- Регуляция биосинтеза белка. Механизмы генетической изменчивости

Содержание
- 2. Регуляция биосинтеза белка В 1961 г. французские исследователи Франсуа Жакоб и Жак Моно предложили теорию Lac-оперона,
- 3. Слева направо - Жакоб Франсуа, Жак Моно, Андре Львов лауреаты Нобелевской премии по физиологии и медицине
- 4. Координированный одним оператором одиночный ген или группа генов образуют оперон. Lac -оперон - участок ДНК, в
- 7. Репрессор блокирует ген-оператор → оперон не работает → транскрипция мРНК не происходит → синтез белка не
- 8. Лактозный оперон регулируется по механизму индукции Вещества, которые инактивируют репрессор, называются индукторами Вещества, переводящие его из
- 9. Лактоза – индуктор, присоединяясь к белку-репрессору, переводит его в неактивную форму, не способную связываться с О.
- 10. 3 фермента, участвующие в метаболизме лактозы: β-галактозидаза β-галактозидпермеаза β-галактозидтрансацетилаза
- 11. После распада лактозы белок-репрессор переходит снова в активную форму, способную связываться с О. Т.к. участки О
- 12. ДНК всех клеток организма идентична; >200 различных типов клеток Гены «домашнего хозяйства» ~ 20% Адаптивно регулируемые
- 13. Энхансеры – участки ДНК размером 10-20 пар оснований, присоединение к которым регуляторных белков активирует РНК-полимеразу и
- 15. Регуляция транскрипции Промоторы генов эукариот находятся под контролем специфических регуляторных участков на молекуле ДНК: ТАТА-, ЦААТ-,
- 16. К регуляторным участкам присоединяются комплексы белков с различными лигандами: цАМФ, стероидными гормонами, метаболитами, ионами металлов и
- 17. С энхансерами взаимодействуют индукторы С сайленсорами - репрессоры Это сложные белки, имеющие несколько доменов: «узнает» локус;
- 18. У эукариотов ведущая роль в экспрессии генов принадлежит стероидным, тиреоидным гормонам, факторам роста, инсулину, вторичным мессенджерам
- 19. Успехи генетики, молекулярной биологии и биохимии привели к формированию в 1990-х гг. двух новых фундаментальных дисциплин
- 20. Геном человека содержит 3,1 млрд пар нуклеотидов Только ~10% из них несут информацию В ходе выполнения
- 21. Теломеры - это концевые участки линейной молекулы ДНК, которые состоят из повторяющейся последовательности нуклеотидов, не кодирующих
- 22. Транспозоны — это участки ДНК организмов, способные к передвижению (транспозиции) и размножению в пределах генома. Транспозоны
- 23. Биохимические основы изменчивости и эволюции Движущей силой эволюции являются мутации – наследуемые изменения первичной структуры ДНК,
- 24. Механизмы возникновения Результат ошибок синтеза ДНК при репликации При репарации повреждения ДНК под влиянием внешних факторов
- 25. Виды мутаций Геномные – изменение всего генома; изменение числа хромосом (н-р, полиплоидия, трисомия (болезнь Дауна) и
- 26. Генные или точечные мутации Замены, при которых одно АО замещается на другое. Вставки, обеспечивающие внедрение в
- 27. Мутации по типу замены Без изменения смысла кодона (нейтральные или молчащие) – мутации, при к-рых замена
- 28. «Миссенс-мутации» - мутации с изменением смысла кодона, при которых замена одного АО приводит к замене АК
- 29. «Нонсенс мутации» - мутации, приводящие к образованию одного из терминирующих кодонов: УАА, УАГ, УГА УГГ→УАГ три
- 30. Мутации по типу вставки Без сдвига «рамки считывания» -происходит вставка лишних 3 НТ или с числом
- 31. → синтезируется полипептид со «случайной» последовательно-стью АК, т.к. изменяется смысл всех кодонов, следующих за местом мутации
- 32. Мутации по типу делеция Без сдвига «рамки считывания» - происходит выпадение 3 НТ или с числом
- 33. → синтезируется полипептид со «случайной» послед-ю АК, т.к. изменяется смысл всех кодонов, следующих за местом мутации
- 34. Частота мутаций 10-5-10-6 на 1 гамету за каждое поколение Может варьировать для разных генов от 10-4
- 35. Мутагенные факторы 1 - физические Лучистая энергия (УФО, рентген., γ-излучение, позитроны, нейтроны) УФО → образование ковалентных
- 36. Сущ-ет система репарации - группа ферментов, вырезающих тиминовые димеры, к-рые кодируются 9 генами. При повреждении любого
- 37. Фоновое излучение (космическое) - под его воздействием происходит отщепление АО. За сутки человек теряет ~ 50·103
- 38. 2 - химические АО в ДНК могут подвергаться различным воздействиям формамида (HCONH2), свободных радикалов, альдегидов, полициклических
- 40. Скачать презентацию

Регуляция биосинтеза белка
В 1961 г. французские исследователи Франсуа Жакоб и Жак
Регуляция биосинтеза белка
В 1961 г. французские исследователи Франсуа Жакоб и Жак

Слева направо - Жакоб Франсуа, Жак Моно, Андре Львов
лауреаты Нобелевской
Слева направо - Жакоб Франсуа, Жак Моно, Андре Львов
лауреаты Нобелевской

Координированный одним оператором одиночный ген или группа генов образуют оперон.
Lac -оперон
Координированный одним оператором одиночный ген или группа генов образуют оперон.
Lac -оперон



Репрессор блокирует ген-оператор → оперон не работает → транскрипция мРНК не
Репрессор блокирует ген-оператор → оперон не работает → транскрипция мРНК не

Лактозный оперон регулируется по механизму индукции
Вещества, которые инактивируют репрессор, называются индукторами
Лактозный оперон регулируется по механизму индукции
Вещества, которые инактивируют репрессор, называются индукторами

Лактоза – индуктор, присоединяясь к белку-репрессору, переводит его в неактивную форму,
Лактоза – индуктор, присоединяясь к белку-репрессору, переводит его в неактивную форму,

3 фермента, участвующие в метаболизме лактозы:
β-галактозидаза
β-галактозидпермеаза
β-галактозидтрансацетилаза
3 фермента, участвующие в метаболизме лактозы:
β-галактозидаза
β-галактозидпермеаза
β-галактозидтрансацетилаза

После распада лактозы белок-репрессор переходит снова в активную форму, способную связываться
После распада лактозы белок-репрессор переходит снова в активную форму, способную связываться

ДНК всех клеток организма идентична;
>200 различных типов клеток
Гены «домашнего хозяйства» ~
ДНК всех клеток организма идентична;
>200 различных типов клеток
Гены «домашнего хозяйства» ~

Энхансеры – участки ДНК размером 10-20 пар оснований, присоединение к
Энхансеры – участки ДНК размером 10-20 пар оснований, присоединение к
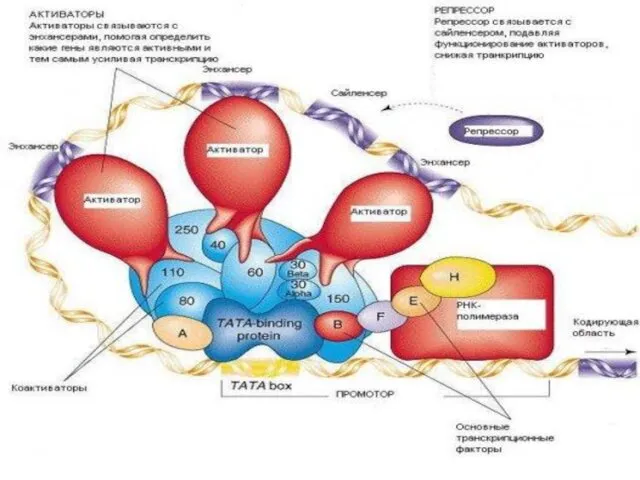

Регуляция транскрипции
Промоторы генов эукариот находятся
под контролем специфических
регуляторных участков на
молекуле
Регуляция транскрипции
Промоторы генов эукариот находятся
под контролем специфических
регуляторных участков на
молекуле

К регуляторным участкам
присоединяются
комплексы белков
с различными лигандами:
цАМФ, стероидными
К регуляторным участкам
присоединяются
комплексы белков
с различными лигандами:
цАМФ, стероидными

С энхансерами взаимодействуют индукторы
С сайленсорами - репрессоры
Это сложные белки, имеющие несколько
С энхансерами взаимодействуют индукторы
С сайленсорами - репрессоры
Это сложные белки, имеющие несколько

У эукариотов ведущая роль в экспрессии генов принадлежит стероидным, тиреоидным гормонам,

Успехи генетики, молекулярной биологии и биохимии привели к формированию в 1990-х
Успехи генетики, молекулярной биологии и биохимии привели к формированию в 1990-х

Геном человека содержит
3,1 млрд пар нуклеотидов
Только ~10% из них несут
Геном человека содержит
3,1 млрд пар нуклеотидов
Только ~10% из них несут

Теломеры - это концевые участки линейной молекулы ДНК, которые состоят из
Теломеры - это концевые участки линейной молекулы ДНК, которые состоят из

Транспозоны — это участки ДНК организмов, способные к передвижению (транспозиции) и размножению в пределах генома.
Транспозоны — это участки ДНК организмов, способные к передвижению (транспозиции) и размножению в пределах генома.

Биохимические основы
изменчивости и эволюции
Движущей силой эволюции являются мутации – наследуемые
Биохимические основы
изменчивости и эволюции
Движущей силой эволюции являются мутации – наследуемые

Механизмы возникновения
Результат ошибок синтеза ДНК при репликации
При репарации повреждения ДНК под
Механизмы возникновения
Результат ошибок синтеза ДНК при репликации
При репарации повреждения ДНК под

Виды мутаций
Геномные – изменение всего генома; изменение числа хромосом (н-р, полиплоидия,
Виды мутаций
Геномные – изменение всего генома; изменение числа хромосом (н-р, полиплоидия,

Генные или точечные мутации
Замены, при которых одно АО замещается на
Генные или точечные мутации
Замены, при которых одно АО замещается на

Мутации по типу замены
Без изменения смысла кодона (нейтральные или молчащие) –
Мутации по типу замены
Без изменения смысла кодона (нейтральные или молчащие) –

«Миссенс-мутации» - мутации с изменением смысла кодона, при которых замена одного
«Миссенс-мутации» - мутации с изменением смысла кодона, при которых замена одного

«Нонсенс мутации» - мутации, приводящие к образованию одного из терминирующих кодонов:
«Нонсенс мутации» - мутации, приводящие к образованию одного из терминирующих кодонов:

Мутации по типу вставки
Без сдвига «рамки считывания» -происходит вставка лишних
Мутации по типу вставки
Без сдвига «рамки считывания» -происходит вставка лишних

→ синтезируется полипептид со «случайной» последовательно-стью АК, т.к. изменяется смысл
→ синтезируется полипептид со «случайной» последовательно-стью АК, т.к. изменяется смысл

Мутации по типу делеция
Без сдвига «рамки считывания» - происходит выпадение 3
Мутации по типу делеция
Без сдвига «рамки считывания» - происходит выпадение 3

→ синтезируется полипептид со «случайной» послед-ю АК, т.к. изменяется смысл
→ синтезируется полипептид со «случайной» послед-ю АК, т.к. изменяется смысл

Частота мутаций 10-5-10-6 на 1 гамету за каждое поколение
Может варьировать для
Частота мутаций 10-5-10-6 на 1 гамету за каждое поколение
Может варьировать для

Мутагенные факторы
1 - физические
Лучистая энергия (УФО, рентген., γ-излучение, позитроны, нейтроны)
УФО
Мутагенные факторы
1 - физические
Лучистая энергия (УФО, рентген., γ-излучение, позитроны, нейтроны)
УФО

Сущ-ет система репарации - группа ферментов, вырезающих тиминовые димеры, к-рые
кодируются
Сущ-ет система репарации - группа ферментов, вырезающих тиминовые димеры, к-рые кодируются

Фоновое излучение (космическое) - под его воздействием происходит отщепление АО. За
Фоновое излучение (космическое) - под его воздействием происходит отщепление АО. За

2 - химические
АО в ДНК могут подвергаться различным воздействиям формамида (HCONH2),
2 - химические
АО в ДНК могут подвергаться различным воздействиям формамида (HCONH2),
 20231213_oge_16_teoriya
20231213_oge_16_teoriya Catalog of models
Catalog of models Леонард Эйлер 1707-1783
Леонард Эйлер 1707-1783 Влияние инфляции на данные финансовой отчетности
Влияние инфляции на данные финансовой отчетности Химические уравнения. Типы химических реакций
Химические уравнения. Типы химических реакций Презентация Семейные ценности. Традиции семьи
Презентация Семейные ценности. Традиции семьи Бонусная программа лояльности Клуб покупателей 5 Элемент
Бонусная программа лояльности Клуб покупателей 5 Элемент Факторы, определяющие индивидуальную реакцию на изменения Индивидуальные предпочтения и изменение
Факторы, определяющие индивидуальную реакцию на изменения Индивидуальные предпочтения и изменение Государственные символы России. Конституция РФ
Государственные символы России. Конституция РФ Развитие мелкой моторики у детей дошкольного возраста на занятиях по тестопластике.
Развитие мелкой моторики у детей дошкольного возраста на занятиях по тестопластике. Методология комплексной оценки состояния здоровья населения. Медико-социальные аспекты смертности
Методология комплексной оценки состояния здоровья населения. Медико-социальные аспекты смертности Патология зрительной системы
Патология зрительной системы Диалог (1 класс)
Диалог (1 класс) Работа над дикцией и артикуляцией: игровые приемы
Работа над дикцией и артикуляцией: игровые приемы Проект Дорожная азбука Диск
Проект Дорожная азбука Диск Презентация Свойства веществ
Презентация Свойства веществ Логопедическое занятие - игра Зима. Зимние забавы для детей старшей группы.
Логопедическое занятие - игра Зима. Зимние забавы для детей старшей группы. Занятие ГПД. В гостях у Снегурочки.
Занятие ГПД. В гостях у Снегурочки. Презентация к логопедическому занятию Полевые цветы
Презентация к логопедическому занятию Полевые цветы Современные модели образовательного процесса ДОУ
Современные модели образовательного процесса ДОУ Методы увязки
Методы увязки Химические элементы
Химические элементы Живые снежинки
Живые снежинки Никель қаптамаларын алу жолдары
Никель қаптамаларын алу жолдары НОУ. Исследовательская работа''Влияние агроклиматических ресурсов на сельскохозяйственную деятельность племзавода „Большемурашкинский’’. Решение продовольственной проблемы в России''.Часть 2.
НОУ. Исследовательская работа''Влияние агроклиматических ресурсов на сельскохозяйственную деятельность племзавода „Большемурашкинский’’. Решение продовольственной проблемы в России''.Часть 2. Необычные растения и животные в природе.
Необычные растения и животные в природе. Музейный урок История утюга
Музейный урок История утюга phrasal verbs Unit 7
phrasal verbs Unit 7