- Пути обмена генетической информацией у микроорганизмов

Содержание
- 2. План лекции: Трансформация. Особенности переноса генетического материала при трансформации: компетентность, проникновение ДНК донора в клетку реципиента,
- 3. Пути обмена генетической информацией у микроорганизмов У прокариот существует три способа включения в геном чужеродной ДНК:
- 4. Передачу генов из клетки в клетку без всякого межклеточного контакта и без каких-либо переносчиков при помощи
- 5. Трансформация Для трансформации не требуется непосредственного контакта между двумя клетками. Способность ДНК проникать в клетку-реципиент зависит
- 6. Виды трансформации Бактериальная трансформация Естественная (конститутивная) (у представителей родов Bacillus, Rhizobium, Streptococcus) Индуцибельная (бактериальная клетка приобретает
- 7. Особенности переноса ДНК при трансформации Способность клетки к трансформации возможна при особом ее состоянии, которое называется
- 8. Модель процесса трансформации Экзогенная дцДНК связывается со специальными белковыми комплексами на поверхности клетки. В узнавании ДНК,
- 9. Стадии трансформации Длина трансформирующей ДНК должна быть от 10 до 20 тысяч н.п. Энергия, выделяющаяся при
- 10. Спонтанная трансформация Процесс трансформации может произвольно происходить в природе у некоторых видов бактерий, чаще грамположительных, когда
- 11. Трансфекция Трансформация известна и для эукариот. Однако на поверхности эукариотических клеток отсутствуют рецепторные сайты, и трансформирующую
- 12. Конъюгация В середине двадцатого века был описан половой процесс у бактерий (Дж. Ледерберг и Э.Татум, 1946).
- 13. Дж. Ледерберг и Э. Татум провели опыт с двумя мутантами Е. coli К12, каждый из которых
- 14. Доказательство возможности передачи ДНК при прямом контакте Эти мутанты не росли на минимальной питательной среде и
- 15. Особенности конъюгации Во время конъюгации образуется цитоплазматический мостик, по которому происходит перенос молекулы ДНК из одной
- 16. Стадии конъюгации Передача генетического материала при конъюгации начинается с расщепления ДНК в районе локализации F -фактора.
- 17. F+ и F- -клетки У кишечной палочки клетка-донор («мужская») имеет продолговатую форму, клетка-реципиент («женская») – изодиаметрическую.
- 18. Половой фактор F (от fertility - плодовитость) – фактор, передающийся при конъюгации из одной клетки в
- 19. F-фактор Фактор F представляет собой кольцевую двухцепочечную молекулу ДНК с массой 45∙106 Да. В качестве внехромосомного
- 20. F+ и F- -клетки F+ -доноры содержат в цитоплазме половой фактор – специфическую F–плазмиду. F–плазмида –
- 21. Клетки-доноры, обеспечивающие высокую частоту рекомбинаций, получили название клеток Hfr (от англ, high frequency of recombinants). Фактор
- 22. Особенности конъюгации Взаимоотношения между половыми типами Escherichia coli: Клетка F- может служить только реципиентом. При коньюгации
- 23. Процесс репликации у кишечной палочки продолжается 20 минут, а процесс конъюгации длится 3-5 минут. За это
- 24. Генетическая карта хромосомы E.coli В результате применения метода прерванной коньюгации, позволяющего выяснить временную последовательность переноса генов
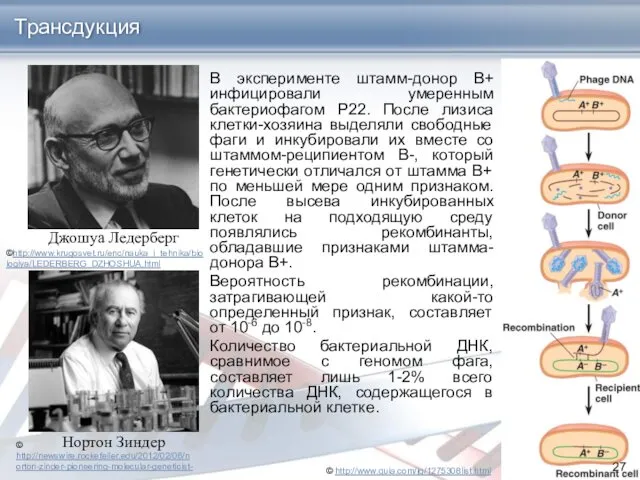
- 25. Трансдукция Трансдукция оказывается возможной, если в процессе размножения фага одна из частиц случайно захватит фрагмент бактериальной
- 26. Открытие трансдукции Явление трансдукции открыли в 1952 г. Н. Зиндер и Дж. Ледерберг. При трансдукции в
- 27. Джошуа Ледерберг Трансдукция Нортон Зиндер В эксперименте штамм-донор В+ инфицировали умеренным бактериофагом Р22. После лизиса клетки-хозяина
- 28. Виды трансдукции Различают общую (неспецифическую), ограниченную (специфическую) и абортивную трансдукцию. Общая трансдукция При общей трансдукции фрагменты
- 29. Виды трансдукции Ограниченная (специфическая) трансдукция Фаговая ДНК интегрирует в бактериальную хромосому с образованием профага. В процессе
- 30. Виды трансдукции При общей и ограниченной трансдукции донорская ДНК замещает гомологичные участки ДНК реципиента. Этот процесс
- 32. Скачать презентацию

План лекции:
Трансформация. Особенности переноса генетического материала при трансформации: компетентность, проникновение ДНК
План лекции:
Трансформация. Особенности переноса генетического материала при трансформации: компетентность, проникновение ДНК

Пути обмена генетической информацией у микроорганизмов
У прокариот существует три способа включения
Пути обмена генетической информацией у микроорганизмов
У прокариот существует три способа включения
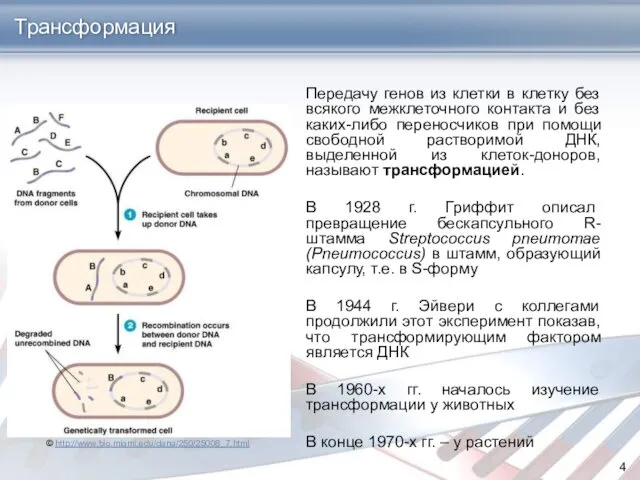
Передачу генов из клетки в клетку без всякого межклеточного контакта и
Передачу генов из клетки в клетку без всякого межклеточного контакта и

Трансформация
Для трансформации не требуется непосредственного контакта между двумя клетками.
Способность ДНК
Трансформация
Для трансформации не требуется непосредственного контакта между двумя клетками.
Способность ДНК
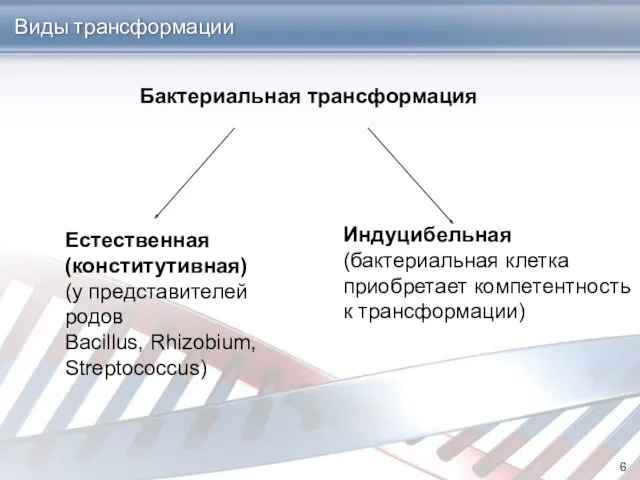
Виды трансформации
Бактериальная трансформация
Естественная
(конститутивная)
(у представителей родов
Bacillus, Rhizobium, Streptococcus)
Индуцибельная
(бактериальная клетка
приобретает компетентность
к
Виды трансформации
Бактериальная трансформация
Естественная
(конститутивная)
(у представителей родов
Bacillus, Rhizobium, Streptococcus)
Индуцибельная
(бактериальная клетка
приобретает компетентность
к

Особенности переноса ДНК при трансформации
Способность клетки к трансформации возможна при особом
Особенности переноса ДНК при трансформации
Способность клетки к трансформации возможна при особом
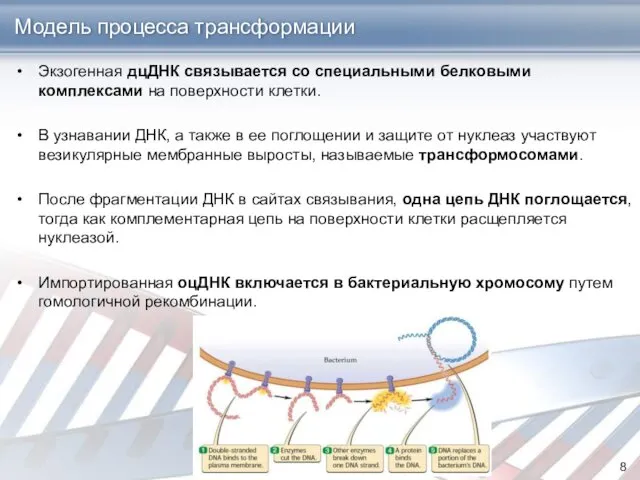
Модель процесса трансформации
Экзогенная дцДНК связывается со специальными белковыми комплексами на поверхности
Модель процесса трансформации
Экзогенная дцДНК связывается со специальными белковыми комплексами на поверхности

Стадии трансформации
Длина трансформирующей ДНК должна быть от 10 до 20
Стадии трансформации
Длина трансформирующей ДНК должна быть от 10 до 20

Спонтанная трансформация
Процесс трансформации может произвольно происходить в природе у некоторых
Спонтанная трансформация
Процесс трансформации может произвольно происходить в природе у некоторых

Трансфекция
Трансформация известна и для эукариот. Однако на поверхности эукариотических клеток отсутствуют
Трансфекция
Трансформация известна и для эукариот. Однако на поверхности эукариотических клеток отсутствуют

Конъюгация
В середине двадцатого века был описан половой процесс у бактерий (Дж.
Конъюгация
В середине двадцатого века был описан половой процесс у бактерий (Дж.

Дж. Ледерберг и Э. Татум провели опыт с двумя мутантами Е.

Доказательство возможности передачи ДНК при прямом контакте
Эти мутанты не росли на
Доказательство возможности передачи ДНК при прямом контакте
Эти мутанты не росли на

Особенности конъюгации
Во время конъюгации образуется цитоплазматический мостик, по которому происходит перенос
Особенности конъюгации
Во время конъюгации образуется цитоплазматический мостик, по которому происходит перенос

Стадии конъюгации
Передача генетического материала при конъюгации начинается с расщепления ДНК в
Стадии конъюгации
Передача генетического материала при конъюгации начинается с расщепления ДНК в
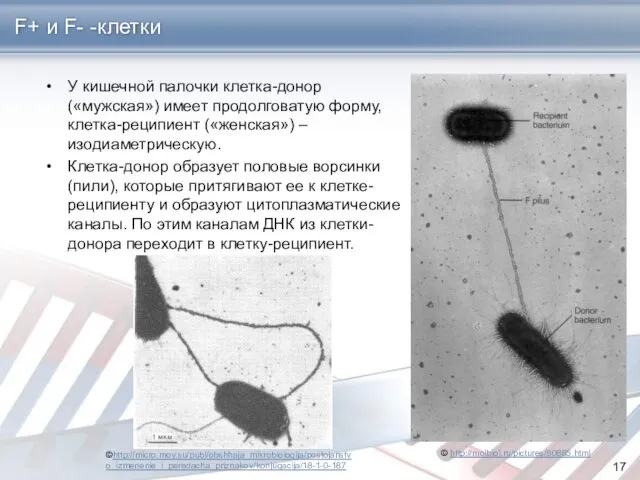
F+ и F- -клетки
У кишечной палочки клетка-донор («мужская») имеет продолговатую форму,
F+ и F- -клетки
У кишечной палочки клетка-донор («мужская») имеет продолговатую форму,
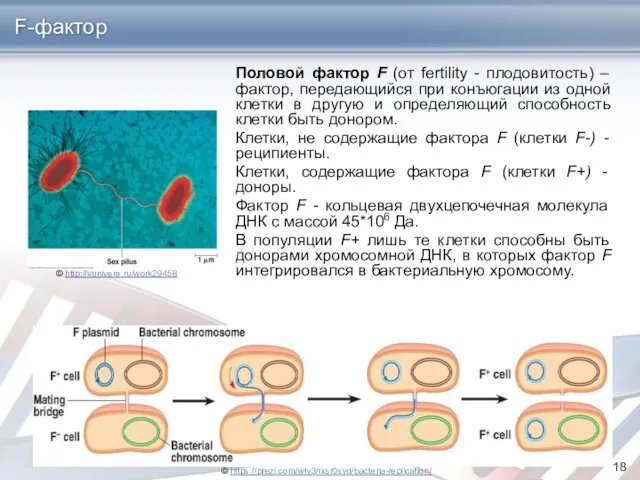
Половой фактор F (от fertility - плодовитость) – фактор, передающийся при
Половой фактор F (от fertility - плодовитость) – фактор, передающийся при

F-фактор
Фактор F представляет собой кольцевую двухцепочечную молекулу ДНК с массой 45∙106
F-фактор
Фактор F представляет собой кольцевую двухцепочечную молекулу ДНК с массой 45∙106

F+ и F- -клетки
F+ -доноры содержат в цитоплазме половой фактор –
F+ и F- -клетки
F+ -доноры содержат в цитоплазме половой фактор –
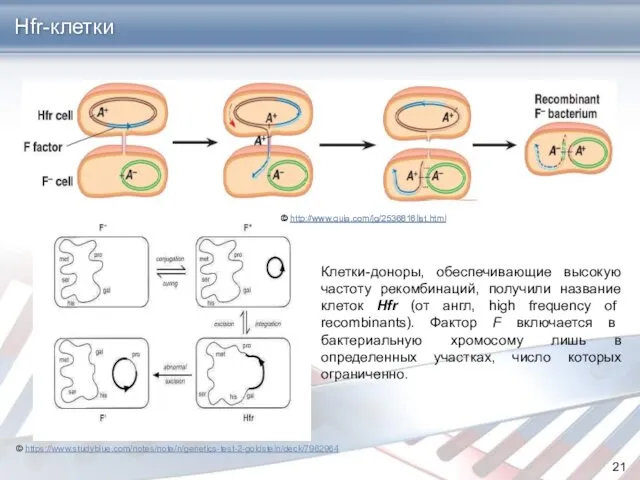
Клетки-доноры, обеспечивающие высокую частоту рекомбинаций, получили название клеток Hfr (от англ,
Клетки-доноры, обеспечивающие высокую частоту рекомбинаций, получили название клеток Hfr (от англ,
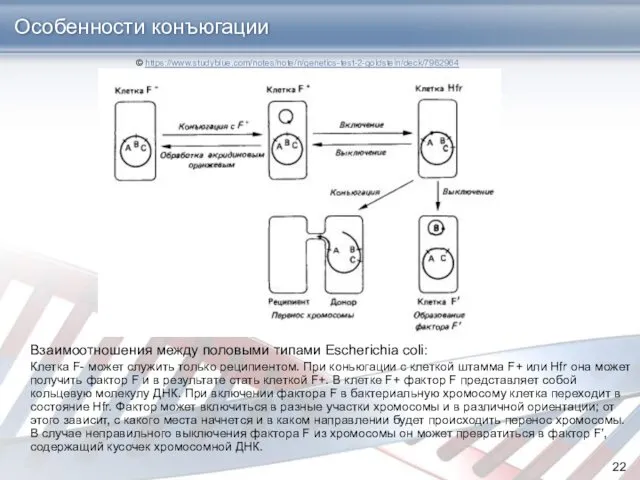
Особенности конъюгации
Взаимоотношения между половыми типами Escherichia coli:
Клетка F- может служить
Особенности конъюгации
Взаимоотношения между половыми типами Escherichia coli:
Клетка F- может служить

Процесс репликации у кишечной палочки продолжается 20 минут, а процесс конъюгации
Процесс репликации у кишечной палочки продолжается 20 минут, а процесс конъюгации

Генетическая карта хромосомы E.coli
В результате применения метода прерванной коньюгации, позволяющего выяснить
Генетическая карта хромосомы E.coli
В результате применения метода прерванной коньюгации, позволяющего выяснить
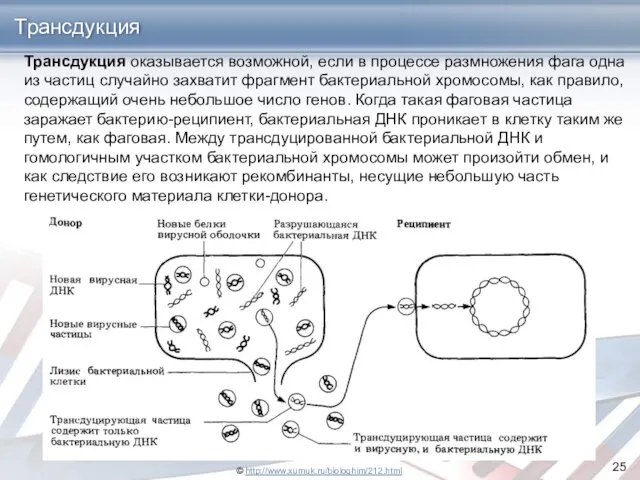
Трансдукция
Трансдукция оказывается возможной, если в процессе размножения фага одна из частиц
Трансдукция
Трансдукция оказывается возможной, если в процессе размножения фага одна из частиц

Открытие трансдукции
Явление трансдукции открыли в 1952 г. Н. Зиндер и Дж.
Открытие трансдукции
Явление трансдукции открыли в 1952 г. Н. Зиндер и Дж.
Джошуа Ледерберг
Трансдукция
Нортон Зиндер
В эксперименте штамм-донор В+ инфицировали умеренным бактериофагом Р22. После
Джошуа Ледерберг
Трансдукция
Нортон Зиндер
В эксперименте штамм-донор В+ инфицировали умеренным бактериофагом Р22. После

Виды трансдукции
Различают общую (неспецифическую), ограниченную (специфическую) и абортивную трансдукцию.
Общая трансдукция
При общей
Виды трансдукции
Различают общую (неспецифическую), ограниченную (специфическую) и абортивную трансдукцию.
Общая трансдукция
При общей

Виды трансдукции
Ограниченная (специфическая) трансдукция
Фаговая ДНК интегрирует в бактериальную хромосому с
Виды трансдукции
Ограниченная (специфическая) трансдукция
Фаговая ДНК интегрирует в бактериальную хромосому с

Виды трансдукции
При общей и ограниченной трансдукции донорская ДНК замещает гомологичные участки
Виды трансдукции
При общей и ограниченной трансдукции донорская ДНК замещает гомологичные участки
 Жизнь и творчество Ван Гога
Жизнь и творчество Ван Гога Материал к конкурсу Учитель года Москвы - 2012
Материал к конкурсу Учитель года Москвы - 2012 Процессуальное право: административная юрисдикция, конституционное судопроизводство
Процессуальное право: административная юрисдикция, конституционное судопроизводство Письмо-просьба
Письмо-просьба Ребусы
Ребусы Обозначение мягкости согласных звуков мягким знаком
Обозначение мягкости согласных звуков мягким знаком Исследовательская работа по теме Плутон
Исследовательская работа по теме Плутон Тамақ өнімдерін қайта өңдеу
Тамақ өнімдерін қайта өңдеу Лесная промышленность
Лесная промышленность Презентация по ритмике с детьми старшей группы
Презентация по ритмике с детьми старшей группы Спряжение глаголов (урок русского языка в 4 классе)
Спряжение глаголов (урок русского языка в 4 классе) Қара бидай мен тритикале сұрыптық ұнға дайындау принциптері
Қара бидай мен тритикале сұрыптық ұнға дайындау принциптері День народного единства
День народного единства Собственные торговые марки торговой сети Монетка
Собственные торговые марки торговой сети Монетка Особенность анестезиологического пособия в амбулаторных условиях
Особенность анестезиологического пособия в амбулаторных условиях Основные понятия гидроэнергетики
Основные понятия гидроэнергетики Моя семья
Моя семья Проект Молодежь сегодня, завтра…
Проект Молодежь сегодня, завтра… Поздравления к 8 марта
Поздравления к 8 марта ОРКСЭ Дружба - презентация.
ОРКСЭ Дружба - презентация. Обоснование программы ресурсосбережения промышленного предприятия
Обоснование программы ресурсосбережения промышленного предприятия Природа Урала
Природа Урала Особенности формирования нравственно-патриотических качеств у детей дошкольного возраста
Особенности формирования нравственно-патриотических качеств у детей дошкольного возраста Презентация Жиры
Презентация Жиры Екінші ретті дифференциалдық теңдеу үшін шекаралық шарттары бөлінген есеп арқылы туындайтын үйірткінің тұрпаты
Екінші ретті дифференциалдық теңдеу үшін шекаралық шарттары бөлінген есеп арқылы туындайтын үйірткінің тұрпаты Модернизация элементов подвешивания тяговой передачи электровоза
Модернизация элементов подвешивания тяговой передачи электровоза Совершенствование организации благоустройства и управления жилым фондом (на примере управы района Крюково города Москвы)
Совершенствование организации благоустройства и управления жилым фондом (на примере управы района Крюково города Москвы) Викторина Великая Отечественная Война
Викторина Великая Отечественная Война