- Цитология

Содержание
- 2. Схема строения плазмолеммы
- 3. Компоненты плазматической мембраны: A — холестерин B — олигосахарид в составе гликопротеина на наружной поверхности C
- 4. Транспорт веществ через плазмалемму Виды транспорта через мембрану: 1 – простая диффузия; 2 – диффузия через
- 5. Эндоцитоз и секреция (экзоцитоз) Опосредованный рецепторами эндоцитоз и секреция. От плазмолеммы отделяется пузырёк, окружённый клатриновой оболочкой,
- 6. Эндоцитоз (А) и экзоцитоз (Б). При эндоцитозе участок плазматической мембраны впячивается и замыкается. Инвагинация плазмолеммы и
- 7. Эндоцитоз Опосредуемый рецепторами эндоцитоз (А). Многие внеклеточные макромолекулы (трансферрин, ЛНП, вирусные частицы и др.) связываются со
- 8. Фагоцитоз Внутриклеточное переваривание. Лизосомные ферменты расщепляют поглощённый клеткой материал, а также части собственной клетки. При фагоцитозе
- 9. Комплекс Гольджи Комплекс Гольджи. Белки и липиды поступают в комплекс Гольджи с цис-компартмента. Транспортные пузырьки переносят
- 10. Схема простого межклеточного соединения а — простое соединение, без участия специальных структур; б — трансмембранные гликопротеиды
- 11. Плотный контакт Плотный контакт. (А) Сшивающие трансмембранные белки образуют непроницаемые для молекул цепочки локальных соединений между
- 12. Адгезивный поясок Адгезивный (сцепляющий) поясок (лента) а — расположение его в клетке; 6 — вид на
- 13. Фокальный контакт а — расположение в фибробласте; 6 — молекулярная схема. 1 — плазматическая мембрана; 2
- 14. Десмосома Десмосома: строение (А) и организация (Б). Плазматические мембраны клеток разделены промежутком 20–30 нм, в котором
- 15. Щелевой контакт Щелевой контакт. Шесть белковых СЕ в плазматической мембране образуют коннексон. При совмещении коннексонов смежных
- 16. Синапс
- 17. Ядро Этапы считывания генетической информации. В ходе транскрипции на ДНК–матрице синтезируется длинная молекула РНК (первичный транскрипт),
- 18. Хромосома Организация хромосомы. В неконденсированном хроматине двойная спираль ДНК лежит на поверхности октамера гистонов (H2A, H2B,
- 19. Клеточный цикл Стадии клеточного цикла. В клеточном цикле различают митоз — сравнительно короткую фазу M и
- 20. Митоз M–фаза клеточного цикла (митоз). После фазы G2 начинается M–фаза клеточного цикла. Она состоит из пяти
- 21. Мейоз Первое деление мейоза (профаза I, метафаза I, анафаза I и телофаза I) — редукционное. Профаза
- 22. Апоптоз и некроз Морфологическая картина апоптоза и некроза клеток. При апоптозе клетка сморщивается, хроматин конденсируется, ядро
- 24. Скачать презентацию
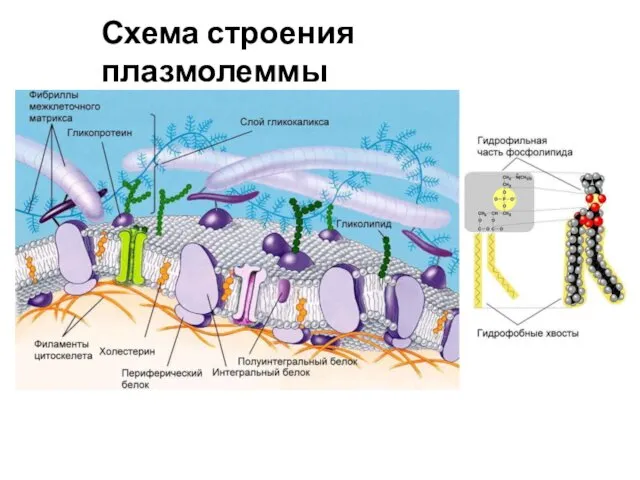
Схема строения плазмолеммы
Схема строения плазмолеммы
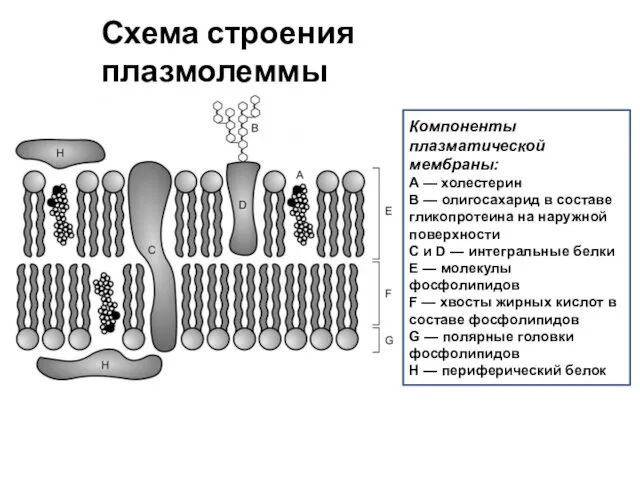
Компоненты плазматической мембраны:
A — холестерин
B — олигосахарид в составе гликопротеина на наружной поверхности
C
Компоненты плазматической мембраны:
A — холестерин
B — олигосахарид в составе гликопротеина на наружной поверхности
C
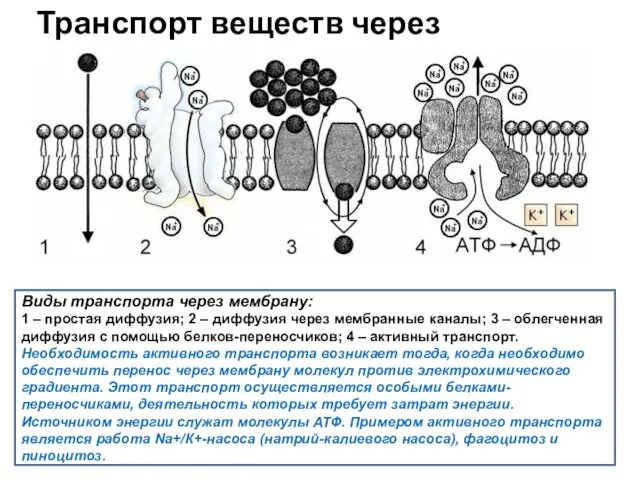
Транспорт веществ через плазмалемму
Виды транспорта через мембрану:
1 – простая диффузия; 2
Транспорт веществ через плазмалемму
Виды транспорта через мембрану:
1 – простая диффузия; 2
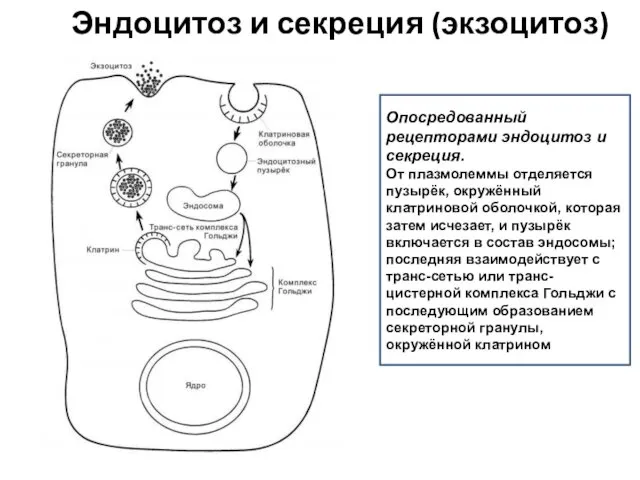
Эндоцитоз и секреция (экзоцитоз)
Опосредованный рецепторами эндоцитоз и секреция.
От плазмолеммы отделяется пузырёк,
Эндоцитоз и секреция (экзоцитоз)
Опосредованный рецепторами эндоцитоз и секреция.
От плазмолеммы отделяется пузырёк,
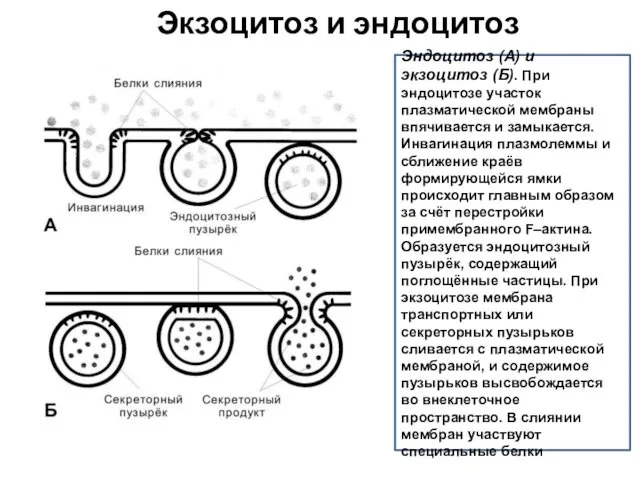
Эндоцитоз (А) и экзоцитоз (Б). При эндоцитозе участок плазматической мембраны впячивается
Эндоцитоз (А) и экзоцитоз (Б). При эндоцитозе участок плазматической мембраны впячивается
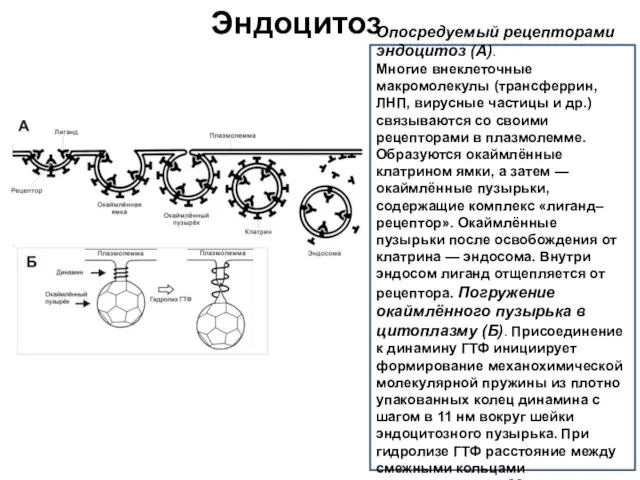
Эндоцитоз
Опосредуемый рецепторами эндоцитоз (А).
Многие внеклеточные макромолекулы (трансферрин, ЛНП, вирусные частицы
Эндоцитоз
Опосредуемый рецепторами эндоцитоз (А).
Многие внеклеточные макромолекулы (трансферрин, ЛНП, вирусные частицы
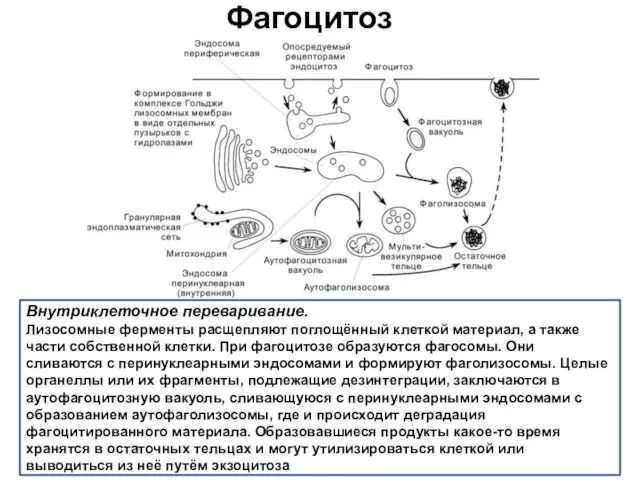
Фагоцитоз
Внутриклеточное переваривание.
Лизосомные ферменты расщепляют поглощённый клеткой материал, а также части
Фагоцитоз
Внутриклеточное переваривание.
Лизосомные ферменты расщепляют поглощённый клеткой материал, а также части
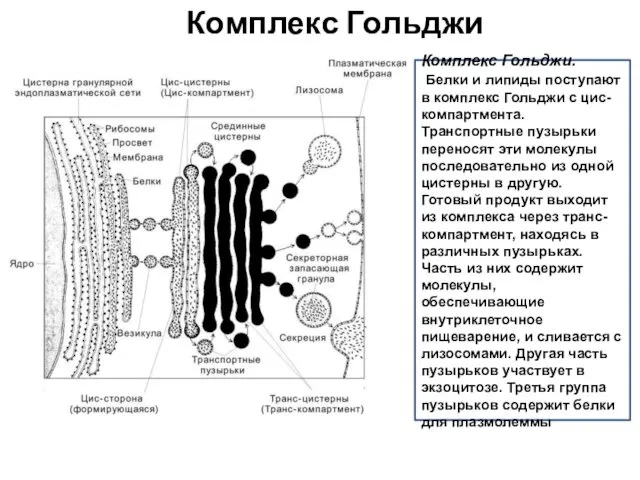
Комплекс Гольджи
Комплекс Гольджи.
Белки и липиды поступают в комплекс Гольджи с
Комплекс Гольджи
Комплекс Гольджи.
Белки и липиды поступают в комплекс Гольджи с
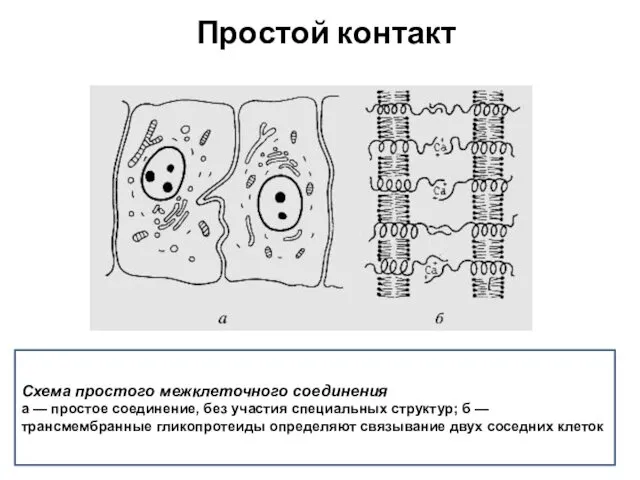
Схема простого межклеточного соединения
а — простое соединение, без участия специальных структур;
Схема простого межклеточного соединения
а — простое соединение, без участия специальных структур;
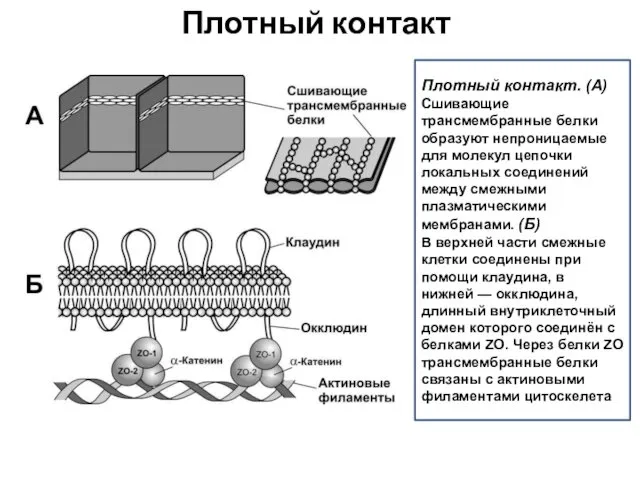
Плотный контакт
Плотный контакт. (А)
Сшивающие трансмембранные белки образуют непроницаемые для молекул цепочки
Плотный контакт
Плотный контакт. (А)
Сшивающие трансмембранные белки образуют непроницаемые для молекул цепочки
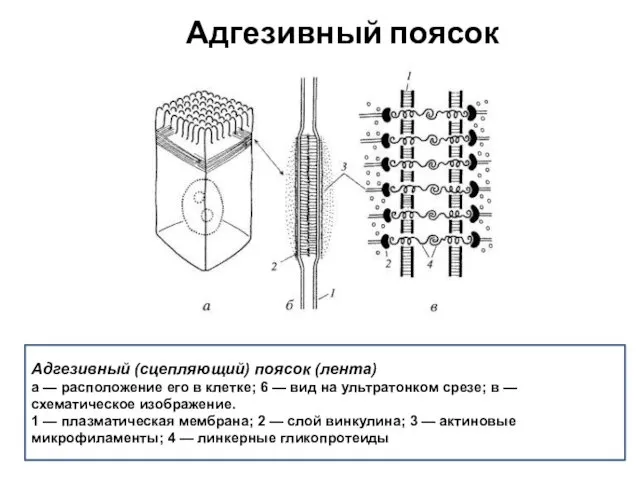
Адгезивный поясок
Адгезивный (сцепляющий) поясок (лента)
а — расположение его в клетке; 6
Адгезивный поясок
Адгезивный (сцепляющий) поясок (лента)
а — расположение его в клетке; 6
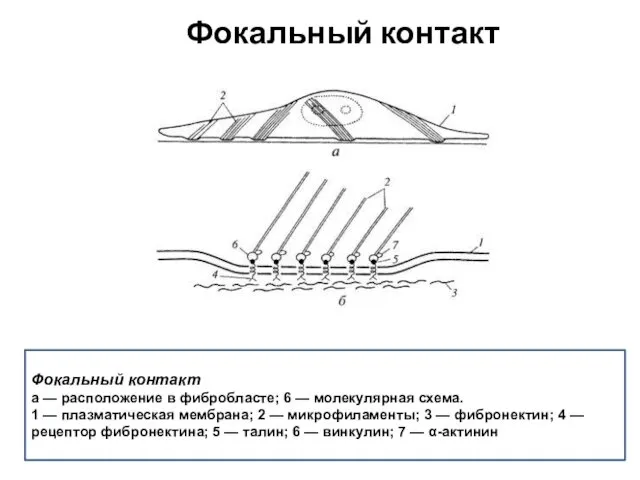
Фокальный контакт
а — расположение в фибробласте; 6 — молекулярная схема.
1 —
Фокальный контакт
а — расположение в фибробласте; 6 — молекулярная схема.
1 —
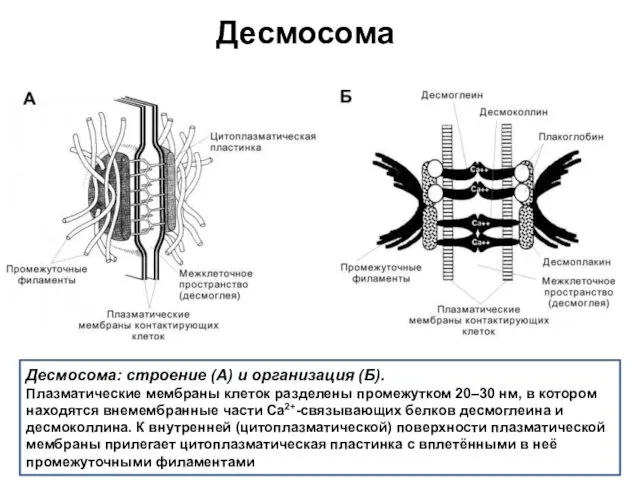
Десмосома
Десмосома: строение (А) и организация (Б).
Плазматические мембраны клеток разделены промежутком
Десмосома
Десмосома: строение (А) и организация (Б).
Плазматические мембраны клеток разделены промежутком
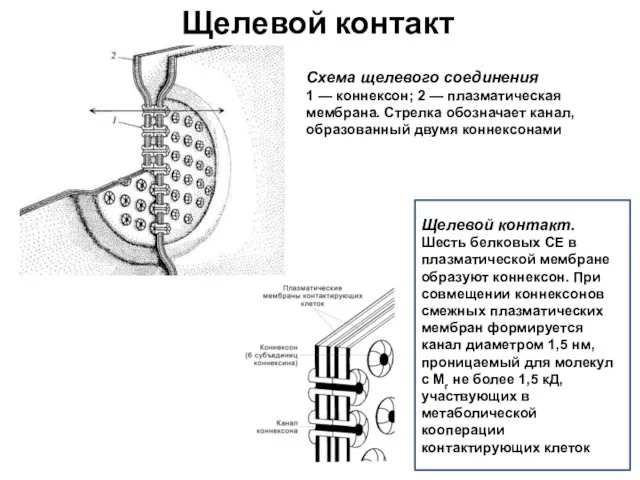
Щелевой контакт
Щелевой контакт.
Шесть белковых СЕ в плазматической мембране образуют коннексон.
Щелевой контакт
Щелевой контакт.
Шесть белковых СЕ в плазматической мембране образуют коннексон.
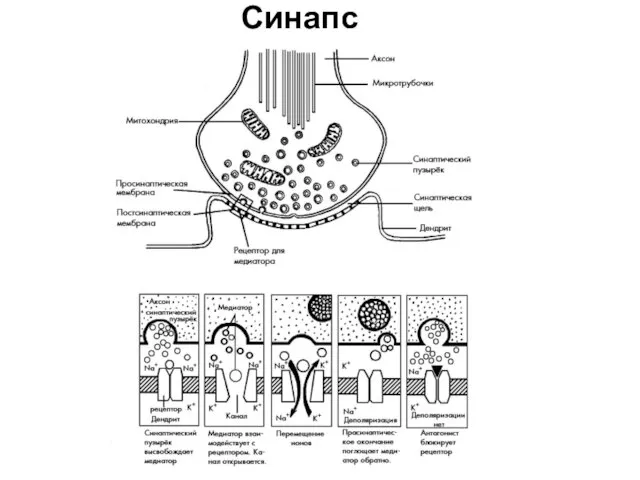
Синапс
Синапс
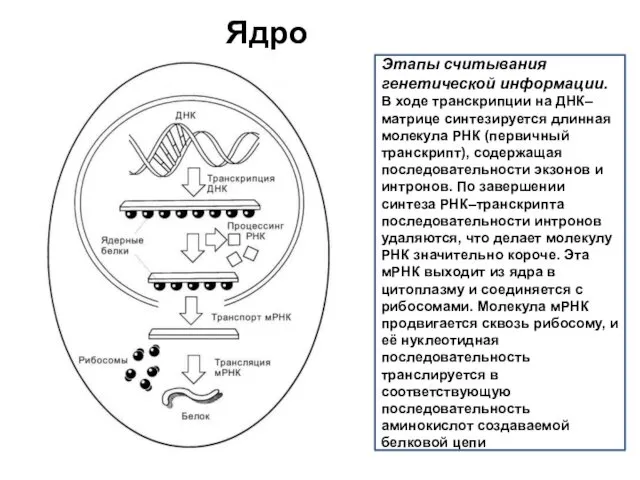
Ядро
Этапы считывания генетической информации.
В ходе транскрипции на ДНК–матрице синтезируется длинная
Ядро
Этапы считывания генетической информации.
В ходе транскрипции на ДНК–матрице синтезируется длинная
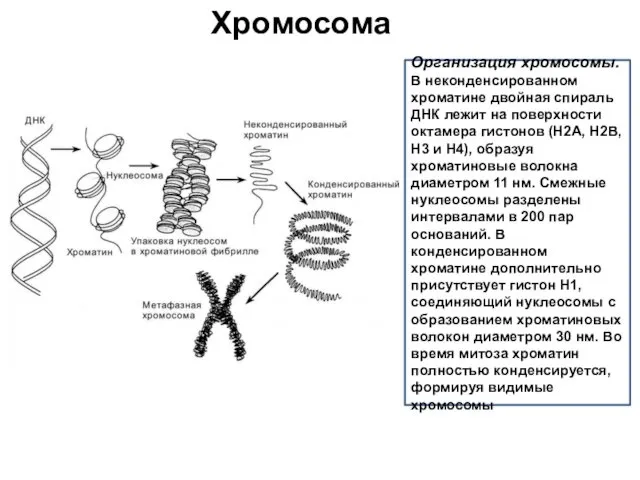
Хромосома
Организация хромосомы.
В неконденсированном хроматине двойная спираль ДНК лежит на поверхности
Хромосома
Организация хромосомы.
В неконденсированном хроматине двойная спираль ДНК лежит на поверхности
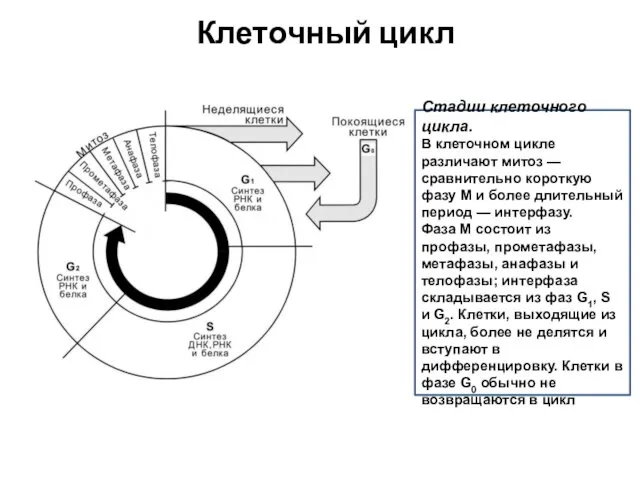
Клеточный цикл
Стадии клеточного цикла.
В клеточном цикле различают митоз — сравнительно короткую
Клеточный цикл
Стадии клеточного цикла.
В клеточном цикле различают митоз — сравнительно короткую
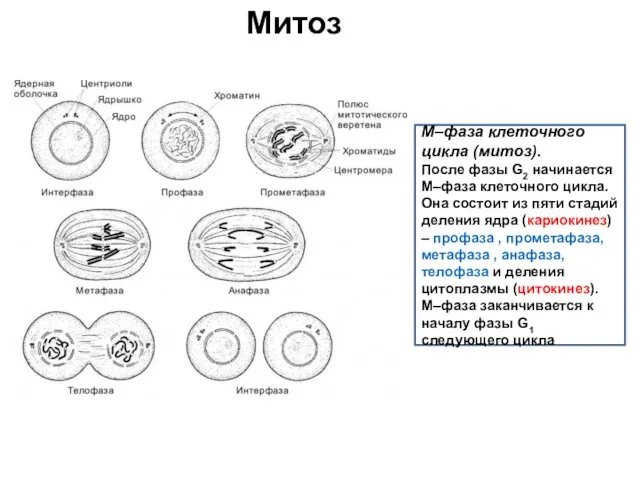
Митоз
M–фаза клеточного цикла (митоз).
После фазы G2 начинается M–фаза клеточного цикла.
Митоз
M–фаза клеточного цикла (митоз).
После фазы G2 начинается M–фаза клеточного цикла.
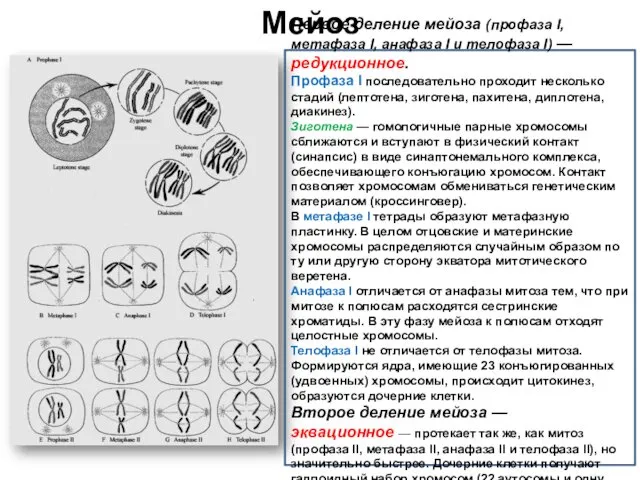
Мейоз
Первое деление мейоза (профаза I, метафаза I, анафаза I и телофаза I) — редукционное.
Профаза I последовательно проходит
Мейоз
Первое деление мейоза (профаза I, метафаза I, анафаза I и телофаза I) — редукционное.
Профаза I последовательно проходит
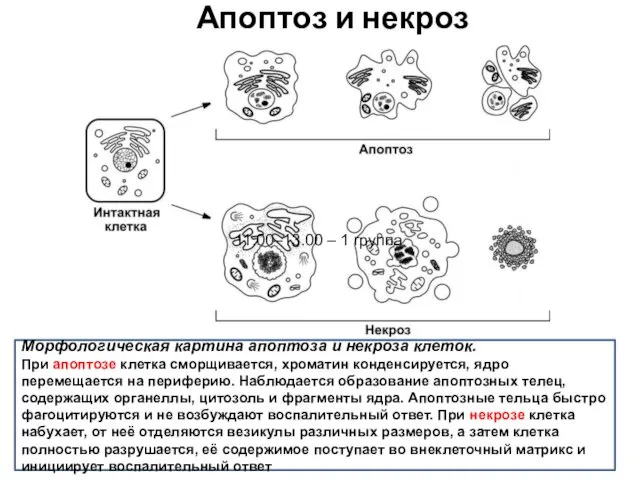
Апоптоз и некроз
Морфологическая картина апоптоза и некроза клеток.
При апоптозе клетка
Апоптоз и некроз
Морфологическая картина апоптоза и некроза клеток.
При апоптозе клетка
 Дефектация и сортировка деталей
Дефектация и сортировка деталей Деловая карьера
Деловая карьера Фаддей Беллинсгаузен
Фаддей Беллинсгаузен Рождество Христово
Рождество Христово Дорожная одежда. Искусственные сооружения на автомобильных дорогах
Дорожная одежда. Искусственные сооружения на автомобильных дорогах Чем что делают
Чем что делают Геологические процессы в недрах Земли
Геологические процессы в недрах Земли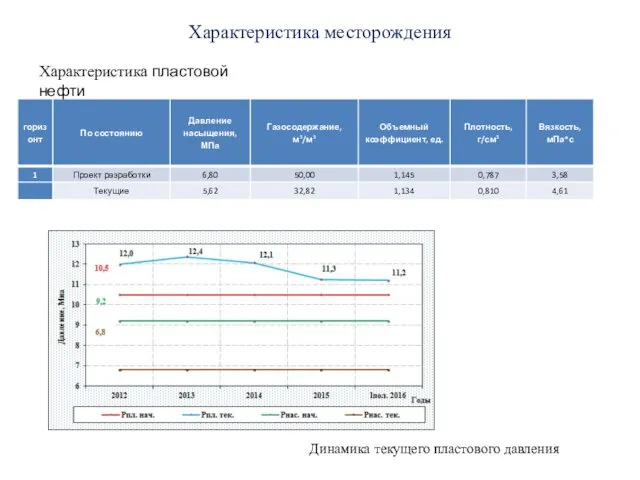 Характеристика месторождения
Характеристика месторождения Игра морской бой Птичье царство
Игра морской бой Птичье царство Игра Вперед, заток! (5-7 классы)
Игра Вперед, заток! (5-7 классы) Международное сотрудничество в области поддержания стабильности и мира
Международное сотрудничество в области поддержания стабильности и мира Ветрогенераторы: классификация и типы, конструкция и схема работы
Ветрогенераторы: классификация и типы, конструкция и схема работы В. Драгунский Всё тайное становится явным
В. Драгунский Всё тайное становится явным физра
физра Стандартные образцы в цветной металлургии
Стандартные образцы в цветной металлургии Нервная ткань
Нервная ткань Подготовка к ГИА и ЕГЭ
Подготовка к ГИА и ЕГЭ Психологические аспекты профессионального обучения и личности педагога
Психологические аспекты профессионального обучения и личности педагога Стекло и его свойства
Стекло и его свойства Что мы хотим сделать…работа с ботом
Что мы хотим сделать…работа с ботом 20231003_prezentatsiya_po_teme_muzyka_i_kino
20231003_prezentatsiya_po_teme_muzyka_i_kino Motor starting
Motor starting Мышцы. Мышечная система
Мышцы. Мышечная система Празднование Рождества в Германии
Празднование Рождества в Германии Виды стоматологических имплантов
Виды стоматологических имплантов Патофизиология углеводного обмена. Сахарный диабет
Патофизиология углеводного обмена. Сахарный диабет Портфолио Чубыкиной Ирины Юрьевны воспитателя МБДОУ Детский сад №18 Колобок г. Брянск (часть IV)
Портфолио Чубыкиной Ирины Юрьевны воспитателя МБДОУ Детский сад №18 Колобок г. Брянск (часть IV) Компетентностный контекст современного образования
Компетентностный контекст современного образования