- Слуховой анализатор

Содержание
- 2. Слуховой анализатор Второй по значению и объему информации, после зрительного. Эволюционно, он возник и развивался вместе
- 3. защитная резонаторная локализация источника звука выравнивание давления обеспечение проведения звука рецепция звука
- 4. Наружное ухо представлено ушной раковиной и наружным слуховым проходом. Ушная раковина — эластический хрящ сложной формы,
- 6. передача и усиление звука в среднем ухе Барабанная перепонка, воспринимая звуковые колебания, передает их на систему
- 7. Защитный акустический рефлекс При громком звуке коэффициент передачи среднего уха резко снижается, благодаря сокращению двух мышц
- 8. Защитный акустический рефлекс
- 9. Внутреннее ухо Внутреннее ухо - лабиринт и каналы, заполненные жидкостью. Улитка (спирально закрученный костный канал имеет
- 10. Внутреннее ухо Внутреннее ухо - лабиринт и каналы, заполненные жидкостью. Улитка (спирально закрученный костный канал имеет
- 11. Жидкости внутреннего уха Эндолимфа средней лестницы сообщается с эндолимфой вестибулярного органа и имеет тот же состав.
- 12. Жидкости внутреннего уха
- 13. Эндокохлеарный потенциал Между эндолимфой и перилимфой существует электрический потенциал — около +80 мВ (внутриулитковый) или эндокохлеарный
- 14. Эндокохлеарный потенциал Волосковые клетки поляризованы эндокохлеарным потенциалом до критического уровня, что повышает их чувствительность к механическому
- 15. Эндокохлеарный потенциал
- 16. Кортиев орган Основная мембрана, в развёрнутом виде, имеет в длину - 3,5 см. Её ширина возрастает
- 18. Текториальная мембрана
- 19. БАЗИЛЯРНАЯ МЕМБРАНА Текториальная мембрана
- 20. Рецепторный потенциал Волосковые клетки контактируют с текторальной мембраной. При передаче звука – базальная мембрана движется и
- 21. Рецепторный потенциал Это увеличивает электрический градиент между вне- и внутриклеточной средой и вызывает движение К+ внутрь
- 22. Такая значительная, исходная мембранная поляризация волосковых клеток, обеспечивает их высокую чувствительность к слабым звуковым колебаниям. Изменение
- 23. Электрические явления в улитке Представлены в форме 5 потенциалов: 1)МП рецепторной клетки (волосковой) и 2) потенциал
- 24. Постоянные потенциалы улитки МП волосковой клетки (-80 мВ) Потенциал эндолимфы (+80 мВ)
- 25. Микрофонный эффект Если ввести в улитку электроды, соединить их с динамиком через усилитель и подействовать на
- 26. Суммационный потенциал (СП) В ответ на сильные звуки большой частоты (высокие тоны) отмечают стойкий сдвиг исходной
- 27. Микрофонный и суммационный потенциалы рассматривают как суммарные рецепторные потенциалы волосковых клеток. Имеются данные, что отрицательный СП
- 29. потенциал слухового нерва ПД ПД регистрируются при отведении от волокон слухового нерва. Их частота зависит от
- 31. ВОЛОСКОВАЯ КЛЕТКА Каждая волосковая клетка настроена на определённую частоту, клетки настроенные на низкую частоту – располагаются
- 32. Особенности функций волосковых клеток Собственно рецепторами (звука) являются внутренние волосковые клетки, от них отходят 95 %
- 33. Последовательность сенсорного преобразования в органе слуха: 1. Колебания овального окна передаются на жидкости и мембраны внутреннего
- 34. 12 500 наружных волосковых клеток иннервированы 2 500 нейронами 2 типа спирального ганглия, каждый нейрон контактирует
- 35. Афферентная иннервация волосковых клеток
- 36. Базилярная мембрана Длина базилярной мебраны – 3,5 см.
- 37. Базилярная мембрана Градиент жёсткости базилярной мембраны определяет её способность к фильтрации низкочастотных звуков. Жесткость мембраны снижается
- 38. Бегущая волна Из-за условий фиксации базилярная мембрана движется в продольном и поперечном направлениях
- 39. Пространственный принцип кодирования частоты
- 40. Базилярная мембрана высокочастотные колебания пробегают очень короткое расстояние длинные волны распространяются далеко
- 41. Механика базилярной мембраны высокие частоты низкие частоты Принцип «места»
- 42. Высота звука — это субъективное восприятие частоты звуковых колебаний. Человеческое ухо воспринимает частоты в диапазоне примерно
- 43. Рецепторные потенциалы волосковых клеток
- 44. СЛУХОВЫЕ ОЩУЩЕНИЯ Диапазон частот, который воспринимает человек, включает около 10 октав музыкальной шкалы (от 16 Гц
- 45. Абсолютный порог слуховой чувствительности — минимальная сила звука, которую слышит человек в 50% случаев его предъявления.
- 48. Сила звука Диапазон силы звука, воспринимаемый человеческим ухом, огромен (болевой порог в 1013 раз выше порога
- 49. Сила звука кодируется как пространственным (изменением числа возбужденных волосковых клеток), так и временным (изменением частоты импульсации
- 50. при раздражении волосков в ответ на звуковое колебание наружные волосковые клетки дополнительно раскачивают основную мембрану в
- 51. СЛУХОВЫЕ ПУТИ И ЦЕНТРЫ Афферентные нервные волокна от улитки входят в спиральный ганглий и от него
- 52. СЛУХОВЫЕ ПУТИ И ЦЕНТРЫ От ядер верхней оливы слуховые пути поднимаются вверх через латеральный лемнисковый путь;
- 53. СЛУХОВЫЕ ПУТИ И ЦЕНТРЫ Далее слуховой путь следует к медиальным коленчатым телам (МКТ), где все волокна
- 54. СЛУХОВЫЕ ПУТИ И ЦЕНТРЫ Базилярная мембрана улитки на всех уровнях слухового пути представлена в форме определённых
- 57. СЛУХОВАЯ КОРА Первичная слуховая кора непосредственно получает сигналы от внутреннего (медиального) коленчатого тела. Слуховая ассоциативная область
- 58. В каждой из 6 тонотопических карт звуки высокой частоты возбуждают нейроны в задней части карты, в
- 59. СЛУХОВАЯ КОРА Первичная слуховая кора непосредственно получает сигналы от внутреннего (медиального) коленчатого тела. Слуховая ассоциативная область
- 60. Проекционные области слуховой коры располагаются не только в верхней части верхней височной извилины, но и простираются
- 61. Тонотопические карты Одна большая карта в первичной слуховой коре почти целиком дискриминирует звуки, которые субъекту кажутся
- 62. Тонотопические карты Диапазон звуковой частоты, на которую отвечают нейроны слуховой коры уже, чем для нейронов спирального
- 63. Тонотопические карты Многие нейроны слуховой коры, особенно в слуховой ассоциативной коре, отвечают не только на специфические
- 64. Бинауральный слух Человек определяет звук, двумя путями: 1. Временем задержки между поступлением звука в одно ухо
- 65. Бинауральный слух . 2. Различием между интенсивностью звуков в двух ушах: при высоких частотах звука размер
- 67. Такое тонкое сравнение минимальной разницы во времени прихода звука осуществляется ЦНС в точках, где осуществляется конвергенция
- 73. Вестибулярный анализатор
- 74. Вестибулярный анализатор Совокупность механорецепторов и нервных структур, обеспечивающих восприятие и анализ ориентации тела в пространстве. Рефлексы,
- 75. Вестибулярный анализатор
- 76. вестибулярный аппарат
- 77. Рецепторный аппарат В преддверии находится 2 мешочка – круглый (sacculus) и эллиптический (маточка, utriculus). В них
- 78. Рецепторный аппарат Возбуждение клеток происходит при скольжении отолитовой мембраны по волоскам, то есть их сгибании. Рецепторные
- 79. Гребешки и пятна — воспринимающие области вестибулярного органа — содержат рецепторные волосковые клетки. В полукружных каналах
- 80. В утрикулисе и саккулисе комплексы волосковых клеток собраны в группы овальной формы –макулы.
- 81. Пятно эллиптического мешочка лежит горизонтально и обеспечивает информацию при движении головы из стороны в сторону. Пятно
- 83. м
- 84. Сила инерции, которая возникает при линейных ускорениях: во время движений "вверх - вниз", "вперед-назад“ действует на
- 85. При направлении раздражающего воздействия от стереоцилий к киноцилии волосковая клетка возбуждается (происходит деполяризация). При противоположном направлении
- 86. Рецепторный потенциал Развитие рецепторного потенциала происходит при изгибе волосков в правильном направлении. При этом возникает активация
- 87. Рецепторный потенциал
- 89. Детекция поворота головы Полукружные каналы детектируют поворот или вращение головы. Когда голова начинает поворачиваться в каком-либо
- 90. Реакции организма, вызванные стимуляцией полукружных каналов Стимуляция полукружных каналов вызывает субъективные ощущения в виде головокружения, тошноты
- 91. ПРОЕКЦИОННЫЕ ПУТИ ВЕСТИБУЛЯРНОГО АППАРАТА Вестибулярная ветвь VIII нерва образована отростками примерно 19 тыс. биполярных нейронов, образующих
- 92. ПРОЕКЦИОННЫЕ ПУТИ ВЕСТИБУЛЯРНОГО АППАРАТА Аксоны нервных клеток второго порядка связаны со спинным мозгом (преддверно–спинномозговой путь, оливо–спинномозговой
- 93. ПРОЕКЦИОННЫЕ ПУТИ ВЕСТИБУЛЯРНОГО АППАРАТА Есть два основных пути поступления вестибулярных сигналов в кору : 1)прямой —
- 95. Вестибулосоматические реакции К вестибулосоматическим реакциям относят вестибулярный нистагм - ритмические скачкообразные движения глазных яблок. Он возникает
- 96. Обонятельная сенсорная система
- 97. Классификация запахов по Эймуэру Первичные или основные: - камфорный - (камфора, 1,8-цинеол) - острый или едкий
- 98. Воспринимающие обонятельные структуры образуют в слизистой оболочке носа специализированную область — парную обонятельную выстилку (обонятельное поле),
- 99. ВТЯГИВАНИЕ ВОЗДУХА Область, содержащая обонятельные рецепторы, плохо вентилируется. Обычно воздух спокойно движется над носовыми раковинами во
- 100. Рецепторный аппарат Обонятельный эпителий содержит опорные эпителиальные клетки и расположенные между ними примерно 100 млн рецепторных
- 101. Рецепторный аппарат От булавы отходит от 8 до 40 тонких обонятельных волосков — ресничек. Именно в
- 102. Рецепторный аппарат Продолжительность жизни обонятельных клеток — 30–35 дней. Они относятся к обновляющейся клеточной популяции. Предшественники
- 103. В обонятельной луковице человека различают шесть слоев, которые располагаются концентрически, считая от поверхности: I слой -
- 104. IV слой – тел митральных клеток. Это крупные нейроны (диаметр сомы не менее 30 мкм) с
- 105. ОБОНЯТЕЛЬНАЯ ЛУКОВИЦА В обонятельной луковице аксоны рецепторных клеток образуют синапсы с дендритами митральных и пучковых клеток,
- 106. ОБОНЯТЕЛЬНАЯ ЛУКОВИЦА В следующем слое обонятельной луковицы дендриты митральных, зернистых и пучковых клеток образуют реципрокные синапсы.
- 108. Скачать презентацию
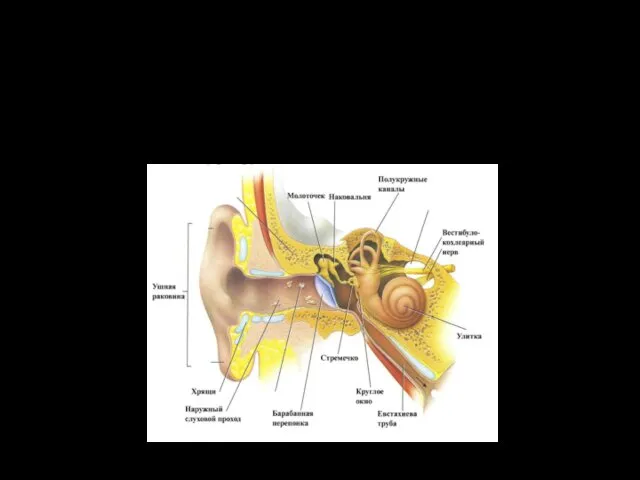
Слуховой анализатор
Второй по значению и объему информации, после зрительного.
Эволюционно, он возник
Слуховой анализатор
Второй по значению и объему информации, после зрительного.
Эволюционно, он возник
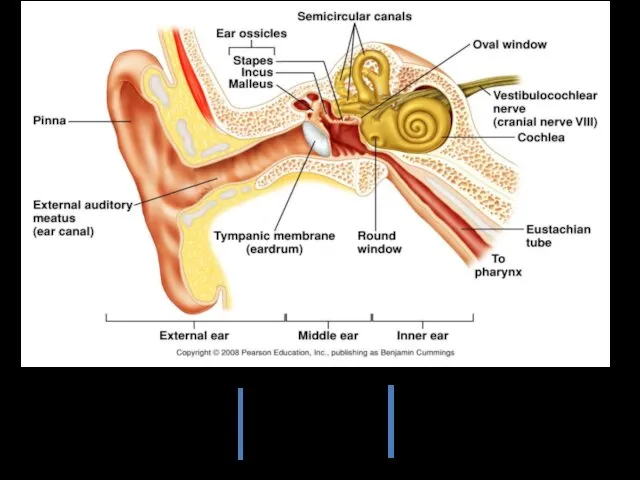
защитная
резонаторная
локализация источника звука
выравнивание давления
обеспечение проведения
звука
рецепция звука
защитная
резонаторная
локализация источника звука
выравнивание давления
обеспечение проведения
звука
рецепция звука

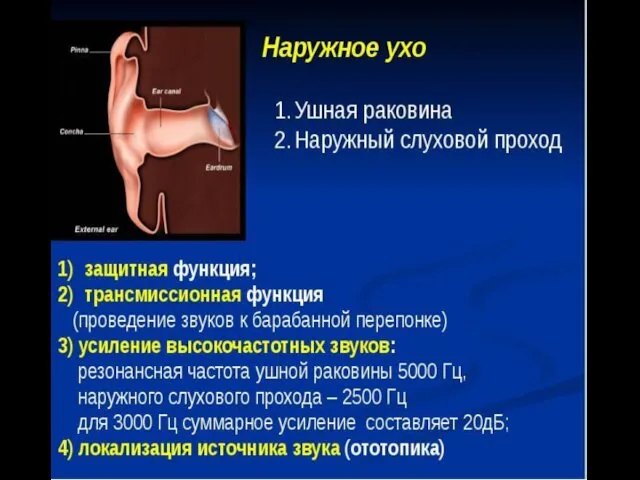
Наружное ухо представлено ушной раковиной и наружным слуховым проходом.
Ушная раковина — эластический
Наружное ухо представлено ушной раковиной и наружным слуховым проходом.
Ушная раковина — эластический

передача и усиление звука в среднем ухе
Барабанная перепонка, воспринимая звуковые
передача и усиление звука в среднем ухе
Барабанная перепонка, воспринимая звуковые

Защитный акустический рефлекс
При громком звуке коэффициент передачи среднего уха резко снижается,
Защитный акустический рефлекс
При громком звуке коэффициент передачи среднего уха резко снижается,
Защитный акустический рефлекс
Защитный акустический рефлекс

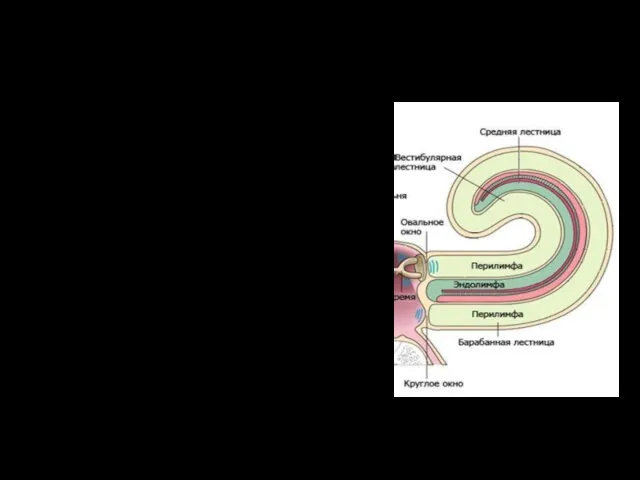
Внутреннее ухо
Внутреннее ухо - лабиринт и каналы, заполненные жидкостью.
Улитка (спирально
Внутреннее ухо
Внутреннее ухо - лабиринт и каналы, заполненные жидкостью.
Улитка (спирально
Внутреннее ухо
Внутреннее ухо - лабиринт и каналы, заполненные жидкостью.
Улитка (спирально
Внутреннее ухо
Внутреннее ухо - лабиринт и каналы, заполненные жидкостью.
Улитка (спирально

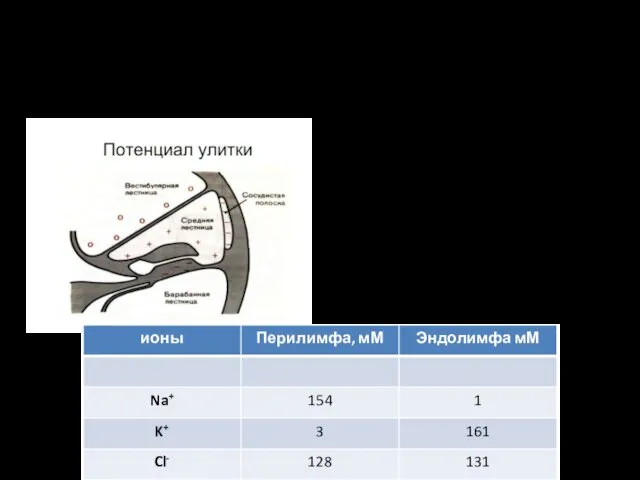
Жидкости внутреннего уха
Эндолимфа средней лестницы сообщается с эндолимфой вестибулярного органа и
Жидкости внутреннего уха
Эндолимфа средней лестницы сообщается с эндолимфой вестибулярного органа и
Жидкости внутреннего уха
Жидкости внутреннего уха

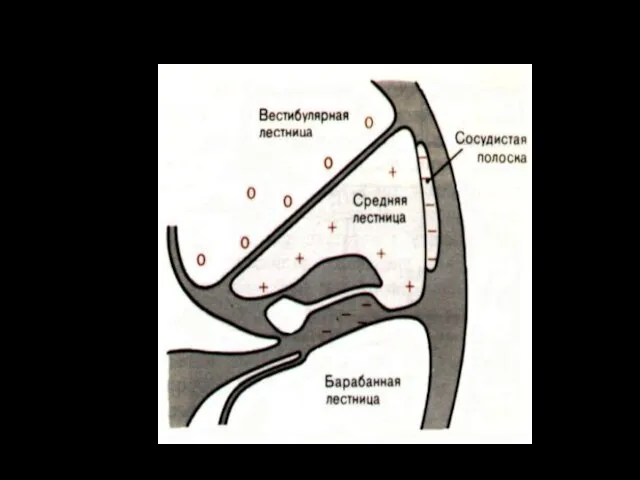
Эндокохлеарный потенциал
Между эндолимфой и перилимфой существует электрический потенциал — около +80 мВ (внутриулитковый)
Эндокохлеарный потенциал
Между эндолимфой и перилимфой существует электрический потенциал — около +80 мВ (внутриулитковый)

Эндокохлеарный потенциал
Волосковые клетки поляризованы эндокохлеарным потенциалом до критического уровня, что повышает
Эндокохлеарный потенциал
Волосковые клетки поляризованы эндокохлеарным потенциалом до критического уровня, что повышает
Эндокохлеарный потенциал
Эндокохлеарный потенциал

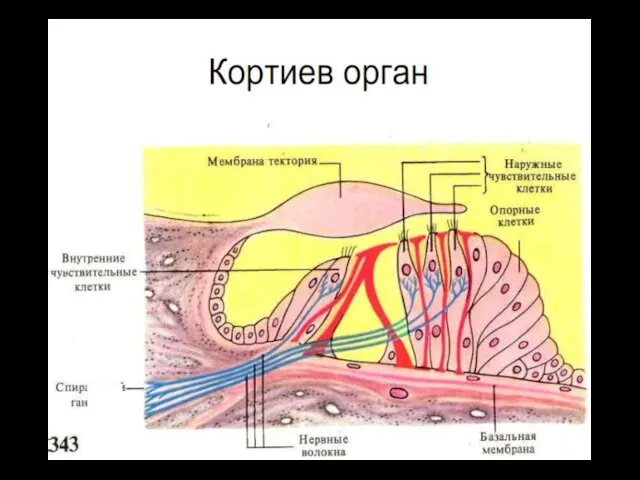
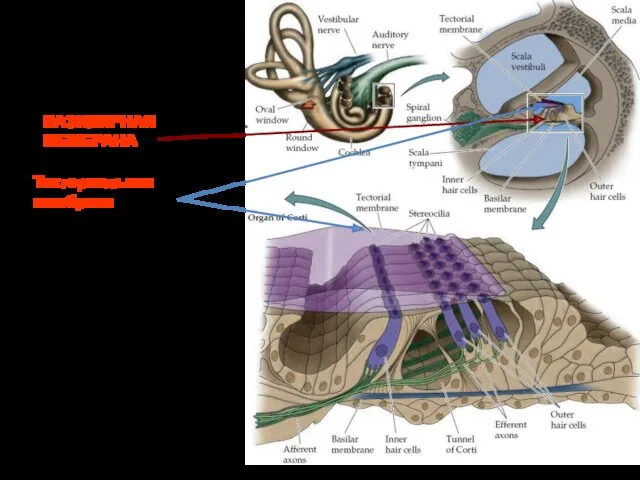
Кортиев орган
Основная мембрана, в развёрнутом виде, имеет в длину - 3,5
Кортиев орган
Основная мембрана, в развёрнутом виде, имеет в длину - 3,5
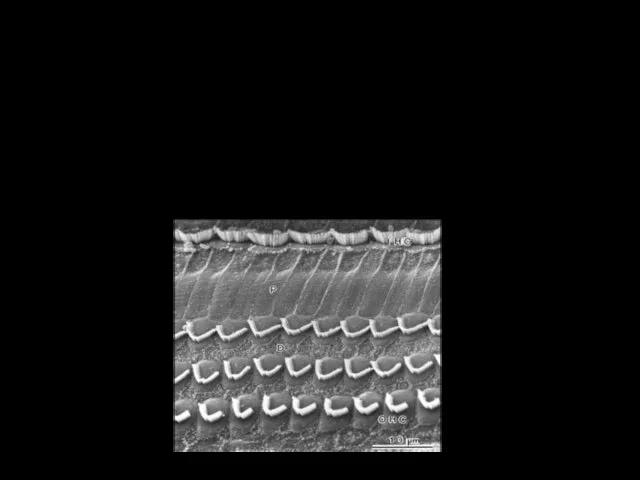
Текториальная
мембрана
Текториальная
мембрана
БАЗИЛЯРНАЯ МЕМБРАНА
Текториальная
мембрана
БАЗИЛЯРНАЯ МЕМБРАНА
Текториальная
мембрана

Рецепторный потенциал
Волосковые клетки контактируют с текторальной мембраной. При передаче звука –
Рецепторный потенциал
Волосковые клетки контактируют с текторальной мембраной. При передаче звука –

Рецепторный потенциал
Это увеличивает электрический градиент между вне- и внутриклеточной средой и
Рецепторный потенциал
Это увеличивает электрический градиент между вне- и внутриклеточной средой и

Такая значительная, исходная мембранная поляризация волосковых клеток, обеспечивает их высокую чувствительность
Такая значительная, исходная мембранная поляризация волосковых клеток, обеспечивает их высокую чувствительность

Электрические явления в улитке
Представлены в форме 5 потенциалов:
1)МП рецепторной клетки (волосковой)
Электрические явления в улитке
Представлены в форме 5 потенциалов:
1)МП рецепторной клетки (волосковой)
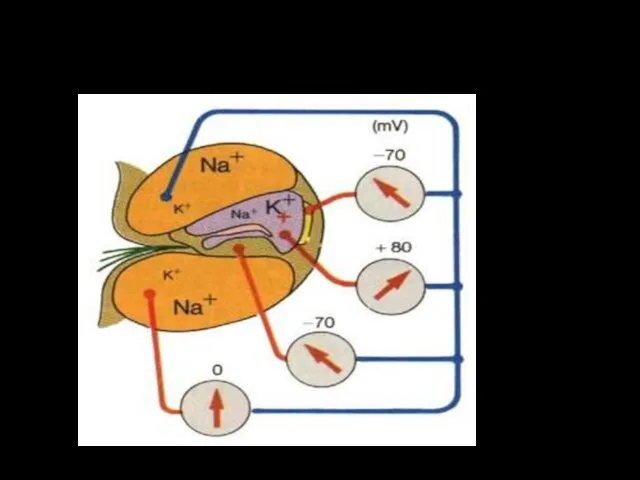
Постоянные потенциалы улитки
МП волосковой клетки (-80 мВ)
Потенциал
эндолимфы (+80 мВ)
Постоянные потенциалы улитки
МП волосковой клетки (-80 мВ)
Потенциал
эндолимфы (+80 мВ)

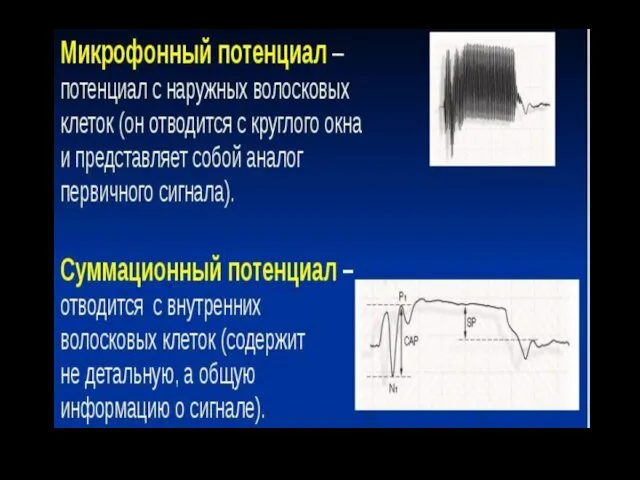
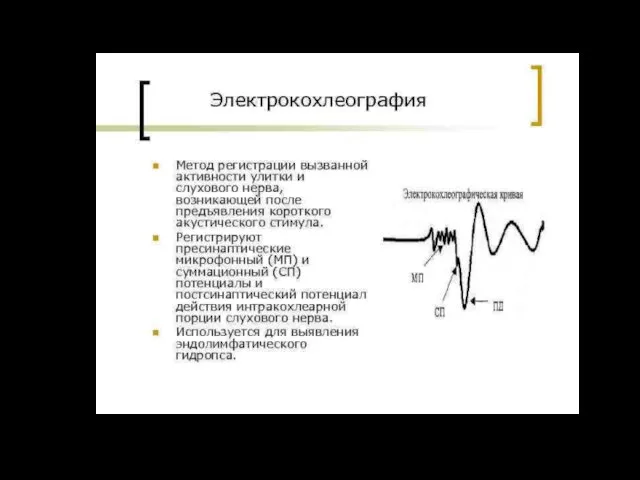
Микрофонный эффект
Если ввести в улитку электроды, соединить их с динамиком через
Микрофонный эффект
Если ввести в улитку электроды, соединить их с динамиком через

Суммационный потенциал (СП)
В ответ на сильные звуки большой частоты (высокие тоны)
Суммационный потенциал (СП)
В ответ на сильные звуки большой частоты (высокие тоны)

Микрофонный и суммационный потенциалы рассматривают как суммарные рецепторные потенциалы волосковых клеток.
Микрофонный и суммационный потенциалы рассматривают как суммарные рецепторные потенциалы волосковых клеток.

потенциал слухового нерва ПД
ПД регистрируются при отведении от волокон слухового нерва.
потенциал слухового нерва ПД
ПД регистрируются при отведении от волокон слухового нерва.

ВОЛОСКОВАЯ КЛЕТКА
Каждая волосковая клетка настроена на определённую частоту, клетки настроенные на
ВОЛОСКОВАЯ КЛЕТКА
Каждая волосковая клетка настроена на определённую частоту, клетки настроенные на

Особенности функций волосковых клеток
Собственно рецепторами (звука) являются внутренние волосковые клетки, от них
Особенности функций волосковых клеток
Собственно рецепторами (звука) являются внутренние волосковые клетки, от них

Последовательность сенсорного преобразования в органе слуха:
1. Колебания овального окна передаются на жидкости
Последовательность сенсорного преобразования в органе слуха:
1. Колебания овального окна передаются на жидкости

12 500 наружных волосковых клеток иннервированы 2 500 нейронами 2 типа
12 500 наружных волосковых клеток иннервированы 2 500 нейронами 2 типа
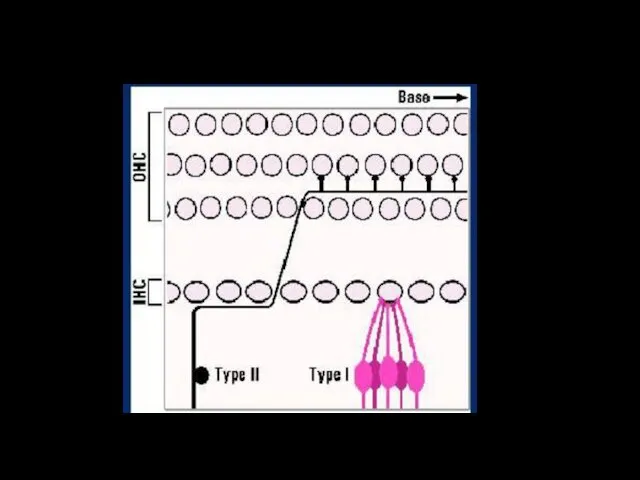
Афферентная иннервация волосковых клеток
Афферентная иннервация волосковых клеток
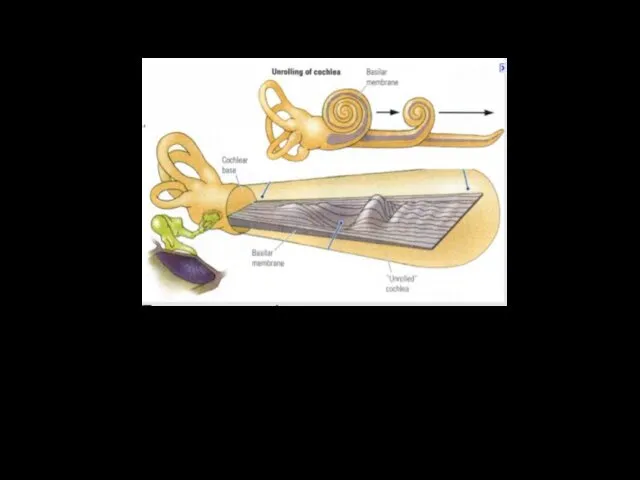
Базилярная мембрана
Длина базилярной мебраны – 3,5 см.
Базилярная мембрана
Длина базилярной мебраны – 3,5 см.

Базилярная мембрана
Градиент жёсткости базилярной мембраны определяет её способность к фильтрации низкочастотных
Базилярная мембрана
Градиент жёсткости базилярной мембраны определяет её способность к фильтрации низкочастотных
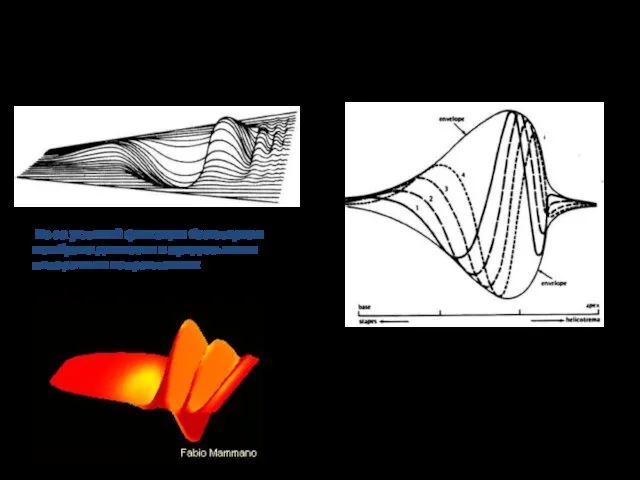
Бегущая волна
Из-за условий фиксации базилярная мембрана движется в продольном и
Бегущая волна
Из-за условий фиксации базилярная мембрана движется в продольном и
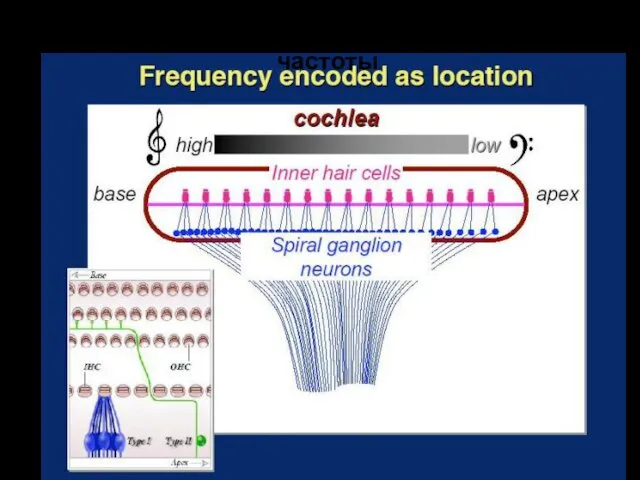
Пространственный принцип кодирования частоты
Пространственный принцип кодирования частоты
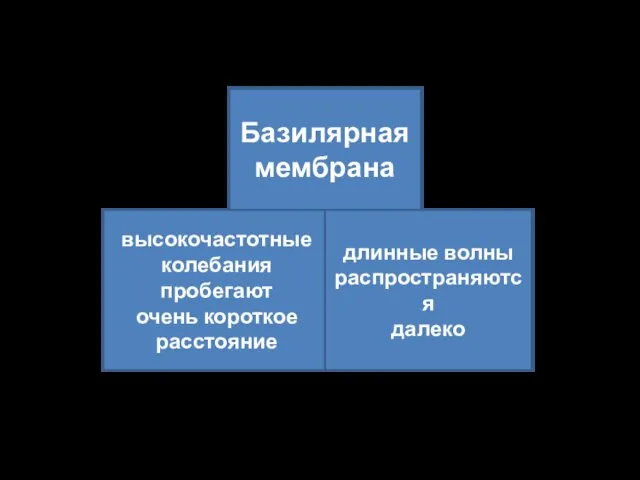
Базилярная
мембрана
высокочастотные
колебания
пробегают
очень короткое
расстояние
длинные волны
распространяются
далеко
Базилярная
мембрана
высокочастотные
колебания
пробегают
очень короткое
расстояние
длинные волны
распространяются
далеко
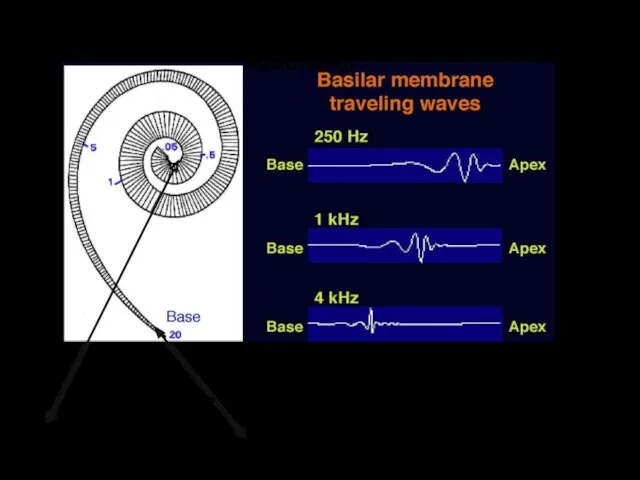
Механика базилярной мембраны
высокие частоты
низкие частоты
Принцип «места»
Механика базилярной мембраны
высокие частоты
низкие частоты
Принцип «места»

Высота звука — это субъективное восприятие частоты звуковых колебаний.
Человеческое ухо
Высота звука — это субъективное восприятие частоты звуковых колебаний.
Человеческое ухо
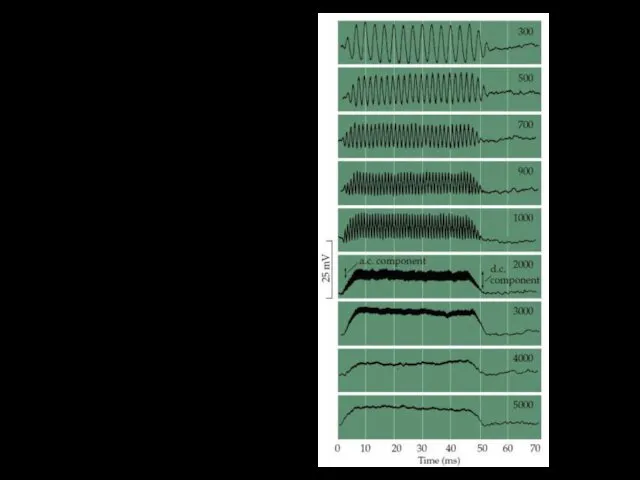
Рецепторные потенциалы волосковых клеток
Рецепторные потенциалы волосковых клеток

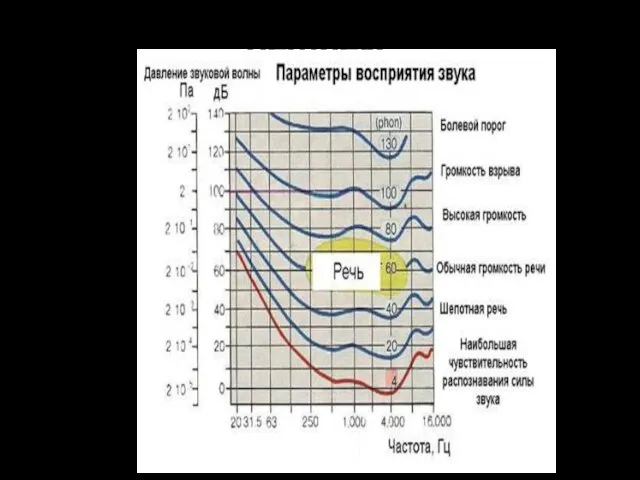
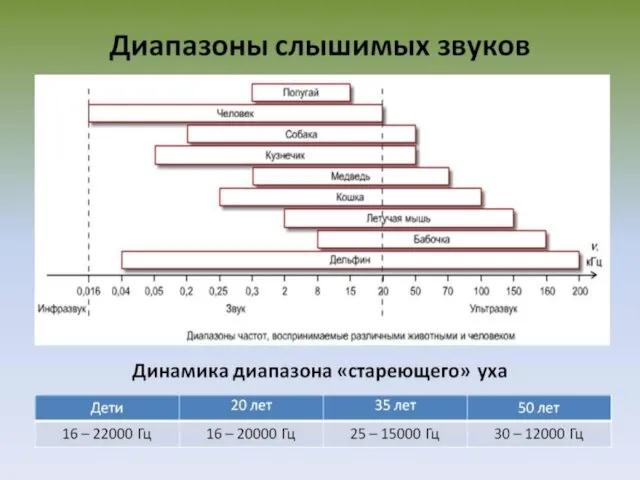
СЛУХОВЫЕ ОЩУЩЕНИЯ
Диапазон частот, который воспринимает человек, включает около 10 октав музыкальной
СЛУХОВЫЕ ОЩУЩЕНИЯ
Диапазон частот, который воспринимает человек, включает около 10 октав музыкальной

Абсолютный порог слуховой чувствительности — минимальная сила звука, которую слышит человек в
Абсолютный порог слуховой чувствительности — минимальная сила звука, которую слышит человек в
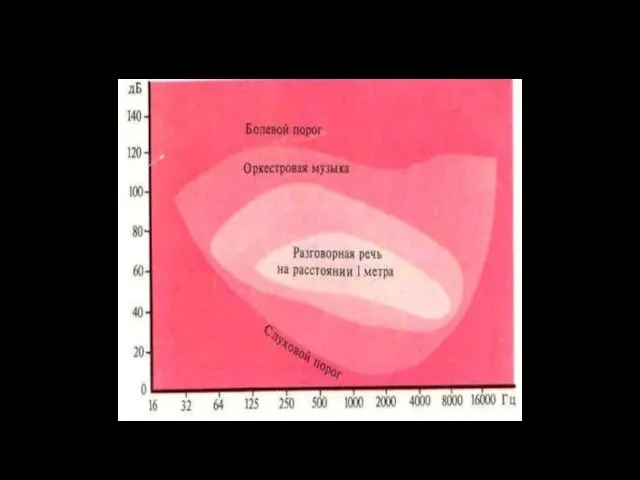

Сила звука
Диапазон силы звука, воспринимаемый человеческим ухом, огромен (болевой порог в 1013 раз
Сила звука
Диапазон силы звука, воспринимаемый человеческим ухом, огромен (болевой порог в 1013 раз

Сила звука кодируется как пространственным (изменением числа возбужденных волосковых клеток), так
Сила звука кодируется как пространственным (изменением числа возбужденных волосковых клеток), так

при раздражении волосков в ответ на звуковое колебание наружные волосковые клетки
при раздражении волосков в ответ на звуковое колебание наружные волосковые клетки

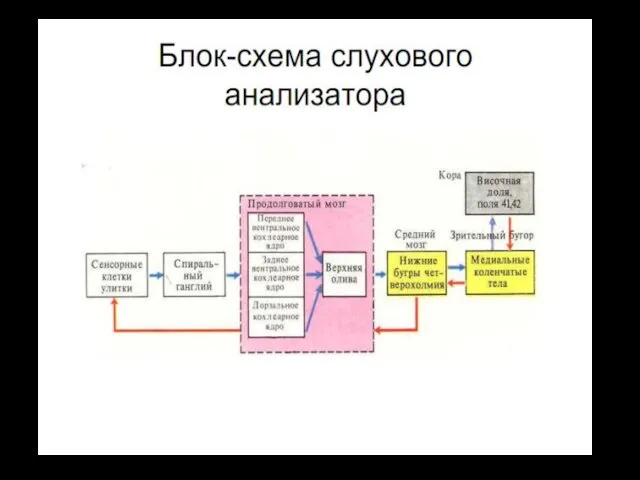
СЛУХОВЫЕ ПУТИ И ЦЕНТРЫ
Афферентные нервные волокна от улитки входят в спиральный
СЛУХОВЫЕ ПУТИ И ЦЕНТРЫ
Афферентные нервные волокна от улитки входят в спиральный

СЛУХОВЫЕ ПУТИ И ЦЕНТРЫ
От ядер верхней оливы слуховые пути поднимаются вверх
СЛУХОВЫЕ ПУТИ И ЦЕНТРЫ
От ядер верхней оливы слуховые пути поднимаются вверх

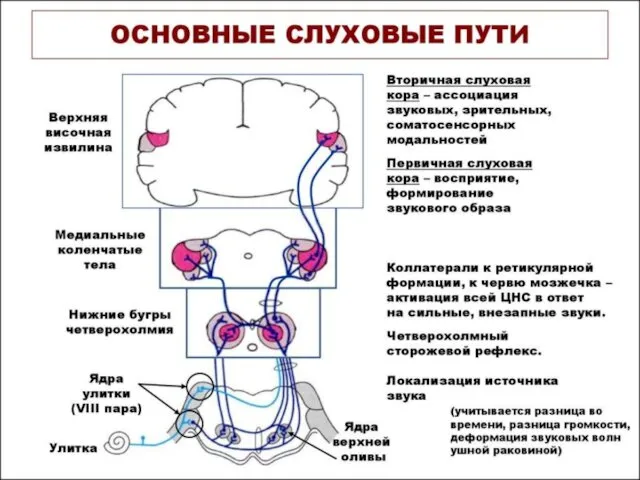
СЛУХОВЫЕ ПУТИ И ЦЕНТРЫ
Далее слуховой путь следует к медиальным коленчатым телам
СЛУХОВЫЕ ПУТИ И ЦЕНТРЫ
Далее слуховой путь следует к медиальным коленчатым телам

СЛУХОВЫЕ ПУТИ И ЦЕНТРЫ
Базилярная мембрана улитки на всех уровнях слухового пути
СЛУХОВЫЕ ПУТИ И ЦЕНТРЫ
Базилярная мембрана улитки на всех уровнях слухового пути

СЛУХОВАЯ КОРА
Первичная слуховая кора непосредственно получает сигналы от внутреннего (медиального) коленчатого
СЛУХОВАЯ КОРА
Первичная слуховая кора непосредственно получает сигналы от внутреннего (медиального) коленчатого

В каждой из 6 тонотопических карт звуки высокой частоты возбуждают нейроны

СЛУХОВАЯ КОРА
Первичная слуховая кора непосредственно получает сигналы от внутреннего (медиального) коленчатого
СЛУХОВАЯ КОРА
Первичная слуховая кора непосредственно получает сигналы от внутреннего (медиального) коленчатого

Проекционные области слуховой коры располагаются не только в верхней части верхней
Проекционные области слуховой коры располагаются не только в верхней части верхней

Тонотопические карты
Одна большая карта в первичной слуховой коре почти целиком дискриминирует
Тонотопические карты
Одна большая карта в первичной слуховой коре почти целиком дискриминирует

Тонотопические карты
Диапазон звуковой частоты, на которую отвечают нейроны слуховой коры уже,
Тонотопические карты
Диапазон звуковой частоты, на которую отвечают нейроны слуховой коры уже,

Тонотопические карты
Многие нейроны слуховой коры, особенно в слуховой ассоциативной коре, отвечают
Тонотопические карты
Многие нейроны слуховой коры, особенно в слуховой ассоциативной коре, отвечают

Бинауральный слух
Человек определяет звук, двумя путями:
1. Временем задержки между поступлением
Бинауральный слух
Человек определяет звук, двумя путями:
1. Временем задержки между поступлением

Бинауральный слух
.
2. Различием между интенсивностью звуков в двух ушах:
Бинауральный слух
.
2. Различием между интенсивностью звуков в двух ушах:
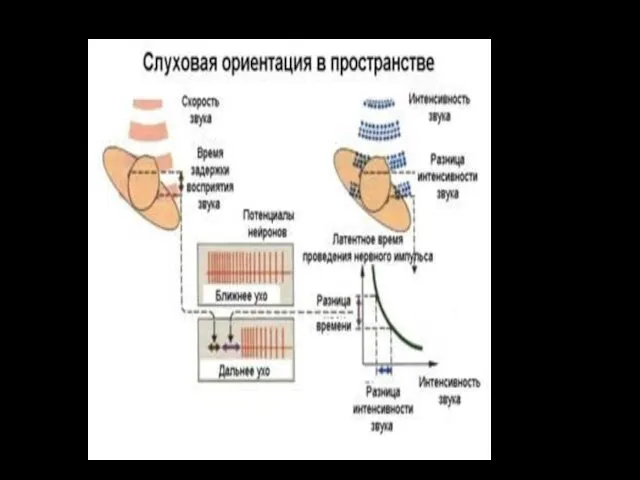

Такое тонкое сравнение минимальной разницы во времени прихода звука осуществляется ЦНС
Такое тонкое сравнение минимальной разницы во времени прихода звука осуществляется ЦНС

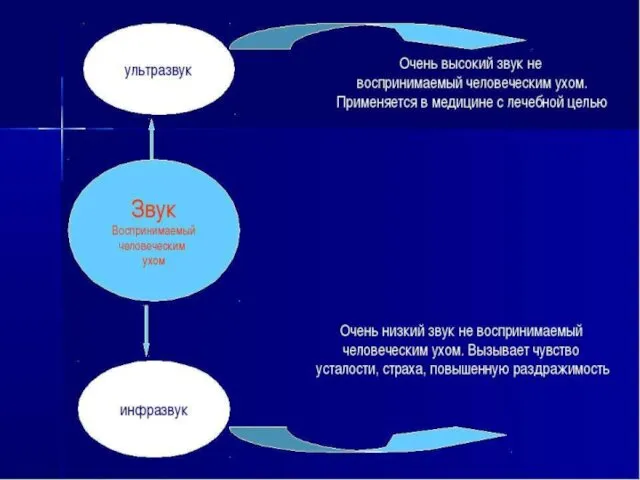

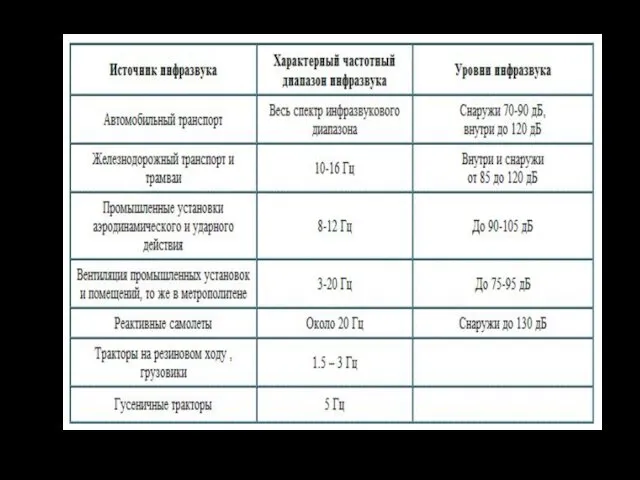


Вестибулярный анализатор
Вестибулярный анализатор

Вестибулярный анализатор
Совокупность механорецепторов и нервных структур, обеспечивающих восприятие и анализ ориентации
Вестибулярный анализатор
Совокупность механорецепторов и нервных структур, обеспечивающих восприятие и анализ ориентации
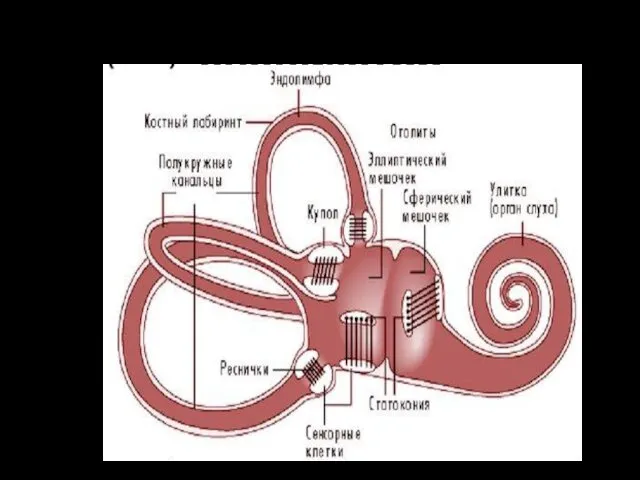
Вестибулярный анализатор
Вестибулярный анализатор
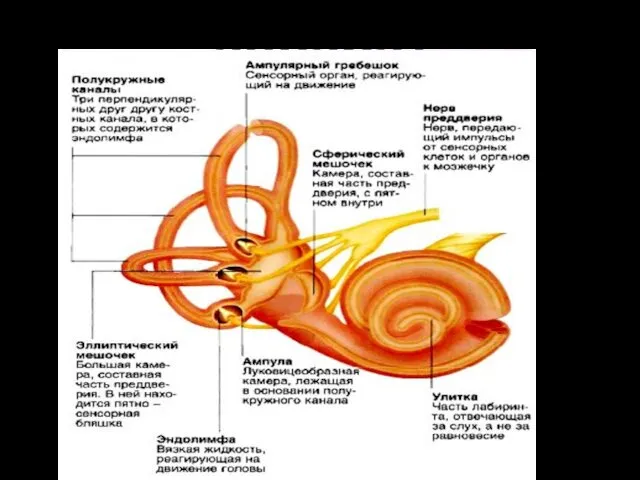
вестибулярный аппарат
вестибулярный аппарат

Рецепторный аппарат
В преддверии находится 2 мешочка – круглый (sacculus) и эллиптический
Рецепторный аппарат
В преддверии находится 2 мешочка – круглый (sacculus) и эллиптический

Рецепторный аппарат
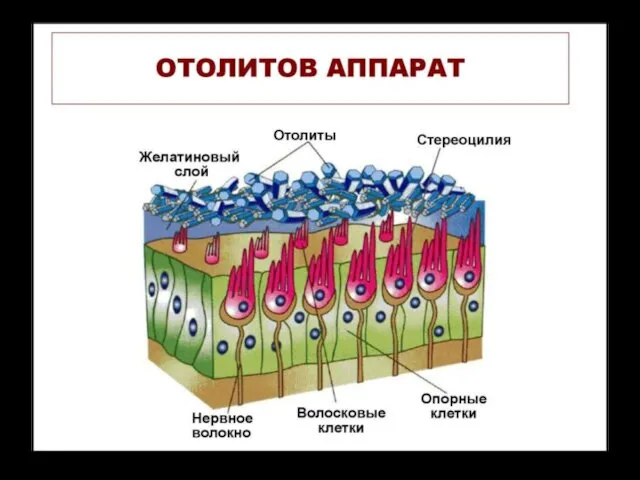
Возбуждение клеток происходит при скольжении отолитовой мембраны по волоскам, то
Рецепторный аппарат
Возбуждение клеток происходит при скольжении отолитовой мембраны по волоскам, то
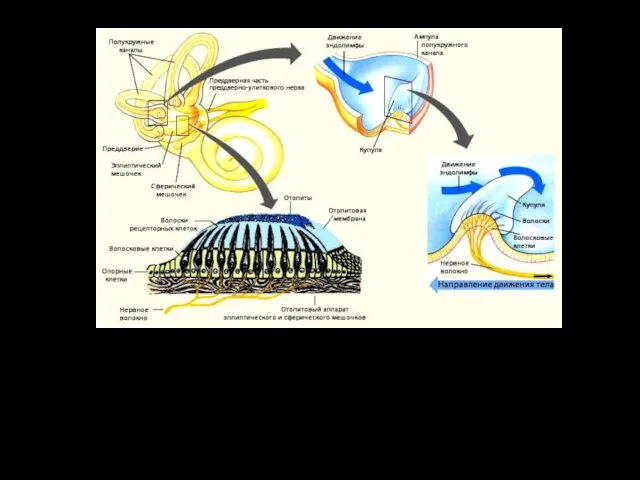
Гребешки и пятна — воспринимающие области вестибулярного органа — содержат рецепторные волосковые клетки.
Гребешки и пятна — воспринимающие области вестибулярного органа — содержат рецепторные волосковые клетки.
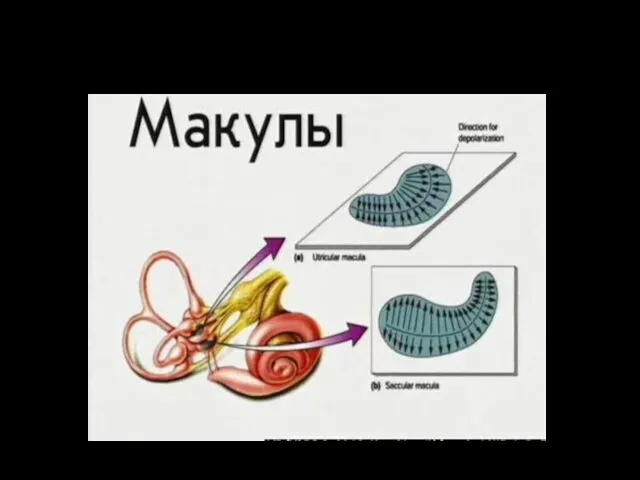
В утрикулисе и саккулисе комплексы волосковых клеток собраны в группы овальной
В утрикулисе и саккулисе комплексы волосковых клеток собраны в группы овальной

Пятно эллиптического мешочка лежит горизонтально и обеспечивает информацию при движении головы
Пятно эллиптического мешочка лежит горизонтально и обеспечивает информацию при движении головы
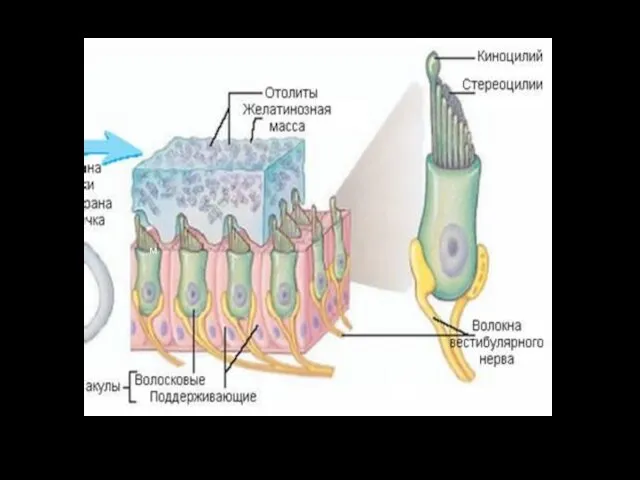
м
м

Сила инерции, которая возникает при линейных ускорениях: во время движений "вверх - вниз", "вперед-назад“ действует на эндо-лимфу и отолитовую мембрану по-разному. Более тяжёлая, и иннерционная мембрана отстает от эндолимфы в начале движения и позднее останавливается в случае торможения.
Поэтому, именно, в эти моменты и
Сила инерции, которая возникает при линейных ускорениях: во время движений "вверх - вниз", "вперед-назад“ действует на эндо-лимфу и отолитовую мембрану по-разному. Более тяжёлая, и иннерционная мембрана отстает от эндолимфы в начале движения и позднее останавливается в случае торможения.
Поэтому, именно, в эти моменты и
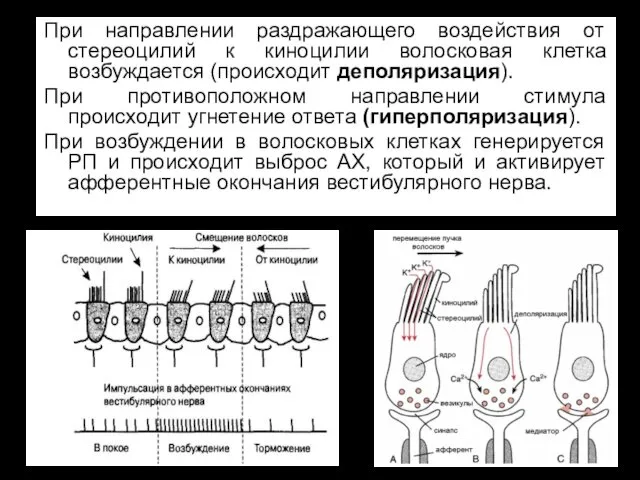
При направлении раздражающего воздействия от стереоцилий к киноцилии волосковая клетка возбуждается
При направлении раздражающего воздействия от стереоцилий к киноцилии волосковая клетка возбуждается
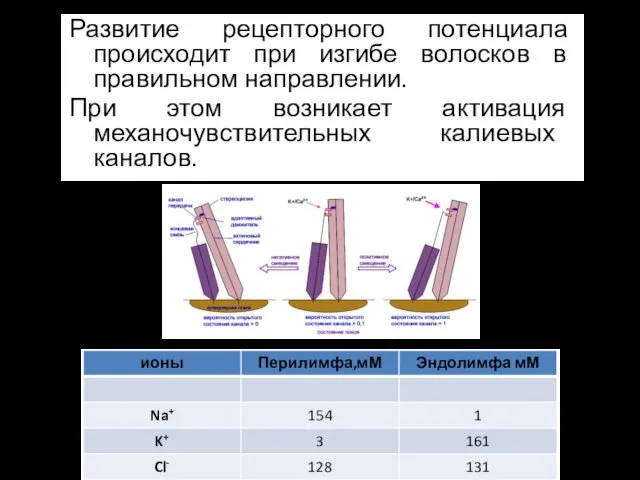
Рецепторный потенциал
Развитие рецепторного потенциала происходит при изгибе волосков в правильном направлении.
Рецепторный потенциал
Развитие рецепторного потенциала происходит при изгибе волосков в правильном направлении.
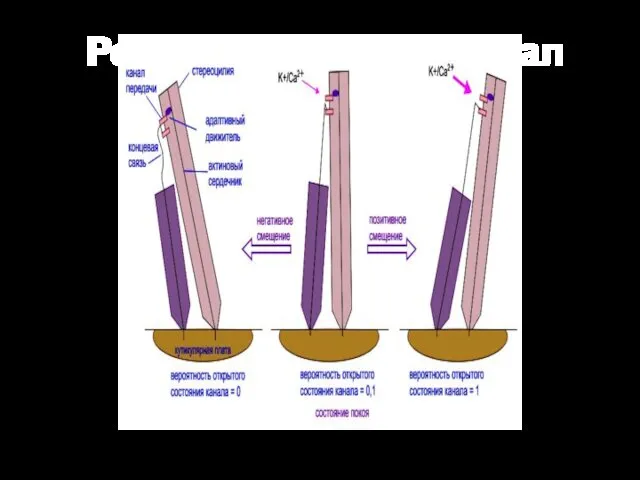
Рецепторный потенциал
Рецепторный потенциал
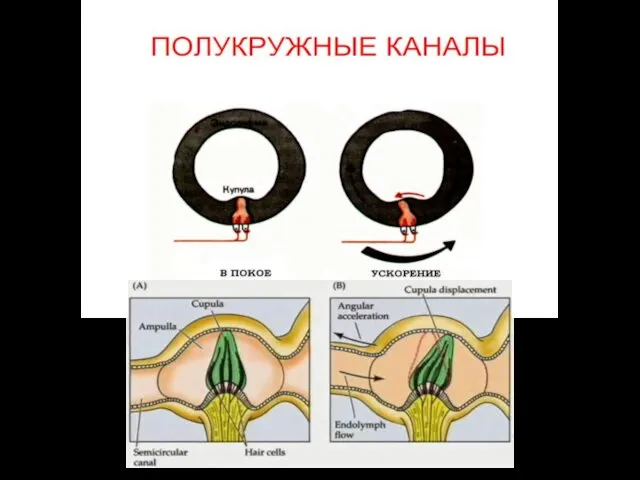

Детекция поворота головы
Полукружные каналы детектируют поворот или вращение головы.
Когда голова
Детекция поворота головы
Полукружные каналы детектируют поворот или вращение головы.
Когда голова

Реакции организма, вызванные стимуляцией полукружных каналов
Стимуляция полукружных каналов вызывает субъективные ощущения
Реакции организма, вызванные стимуляцией полукружных каналов
Стимуляция полукружных каналов вызывает субъективные ощущения

ПРОЕКЦИОННЫЕ ПУТИ ВЕСТИБУЛЯРНОГО АППАРАТА
Вестибулярная ветвь VIII нерва образована отростками примерно 19
ПРОЕКЦИОННЫЕ ПУТИ ВЕСТИБУЛЯРНОГО АППАРАТА
Вестибулярная ветвь VIII нерва образована отростками примерно 19

ПРОЕКЦИОННЫЕ ПУТИ ВЕСТИБУЛЯРНОГО АППАРАТА
Аксоны нервных клеток второго порядка связаны со спинным
ПРОЕКЦИОННЫЕ ПУТИ ВЕСТИБУЛЯРНОГО АППАРАТА
Аксоны нервных клеток второго порядка связаны со спинным

ПРОЕКЦИОННЫЕ ПУТИ ВЕСТИБУЛЯРНОГО АППАРАТА
Есть два основных пути поступления вестибулярных сигналов в
ПРОЕКЦИОННЫЕ ПУТИ ВЕСТИБУЛЯРНОГО АППАРАТА
Есть два основных пути поступления вестибулярных сигналов в


Вестибулосоматические реакции
К вестибулосоматическим реакциям относят вестибулярный нистагм - ритмические скачкообразные движения глазных яблок.
Вестибулосоматические реакции
К вестибулосоматическим реакциям относят вестибулярный нистагм - ритмические скачкообразные движения глазных яблок.
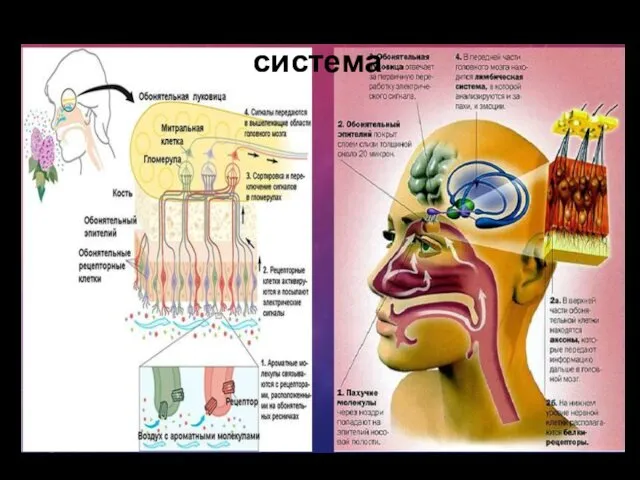
Обонятельная сенсорная система
Обонятельная сенсорная система

Классификация запахов по Эймуэру
Первичные или основные:
- камфорный - (камфора, 1,8-цинеол)
Классификация запахов по Эймуэру
Первичные или основные:
- камфорный - (камфора, 1,8-цинеол)
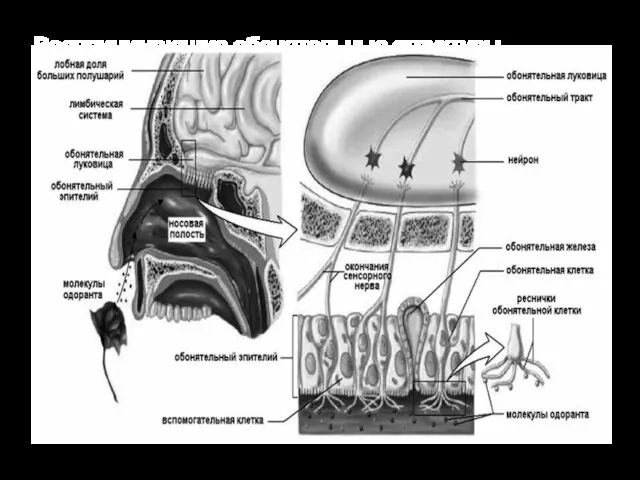
Воспринимающие обонятельные структуры образуют в слизистой оболочке носа специализированную область — парную
Воспринимающие обонятельные структуры образуют в слизистой оболочке носа специализированную область — парную

ВТЯГИВАНИЕ ВОЗДУХА
Область, содержащая обонятельные рецепторы, плохо вентилируется. Обычно воздух спокойно движется
ВТЯГИВАНИЕ ВОЗДУХА
Область, содержащая обонятельные рецепторы, плохо вентилируется. Обычно воздух спокойно движется
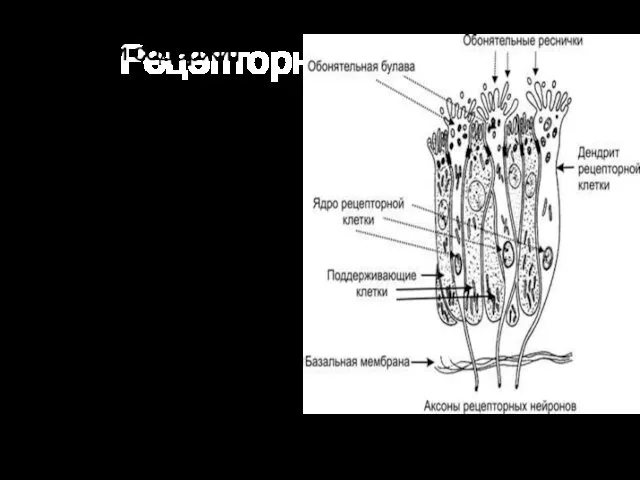
Рецепторный аппарат
Обонятельный эпителий содержит опорные эпителиальные клетки и расположенные между ними
Рецепторный аппарат
Обонятельный эпителий содержит опорные эпителиальные клетки и расположенные между ними
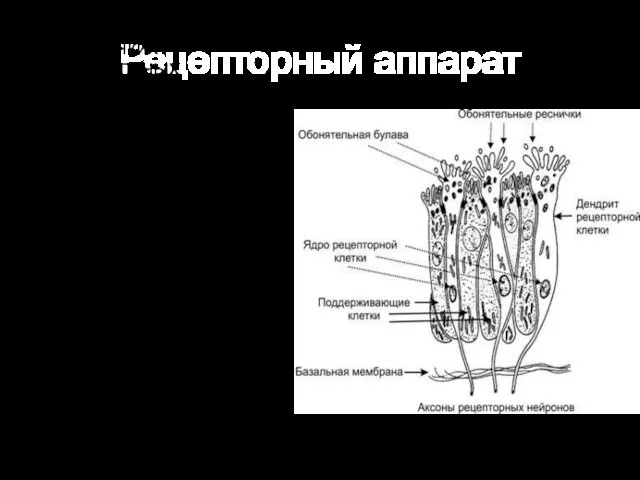
Рецепторный аппарат
От булавы отходит от 8 до 40 тонких обонятельных волосков —
Рецепторный аппарат
От булавы отходит от 8 до 40 тонких обонятельных волосков —
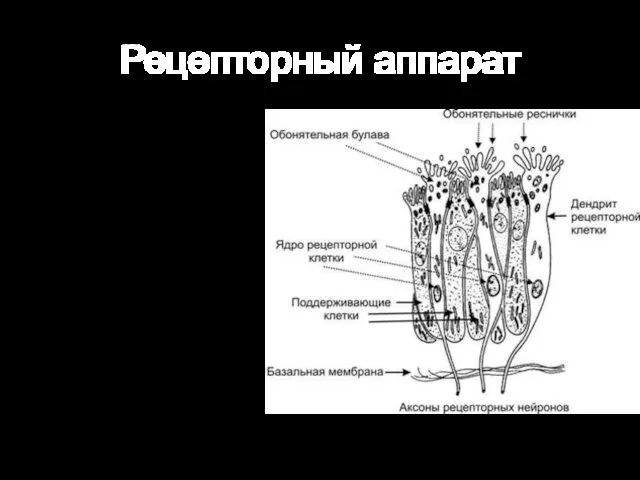
Рецепторный аппарат
Продолжительность жизни обонятельных клеток — 30–35 дней. Они относятся к обновляющейся клеточной
Рецепторный аппарат
Продолжительность жизни обонятельных клеток — 30–35 дней. Они относятся к обновляющейся клеточной

В обонятельной луковице человека различают шесть слоев, которые располагаются концентрически, считая
В обонятельной луковице человека различают шесть слоев, которые располагаются концентрически, считая

IV слой – тел митральных клеток. Это крупные нейроны (диаметр сомы
IV слой – тел митральных клеток. Это крупные нейроны (диаметр сомы
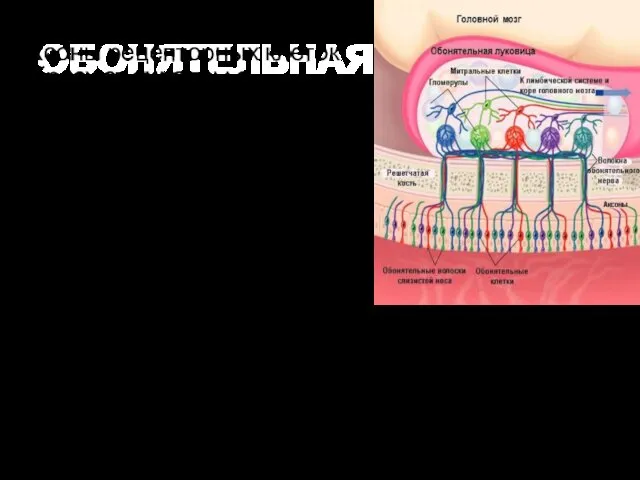
ОБОНЯТЕЛЬНАЯ ЛУКОВИЦА
В обонятельной луковице аксоны рецепторных клеток образуют синапсы с дендритами
ОБОНЯТЕЛЬНАЯ ЛУКОВИЦА
В обонятельной луковице аксоны рецепторных клеток образуют синапсы с дендритами
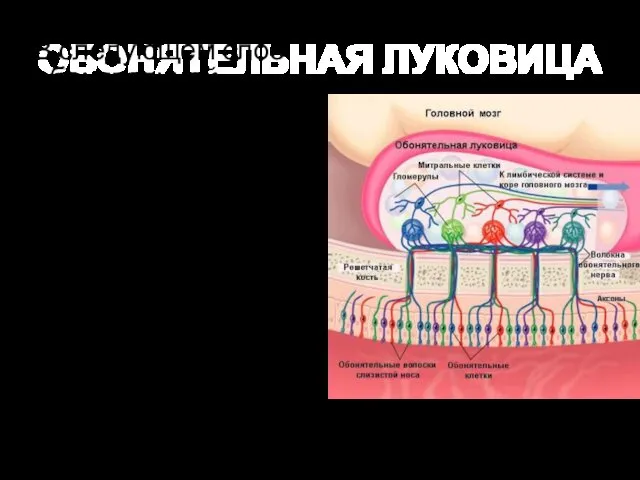
ОБОНЯТЕЛЬНАЯ ЛУКОВИЦА
В следующем слое обонятельной луковицы дендриты митральных, зернистых и пучковых
ОБОНЯТЕЛЬНАЯ ЛУКОВИЦА
В следующем слое обонятельной луковицы дендриты митральных, зернистых и пучковых
 Буква Э
Буква Э Дорожные знаки
Дорожные знаки Мультиметр (авометр). Практическая работа №1 Измерения мультиметром М832
Мультиметр (авометр). Практическая работа №1 Измерения мультиметром М832 Основные методы и средства измерения, применяемые для контроля технологического процесса котельного оборудования
Основные методы и средства измерения, применяемые для контроля технологического процесса котельного оборудования Четвериков Андрей
Четвериков Андрей Становление инноватики. Эволюционная экономика
Становление инноватики. Эволюционная экономика Opening a fast food outlet
Opening a fast food outlet Химическая промышленность
Химическая промышленность Применение информационных технологий в автомобильном транспорте
Применение информационных технологий в автомобильном транспорте Before and after
Before and after Игра Разговор о правильном питании
Игра Разговор о правильном питании Допустимые уловы (ОДУ) водных биологических ресурсов на 2020 год в Удмуртском секторе Воткинского водохранилища
Допустимые уловы (ОДУ) водных биологических ресурсов на 2020 год в Удмуртском секторе Воткинского водохранилища Покрытия промышленных зданий
Покрытия промышленных зданий Дифференциация звуков Ш-Ж
Дифференциация звуков Ш-Ж Реконструкция. Методы усиления металлических конструкций
Реконструкция. Методы усиления металлических конструкций Аминокислоты. Химический состав белков, строение белковой молекулы. Свойства и функции белков
Аминокислоты. Химический состав белков, строение белковой молекулы. Свойства и функции белков Музыкальная викторина
Музыкальная викторина Роль детского технопарка Кванториум в системе выявления и сопровождения одаренных детей
Роль детского технопарка Кванториум в системе выявления и сопровождения одаренных детей Презентация к уроку Франция и Германия
Презентация к уроку Франция и Германия Деление дробей. Устно
Деление дробей. Устно Качественные показатели каналов и трактов военных радиорелейных линий. (Тема 1.5)
Качественные показатели каналов и трактов военных радиорелейных линий. (Тема 1.5) Алгоритм создания интерактивной карты
Алгоритм создания интерактивной карты Цели и задачи испытаний РЭА
Цели и задачи испытаний РЭА Оборотные средства предприятитя
Оборотные средства предприятитя Конституция РФ – гарант свобод и прав народных
Конституция РФ – гарант свобод и прав народных 9 мая – день Победы
9 мая – день Победы 20240124_izo_7_klass._tema_11._vzaimosvyaz_obektov_v_arhitekturnom_makete
20240124_izo_7_klass._tema_11._vzaimosvyaz_obektov_v_arhitekturnom_makete Народная кукла, как средство приобщения детей дошкольного возраста к русской традиционной культуре
Народная кукла, как средство приобщения детей дошкольного возраста к русской традиционной культуре