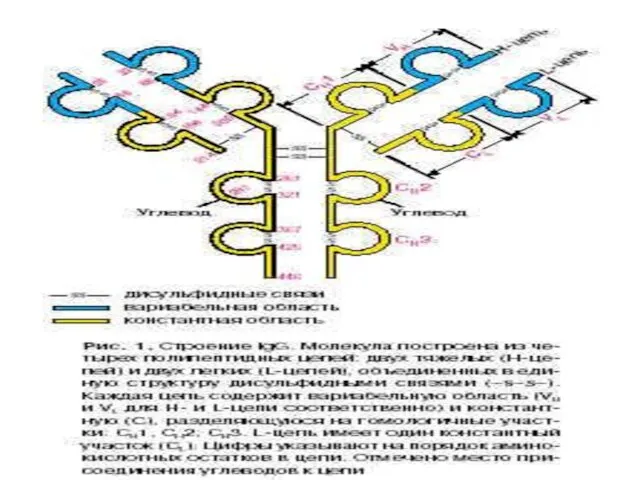
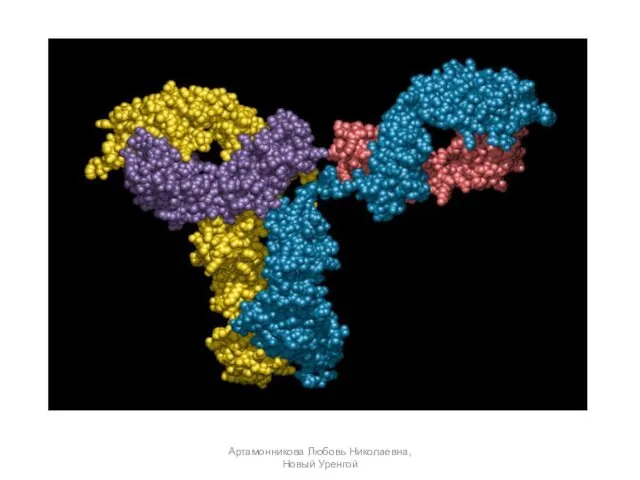
- 18. Молекула каждого иммуноглобулина состоит из двух легких и двух тяжелых полипептидных цепей. Каждая из этих четырех
- 19. Как можно закодировать такое большое разнообразие? Гены иммуноглобулинов имеют прерывистую структуру ,отдельные участки лежат на некотором
- 20. Строение молекулы иммуноглобулина: / — неизменяемая часть молекулы; 2 — вариабельная часть молекулы; 3 — тяжелая
- 21. Вариабельные участки иммуноглобулина образуют активный центр антитела, который взаимодействует с антигеном. Поэтому пространственная структура активного центра
- 23. В период эмбрионального развития организма при размножении клеток—предшественниц лимфоцитов происходит выборочная репликация этих генов, при которой
- 25. Мутационный процесс создает дополнительный источник разнообразия структуры антител. Лимфоцит с уже готовыми собранными генами может синтезировать
- 26. Артамонникова Любовь Николаевна,Новый Уренгой
- 28. Незапрограммированные перестройки генома. К таким перестройкам относят перемещения мобильных генетических элементов (МГЭ). Б. Мак Клинток -лауреат
- 29. Мобильные генетические элементы –МГЭ -последовательности ДНК, которые могут перемещаться внутри генома. Существует несколько классов мобильных элементов
- 31. Скачать презентацию

План
Перестройки генома у прокариот
Перестройки генома у эукариот
Иммуноглобулиновые гены у млекопитающих
Незапрограммированные перестройки
План
Перестройки генома у прокариот
Перестройки генома у эукариот
Иммуноглобулиновые гены у млекопитающих
Незапрограммированные перестройки
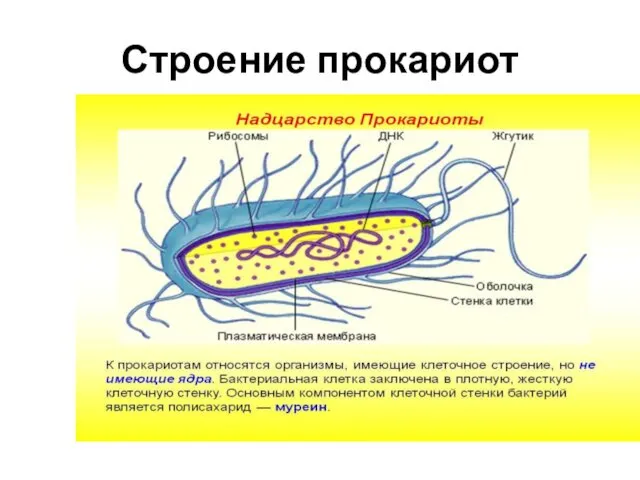
Строение прокариот
Строение прокариот

Перестройка генома у прокариот
Геном прокариот проще, т.к. у них меньше генов
У
Перестройка генома у прокариот
Геном прокариот проще, т.к. у них меньше генов
У

У бактерий в геноме присутствуют гены, ненужные в нормальных условиях,
У бактерий в геноме присутствуют гены, ненужные в нормальных условиях,

Например, ген фермента нитрогеназы у азотфиксирующих цианобактерий.
Этот фермент восстанавливает атмосферный
Например, ген фермента нитрогеназы у азотфиксирующих цианобактерий. Этот фермент восстанавливает атмосферный
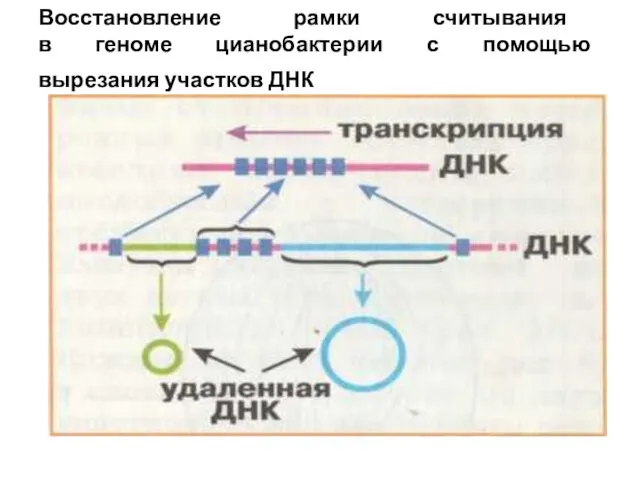
Восстановление рамки считывания
в геноме цианобактерии с помощью
вырезания участков ДНК
Восстановление рамки считывания
в геноме цианобактерии с помощью
вырезания участков ДНК

Перестройка генома у эукариот
Геномные перестройки, значительно более сложные, чем
Перестройка генома у эукариот
Геномные перестройки, значительно более сложные, чем

Перестройка генома у эукариот
у некоторых червей и членистоногих генетический материал полностью
Перестройка генома у эукариот
у некоторых червей и членистоногих генетический материал полностью

У человеческой аскариды зигота и клетки –предшественники половых, имеют всего две
У человеческой аскариды зигота и клетки –предшественники половых, имеют всего две
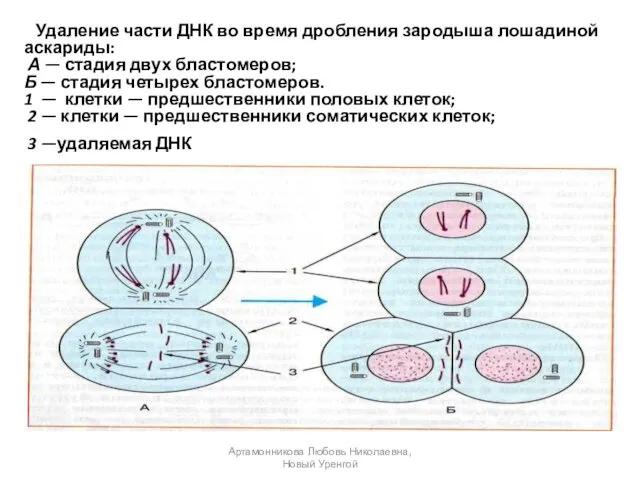
Удаление части ДНК во время дробления зародыша лошадиной аскариды:
А
Удаление части ДНК во время дробления зародыша лошадиной аскариды: А
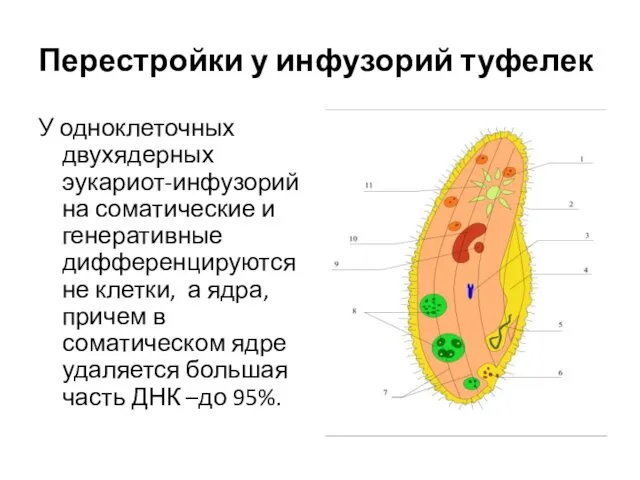
Перестройки у инфузорий туфелек
У одноклеточных двухядерных эукариот-инфузорий на соматические и генеративные
Перестройки у инфузорий туфелек
У одноклеточных двухядерных эукариот-инфузорий на соматические и генеративные

Микронуклеус содержит полный геном, с его генов почти не считываются
Микронуклеус содержит полный геном, с его генов почти не считываются

Удаление ДНК иногда происходит и у млекопитающих, хотя и в
Удаление ДНК иногда происходит и у млекопитающих, хотя и в
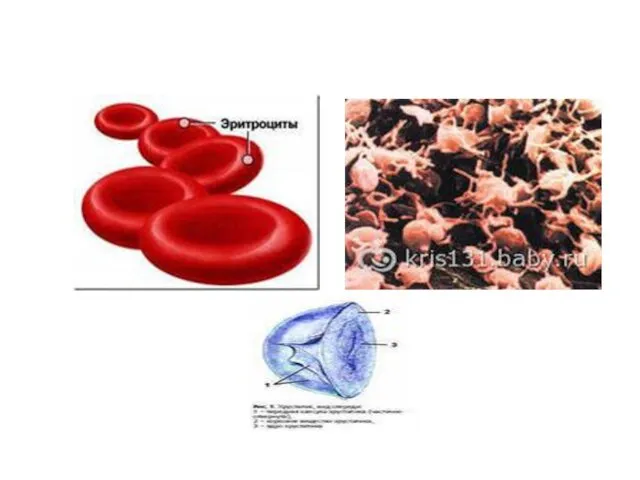

Перестройки генома могут быть использованы клетками не только для необратимого
Перестройки генома могут быть использованы клетками не только для необратимого

Наиболее сложные изменения генома происходят у млекопитающих при формировании механизмов иммунитета
Наиболее сложные изменения генома происходят у млекопитающих при формировании механизмов иммунитета

Молекула каждого иммуноглобулина состоит из двух легких и двух тяжелых
Молекула каждого иммуноглобулина состоит из двух легких и двух тяжелых

Как можно закодировать такое большое разнообразие?
Гены иммуноглобулинов имеют прерывистую структуру
Как можно закодировать такое большое разнообразие? Гены иммуноглобулинов имеют прерывистую структуру
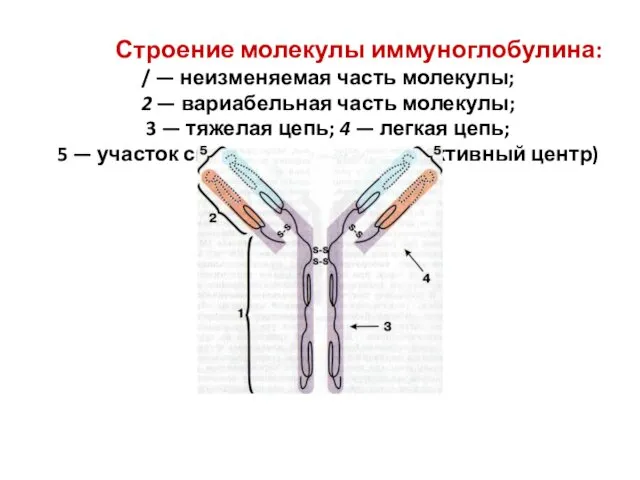
Строение молекулы иммуноглобулина:
/ — неизменяемая часть молекулы;
2 — вариабельная
Строение молекулы иммуноглобулина: / — неизменяемая часть молекулы; 2 — вариабельная

Вариабельные участки иммуноглобулина образуют активный центр антитела,
который взаимодействует с антигеном.
Поэтому пространственная
Вариабельные участки иммуноглобулина образуют активный центр антитела, который взаимодействует с антигеном. Поэтому пространственная
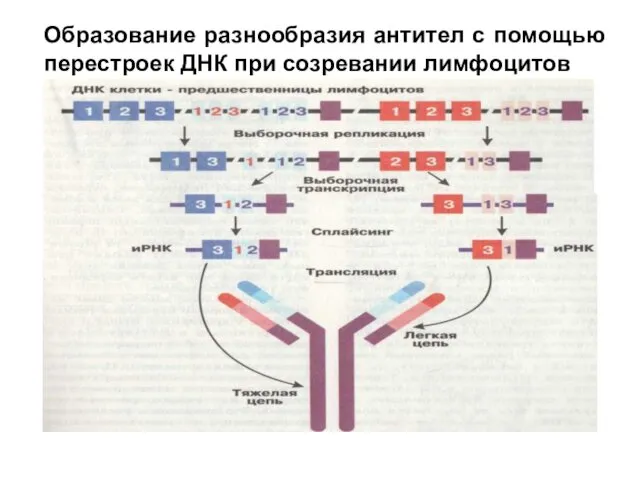

В период эмбрионального развития организма при размножении клеток—предшественниц лимфоцитов происходит
В период эмбрионального развития организма при размножении клеток—предшественниц лимфоцитов происходит

Мутационный процесс создает дополнительный источник разнообразия структуры антител.
Лимфоцит с
Мутационный процесс создает дополнительный источник разнообразия структуры антител. Лимфоцит с
Артамонникова Любовь Николаевна,Новый Уренгой
Артамонникова Любовь Николаевна,Новый Уренгой

Незапрограммированные перестройки генома.
К таким перестройкам относят перемещения мобильных генетических элементов
Незапрограммированные перестройки генома.
К таким перестройкам относят перемещения мобильных генетических элементов

Мобильные генетические элементы –МГЭ -последовательности ДНК, которые могут перемещаться внутри генома.
Существует
Мобильные генетические элементы –МГЭ -последовательности ДНК, которые могут перемещаться внутри генома.
Существует
 Движущие силы эволюции. Борьба за существование
Движущие силы эволюции. Борьба за существование Мой знак зодиака
Мой знак зодиака Технология каменной кладки
Технология каменной кладки Дмитрий Донской
Дмитрий Донской Презентация для педагогов дошкольных образовательных учреждений: Безопасность и здоровье детей дошкольного возраста.
Презентация для педагогов дошкольных образовательных учреждений: Безопасность и здоровье детей дошкольного возраста. Мультиплікатор: сутність, причини виникнення і наслідки для економіки
Мультиплікатор: сутність, причини виникнення і наслідки для економіки Простое слово - извините
Простое слово - извините Прокат спорт товаров
Прокат спорт товаров Глоссарий. Информационное право
Глоссарий. Информационное право Благодарность, смысл жизни. Сочинение на ОГЭ-2017. (Задание 15.3)
Благодарность, смысл жизни. Сочинение на ОГЭ-2017. (Задание 15.3) Требования к оформлению реквизитов организационно-распорядительных документов
Требования к оформлению реквизитов организационно-распорядительных документов Гармония. Адаптивная физическая культура
Гармония. Адаптивная физическая культура Конструирование одежды
Конструирование одежды Топографическая анатомия в области предплечья и кисти. (Занятие 3)
Топографическая анатомия в области предплечья и кисти. (Занятие 3) Электронное портфолио к конкурсу Учитель года 2013 3 часть
Электронное портфолио к конкурсу Учитель года 2013 3 часть Пример использования проматриц в задачах управления электромеханическими системами
Пример использования проматриц в задачах управления электромеханическими системами Военно-патриотический центр Русский витязь
Военно-патриотический центр Русский витязь Конфликтілердің алдын алу
Конфликтілердің алдын алу Сущность научного знания и его метод
Сущность научного знания и его метод Основные требования к студентам, проходящим производственную практику по получению профессиональных умений
Основные требования к студентам, проходящим производственную практику по получению профессиональных умений иваньков
иваньков 20231106_23-24_rk_5-e_po_rezultatu_dlya_sayta_chast_3
20231106_23-24_rk_5-e_po_rezultatu_dlya_sayta_chast_3 Презентация Конфликты в семье.
Презентация Конфликты в семье. Табиғи және техногенді радияциялық фон. Иондық сәулелену көзінің адам ағзасына әсерінің салдары
Табиғи және техногенді радияциялық фон. Иондық сәулелену көзінің адам ағзасына әсерінің салдары Если бы я была строителем .. 7 класс
Если бы я была строителем .. 7 класс Филогенез систем органов позвоночных животных
Филогенез систем органов позвоночных животных Экологические группы растений по отношению к воде
Экологические группы растений по отношению к воде Посвящение в химики
Посвящение в химики