Влияние температуры на рост и фотосинтез водорослей. Зависимость фотосинтеза от обеспеченности биогенными элементами презентация
- Влияние температуры на рост и фотосинтез водорослей. Зависимость фотосинтеза от обеспеченности биогенными элементами

Содержание
- 2. Сегодня: Влияние температуры на рост и фотосинтез водорослей Зависимость фотосинтеза от обеспеченности биогенными элементами Для ответа
- 3. Влияние температуры на рост и фотосинтез водорослей
- 4. Относительная скорость роста (1/сут) Диапазон оптимальной температуры Температура максимальной скорости роста Влияние температуры на рост водорослей
- 5. Данные экспериментов при насыщающей освещенности и обеспеченности биогенными элементами Относительная скорость роста (1/сут), усреднение для классов
- 6. Скорость фотохимических реакций от температуры НЕ зависит Улавливание фотонов, передача энергии возбуждения на РЦ ФС1 и
- 7. Белковые комплексы, осуществляющие транспорт электронов, встроены в тилакоидные мембраны - скорость реакций в них зависит от
- 8. На светолимитированном участке фотосинтез определяет улавливание световой энергии и первичное разделение зарядов. Кратковременное изменение То незначительно
- 9. Освещенность Квантовая эффективность фотосинтеза 25 оС 15 оС На светонасыщающем участке c уменьшением температуры снижается квантовый
- 10. α Рmax Ен Хл/C Т, Со Изменения параметров фотосинтеза при изменении температуры Скорость фотосинтетической фиксации углерода
- 11. Следует отметить, что возрастание интенсивности дыхания идет медленнее, фотосинтеза – важно для оценки чистой первичной продукции,
- 12. Увеличение клеточных потребностей в азоте, и если его НЕ хватает – усилению азотного лимитирования Другие аспекты
- 13. ЗАВИСИМОСТЬ ФОТОСИНТЕЗА ОТ ОБЕСПЕЧЕННОСТИ БИОГЕННЫМИ ЭЛЕМЕНТАМИ Элементный состав взвешенного органического вещества в океане в среднем соответствует
- 14. Исходя из соотношения Редфилда: Значения N:P, близкие к соотношению Редфилда, свидетельствуют об оптимальных условиях обеспеченности фитопланктона
- 15. Культуры выращивали в хемостате при оптимальных условиях роста. Соответственно, соотношение N:P отражает соотношение структурных компонентов клеток
- 16. АЗОТ Формы азота в водных экосистемах: молекулярный азот (наибольшее количество), нитраты, нитриты, аммоний и органический азот.
- 17. Ассимиляция нитратов I. Активный транспорт нитратов в клетку (процесс с использованием энергии). II. Восстановление нитратов до
- 18. NO3- +8e- +10H+ → NН4+ + 3H2O фотосинтетически генерированные IV. Включение аммония в аминокислоты (в хлоропластах):
- 19. Нитратредуктаза – фермент, кодируемый ядерным геномом. Регуляция экспрессии на уровне транскрипции. Предположительно, сигнал экспрессии – восстановительно-окислительный
- 20. При освещенности ниже насыщающей (светолимитированный участок Р-Е кривой) в зависимости от того, нитраты или аммоний используются
- 21. Скорость ассимиляции минерального азота зависит от количества углеродных скелетов, восстановителя и АТФ, которые поставляются как фотосинтетическим
- 22. Популяционный уровень: снижение скорости роста, при увеличении степени клеточного дефицита азота смертность превышает скорость роста –
- 23. ВЫХОД ИЗ СОСТОЯНИЯ АЗОТНОГО ЛИМИТИРОВАНИЯ. Изменение фотосинтеза у водорослей, испытывавших дефицит азота, при поступлении в среду
- 24. Причины: 1. Конкуренция между процессами ассимиляции нитратов (нитритов и аммония) и цикла Кальвина за восстановитель и
- 25. II стадия (более суток). Скорость фотосинтетической фиксации углерода возрастает: Световые реакции: (а) возрастание синтеза белков РЦ
- 26. Концентрация Nорг в фотическом слое 8,6 – 31,4 мкмоль азота. Лабильная фракция Nорг : мочевина и
- 27. ПОТРЕБЛЕНИЕ МОЧЕВИНЫ Потребление мочевины идет за счет активного транспорта при использовании фотосинтетически генерированной АТФ (у некоторых
- 28. ПОТРЕБЛЕНИЕ АМИНОКИСЛОТ Потребление аминокислот - активный транспорт. Предположительно: наличие трех транспортных систем – для кислых, нейтральных
- 29. Потребление аминокислот зависит от вида водорослей и обеспеченности Nмин:
- 30. Относительный вклад разных субстратов (%) в суммарное количество потребляемых азот содержащих субстратов (на примере Балтийского моря)
- 31. Относительный вклад разных субстратов (%) в суммарное количество потребляемых азот содержащих субстратов аминокислоты мочевина
- 32. Лимитирование фотосинтеза недостатком фосфора: Ограничение синтеза нуклеиновых кислот. Проявляется на уровне репликации генома или на уровне
- 33. Концентрация Pорг в поверхностных морских водах может достигать 1 мкмоль и в отдельные периоды вегетационного сезона
- 34. Два типа фосфатаз: (1) конститутивные фосфатазы, активность которых не зависит от наличия и количества Pмин во
- 35. Способность к экспрессии щелочных фосфатаз выявлена у пикопланктонных цианобактерий Prochlorococcus и Synechococcus. Эти водоросли преобладают в
- 36. Лимитирование недостатком серы ведет к снижению скорости синтеза фотосинтетических пигментов и активности фотосинтетических ферментов и, соответственно,
- 37. Лимитирование недостатком железа: разобщение антенны ФС2 с РЦ ФС2: накопление неактивных РЦ ФС2 в результате снижения
- 38. Распределение пыли (г/м2 в год), приносимой ветрами из Сахары, пустынь Аравийского полуострова и Центральной Азии. С
- 39. Буря в северо-западной Африке. Облака пыли из района Сахары сносятся ветром на запад, в Атлантический океан.
- 40. Фиксация молекулярного азота Чтобы разорвать прочную тройную связь в молекуле N2, нужны либо высокие давление и
- 41. Комплекс состоит из двух ферментов: собственно нитрогеназы (называемой также Mo-Fe-белком или динитрогеназой) и дегидрогеназы (ди)нитрогеназы (Fe-белок).
- 42. Новые данные (на основе молекулярно-генетических методов): Процесс азотфиксации «стартовал» по крайней мере 3–3,5 млрд лет назад,
- 43. Гетероцисты – результат клеточной дифференциации. Дезактивация ФС2: хл-белковые комплексы и компоненты ФС2 разрушаются (активность ФС1 сохраняется);
- 44. Cyanothece 51142 Временное разделение. У ряда цианобактерий, не образующих гетероцисты, фотосинтез протекает в присутствии света (днем),
- 45. Диаметр клетки 2.5-6 мкм. Вегетирует в теплых водах (> 24 оC). В олиготрофных водах тропической зоны
- 46. Trichodesmium spp. Цианобактерии Trichodesmium фиксируют азот ТОЛЬКО на свету и при этом у них НЕТ гетероцист
- 47. Trichodesmium tenue Trichodesmium sp. Trichodesmium contortum Trichodesmium pelagicum До 80 % клеточного объема могут занимать газовые
- 48. Как же осуществляется защита нитрогеназ от фотосинтетически выделяемого кислорода у Trichodesmium? Как экспрессия Nif генов, так
- 49. Разделение азотфиксации и фотосинтеза в пространстве у Trichodesmium Окраска специфическим красителем Nif белков Фиксация азота идет
- 50. Разделение азотфиксации и фотосинтеза во времени у Trichodesmium Суточный ритм регулирует транскрипцию nif генов, синтез de
- 51. Способность к фиксации СО2 также снижается в диазоцитах Trichodesmium во время полуденной депрессии, при этом возрастает
- 52. Механизмы предотвращения блокирования нитрогеназ фотосинтетически выделяемым кислородом, ПРЕДПОЛОЖИТЕЛЬНО, также включают: тетразол Формирование в диазоцитах восстановленных зон
- 53. Гипотетический сценарий эволюционного образования диазоцитов (предложен на основе результатов расшифровки геномов цианобактерий) Диазоциты Диазоциты НЕ являются
- 54. Дальнейшая ассимиляция глутамата, образованного при азотфиксации, происходит в глутаминсинтетазо - глутамин оксоглутарат аминотрансферазном (ГС-ГОГАТ) метаболитном цикле
- 55. Для потребления фосфатов колонии Trichodesmium мигрируют – опускаются ниже пикноклина (зарегистрированы вертикальные миграции до глубины 100
- 56. Дополнительные механизмы покрытия потребностей в фосфоре при фосфорном лимитировании у Trichodesmium Мембраны - замена фосфолипидов на
- 57. Обеспечение потребностей в железе у Trichodesmium В клетках Trichodesmium высокое содержание Fe. Учитывая, что массовое развитие
- 58. Лимитирование недостатком железа ведет к снижению числа диазоцитов и скорости азофиксации А вот фотосинтетическая активность продолжает
- 59. Экосистемная роль Trichodesmium Пределы температурной толерантности для роста и азотфиксации у Trichodesmium: 20 – 34 оС,
- 60. Цветение Trichodesmium. Большой барьерный риф побережья Австралии. Снимок NASA Цветение Trichodesmium. Красное море Снимок NASA Массовое
- 61. Для цветений Trichodesmium характерно как прижизненное выделение значительно количества органического вещества, так и при отмирании клеток
- 62. Это довольно НЕОЖИДАННЫЙ результат, поскольку Trichodesmium синтезирует токсины, вызывающие гибель копепод, таких как Acartia tonsa, а
- 63. Прогнозы динамики Trichodesmium при наблюдаемом климатическом тренде Есть все основания полагать, что повышение температуры и концентрации
- 64. КОЛИМИТИРОВАНИЕ Длительное время основной парадигмой в экологии фитопланктона было утверждение, что только один ресурс лимитирует рост
- 65. 2. В так называемых «районах высокой концентрации биогенных элементов и низкой концентрации хлорофилла» фотосинтетическую активность и
- 66. 3. В ряде районов Мирового океана (Северная Атлантика, Средиземное море, Бискайский залив, норвежские фиорды, Белое море)
- 67. Мультиресурсное ко-лимитирование. Два ресурса в такой низкой концентрации (ниже порогового уровня), что их потребление уже невозможно.
- 68. Колимитирование на уровне сообщества. Добавка ресурса А стимулирует рост одной популяции, добавка ресурса В стимулирует другую.
- 69. Схема колимитирования
- 70. На следующей лекции: Пространственно-временная изменчивость первичной продукции в Мировом океане Вопросы?
- 72. Скачать презентацию

Сегодня:
Влияние температуры на рост и фотосинтез водорослей
Зависимость фотосинтеза от обеспеченности биогенными
Сегодня:
Влияние температуры на рост и фотосинтез водорослей
Зависимость фотосинтеза от обеспеченности биогенными

Влияние температуры на рост
и фотосинтез водорослей
Влияние температуры на рост
и фотосинтез водорослей
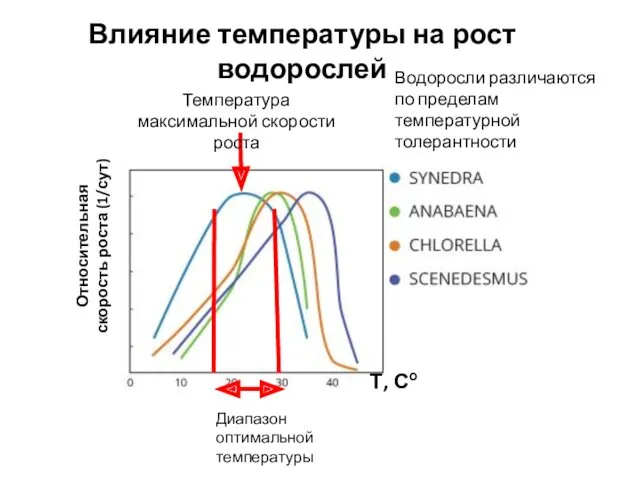
Относительная скорость роста (1/сут)
Диапазон оптимальной температуры
Температура максимальной скорости роста
Влияние температуры на
Относительная скорость роста (1/сут)
Диапазон оптимальной температуры
Температура максимальной скорости роста
Влияние температуры на
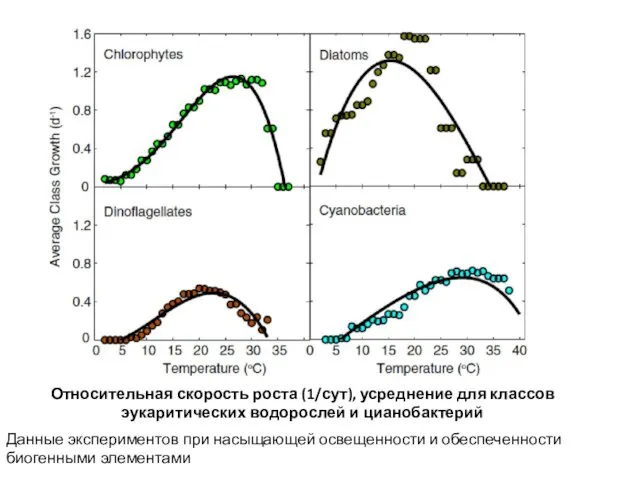
Данные экспериментов при насыщающей освещенности и обеспеченности биогенными элементами
Относительная скорость роста
Данные экспериментов при насыщающей освещенности и обеспеченности биогенными элементами
Относительная скорость роста
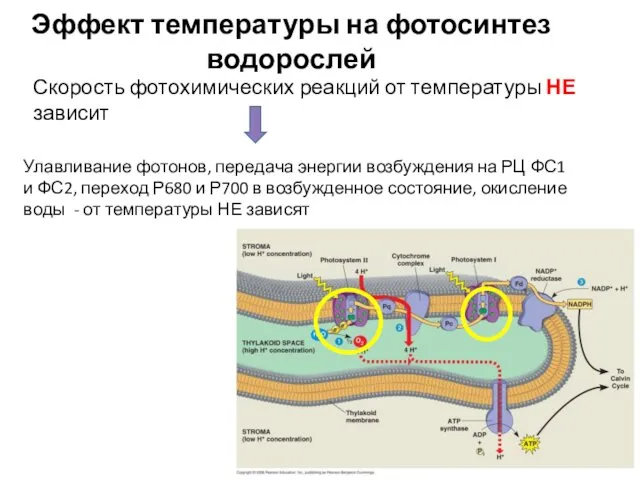
Скорость фотохимических реакций от температуры НЕ зависит
Улавливание фотонов, передача энергии возбуждения
Скорость фотохимических реакций от температуры НЕ зависит
Улавливание фотонов, передача энергии возбуждения
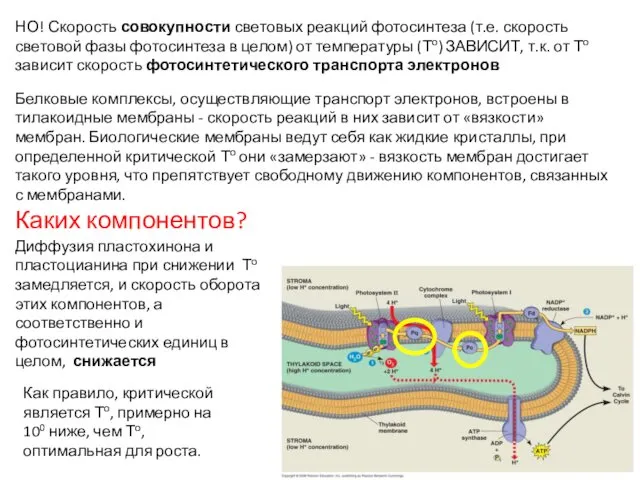
Белковые комплексы, осуществляющие транспорт электронов, встроены в тилакоидные мембраны - скорость
Белковые комплексы, осуществляющие транспорт электронов, встроены в тилакоидные мембраны - скорость
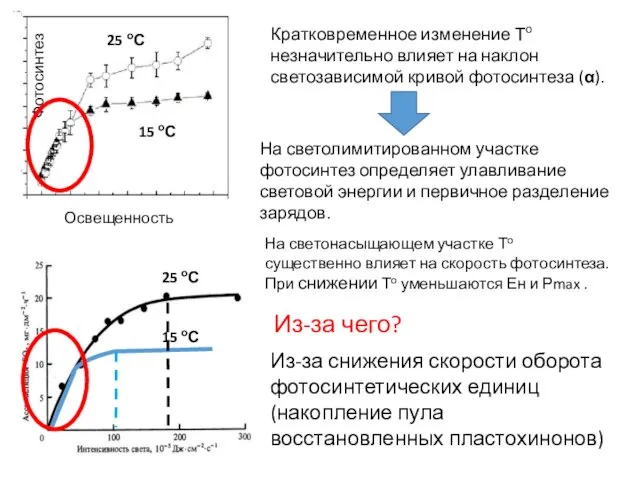
На светолимитированном участке фотосинтез определяет улавливание световой энергии и первичное разделение
На светолимитированном участке фотосинтез определяет улавливание световой энергии и первичное разделение
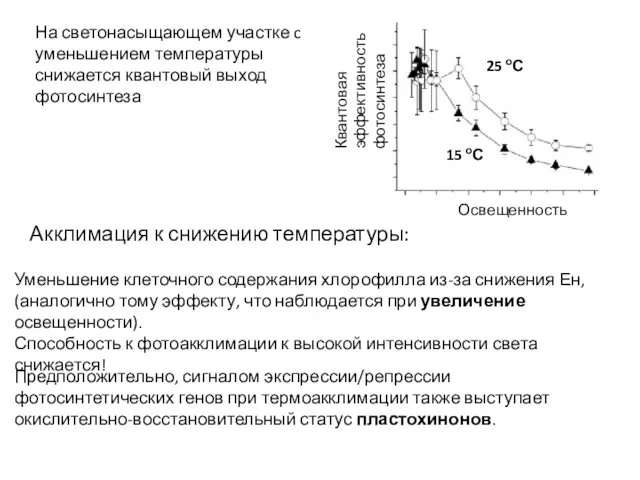
Освещенность
Квантовая эффективность фотосинтеза
25 оС
15 оС
На светонасыщающем участке c уменьшением температуры снижается
Освещенность
Квантовая эффективность фотосинтеза
25 оС
15 оС
На светонасыщающем участке c уменьшением температуры снижается
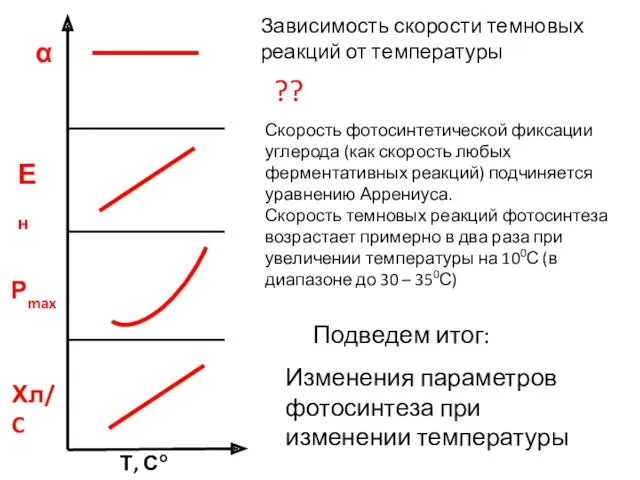
α
Рmax
Ен
Хл/C
Т, Со
Изменения параметров фотосинтеза при изменении температуры
Скорость фотосинтетической фиксации углерода
α
Рmax
Ен
Хл/C
Т, Со
Изменения параметров фотосинтеза при изменении температуры
Скорость фотосинтетической фиксации углерода
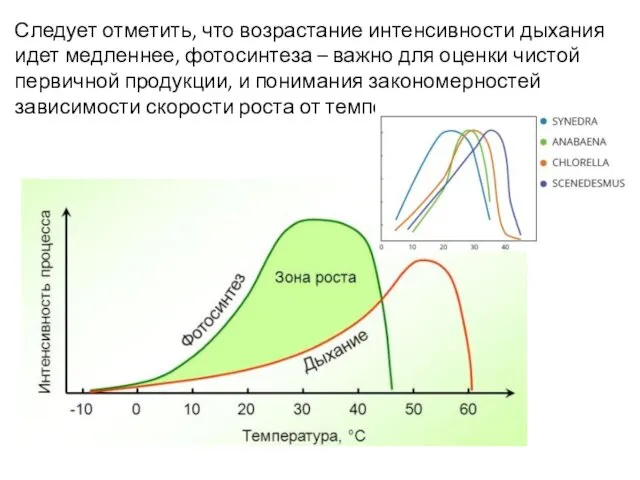
Следует отметить, что возрастание интенсивности дыхания идет медленнее, фотосинтеза – важно
Следует отметить, что возрастание интенсивности дыхания идет медленнее, фотосинтеза – важно
Увеличение клеточных потребностей в азоте, и если его НЕ хватает –
Увеличение клеточных потребностей в азоте, и если его НЕ хватает –
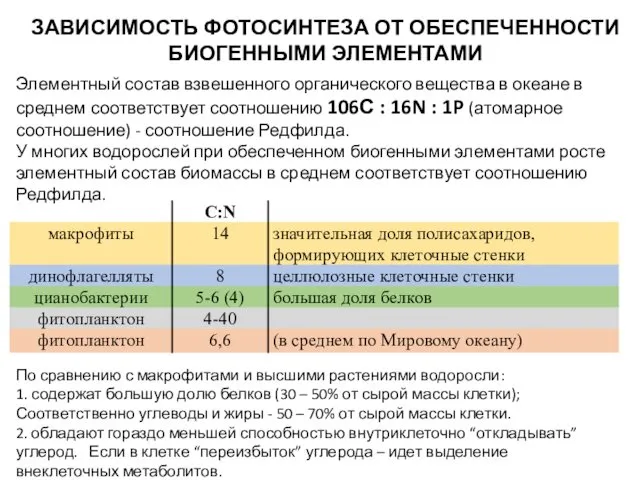
ЗАВИСИМОСТЬ ФОТОСИНТЕЗА ОТ ОБЕСПЕЧЕННОСТИ БИОГЕННЫМИ ЭЛЕМЕНТАМИ
Элементный состав взвешенного органического вещества в
ЗАВИСИМОСТЬ ФОТОСИНТЕЗА ОТ ОБЕСПЕЧЕННОСТИ БИОГЕННЫМИ ЭЛЕМЕНТАМИ
Элементный состав взвешенного органического вещества в

Исходя из соотношения Редфилда:
Значения N:P, близкие к соотношению Редфилда, свидетельствуют об
Исходя из соотношения Редфилда:
Значения N:P, близкие к соотношению Редфилда, свидетельствуют об
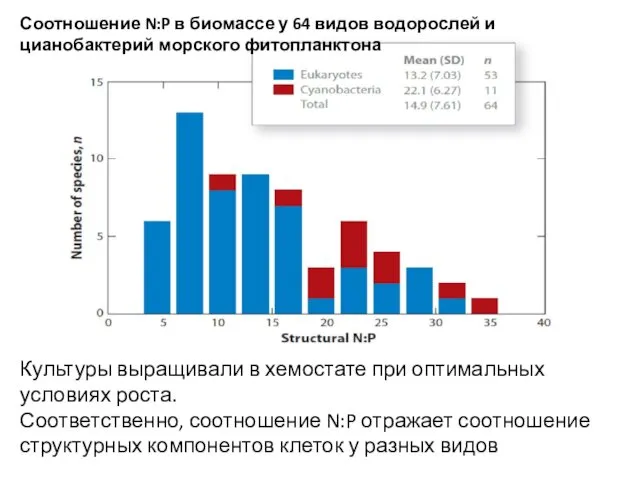
Культуры выращивали в хемостате при оптимальных условиях роста.
Соответственно, соотношение N:P
Культуры выращивали в хемостате при оптимальных условиях роста.
Соответственно, соотношение N:P

АЗОТ
Формы азота в водных экосистемах: молекулярный азот (наибольшее количество), нитраты, нитриты,
АЗОТ
Формы азота в водных экосистемах: молекулярный азот (наибольшее количество), нитраты, нитриты,
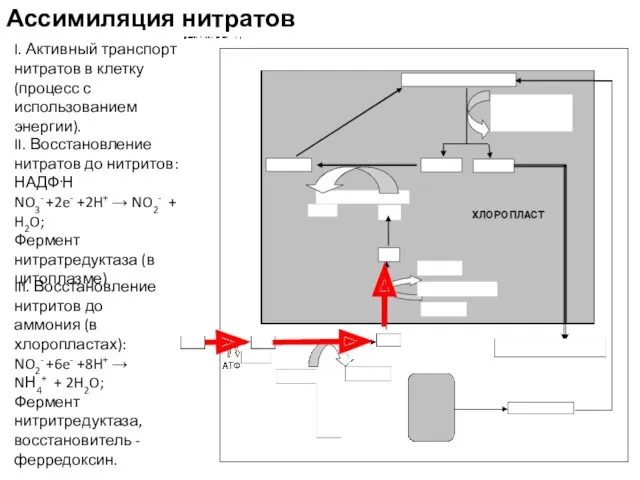
Ассимиляция нитратов
I. Активный транспорт нитратов в клетку (процесс с использованием энергии).
Ассимиляция нитратов
I. Активный транспорт нитратов в клетку (процесс с использованием энергии).
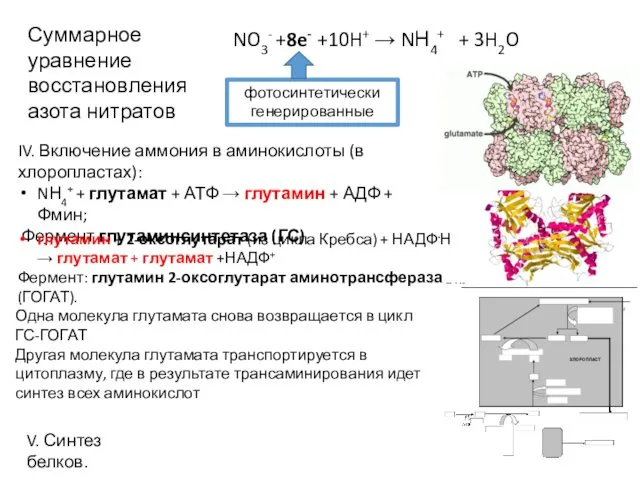
NO3- +8e- +10H+ → NН4+ + 3H2O
фотосинтетически генерированные
IV. Включение аммония
NO3- +8e- +10H+ → NН4+ + 3H2O
фотосинтетически генерированные
IV. Включение аммония
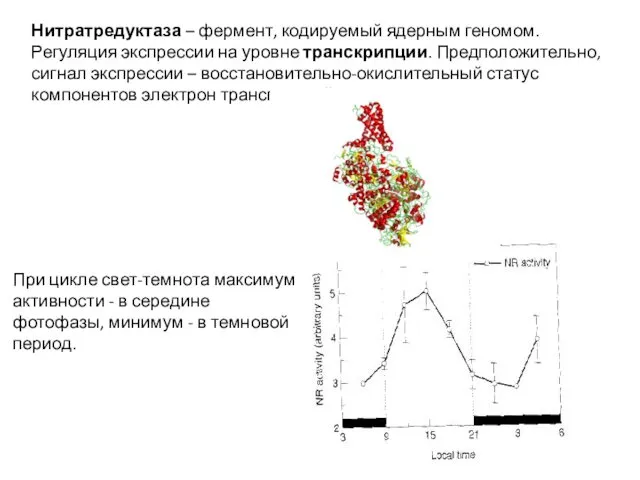
Нитратредуктаза – фермент, кодируемый ядерным геномом.
Регуляция экспрессии на уровне транскрипции.
Нитратредуктаза – фермент, кодируемый ядерным геномом.
Регуляция экспрессии на уровне транскрипции.

При освещенности ниже насыщающей (светолимитированный участок Р-Е кривой) в зависимости от
При освещенности ниже насыщающей (светолимитированный участок Р-Е кривой) в зависимости от

Скорость ассимиляции минерального азота зависит от количества углеродных скелетов, восстановителя и
Скорость ассимиляции минерального азота зависит от количества углеродных скелетов, восстановителя и

Популяционный уровень: снижение скорости роста, при увеличении степени клеточного дефицита азота
Популяционный уровень: снижение скорости роста, при увеличении степени клеточного дефицита азота
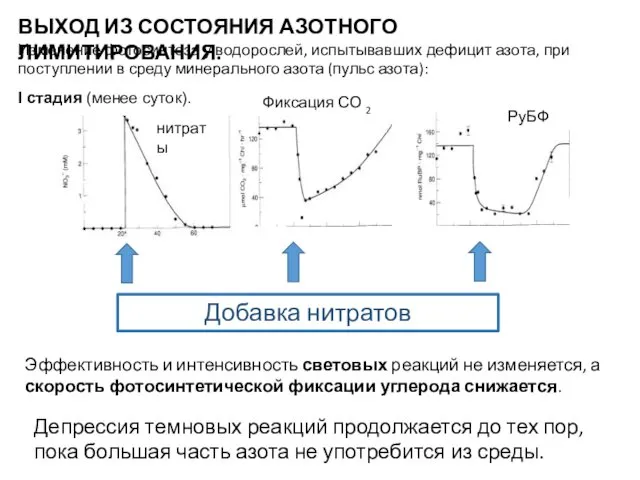
ВЫХОД ИЗ СОСТОЯНИЯ АЗОТНОГО ЛИМИТИРОВАНИЯ.
Изменение фотосинтеза у водорослей, испытывавших дефицит азота,
ВЫХОД ИЗ СОСТОЯНИЯ АЗОТНОГО ЛИМИТИРОВАНИЯ.
Изменение фотосинтеза у водорослей, испытывавших дефицит азота,

Причины:
1. Конкуренция между процессами ассимиляции нитратов (нитритов и аммония) и
Причины:
1. Конкуренция между процессами ассимиляции нитратов (нитритов и аммония) и

II стадия (более суток). Скорость фотосинтетической фиксации углерода возрастает:
Световые реакции:
(а) возрастание
II стадия (более суток). Скорость фотосинтетической фиксации углерода возрастает:
Световые реакции:
(а) возрастание
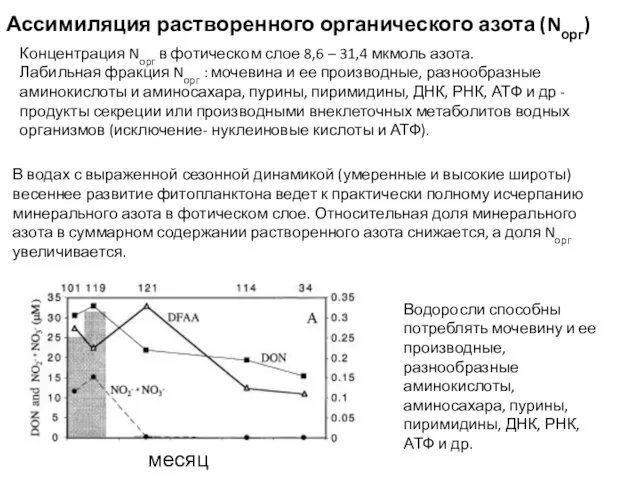
Концентрация Nорг в фотическом слое 8,6 – 31,4 мкмоль азота.
Лабильная
Концентрация Nорг в фотическом слое 8,6 – 31,4 мкмоль азота.
Лабильная

ПОТРЕБЛЕНИЕ МОЧЕВИНЫ
Потребление мочевины идет за счет активного транспорта при использовании фотосинтетически
ПОТРЕБЛЕНИЕ МОЧЕВИНЫ
Потребление мочевины идет за счет активного транспорта при использовании фотосинтетически

ПОТРЕБЛЕНИЕ АМИНОКИСЛОТ
Потребление аминокислот - активный транспорт.
Предположительно: наличие трех транспортных систем
ПОТРЕБЛЕНИЕ АМИНОКИСЛОТ
Потребление аминокислот - активный транспорт.
Предположительно: наличие трех транспортных систем

Потребление аминокислот зависит от вида водорослей и обеспеченности Nмин:
Потребление аминокислот зависит от вида водорослей и обеспеченности Nмин:
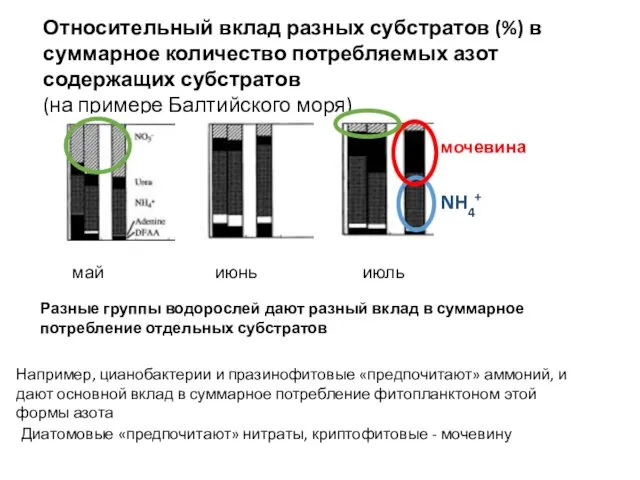
Относительный вклад разных субстратов (%) в суммарное количество потребляемых азот содержащих
Относительный вклад разных субстратов (%) в суммарное количество потребляемых азот содержащих
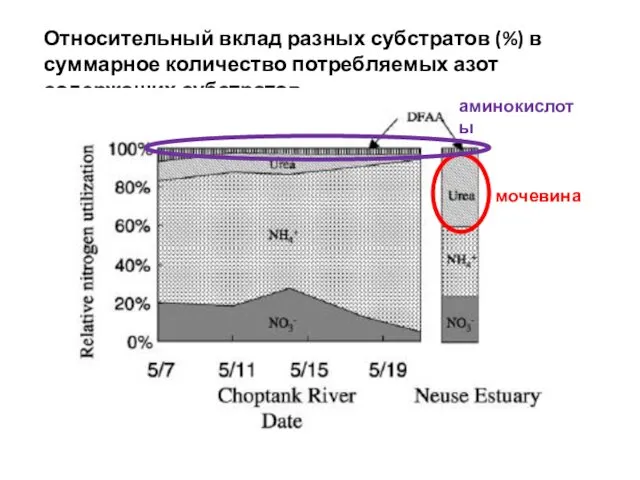
Относительный вклад разных субстратов (%) в суммарное количество потребляемых азот содержащих
Относительный вклад разных субстратов (%) в суммарное количество потребляемых азот содержащих

Лимитирование фотосинтеза недостатком фосфора:
Ограничение синтеза нуклеиновых кислот.
Проявляется на уровне
Лимитирование фотосинтеза недостатком фосфора:
Ограничение синтеза нуклеиновых кислот.
Проявляется на уровне

Концентрация Pорг в поверхностных морских водах может достигать 1 мкмоль и
Концентрация Pорг в поверхностных морских водах может достигать 1 мкмоль и

Два типа фосфатаз: (1) конститутивные фосфатазы, активность которых не зависит от
Два типа фосфатаз: (1) конститутивные фосфатазы, активность которых не зависит от
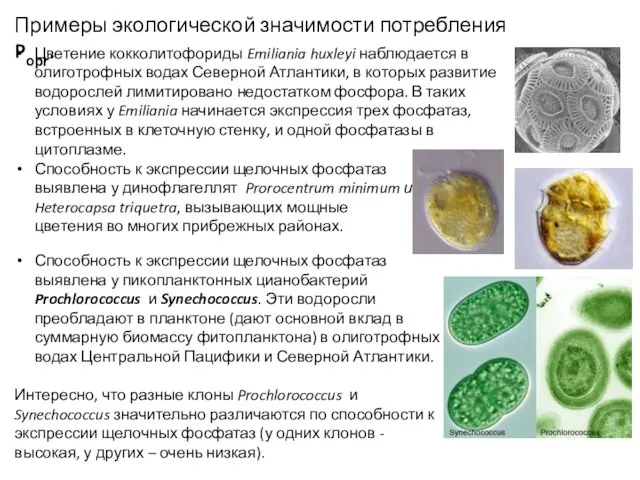
Способность к экспрессии щелочных фосфатаз выявлена у пикопланктонных цианобактерий Prochlorococcus и
Способность к экспрессии щелочных фосфатаз выявлена у пикопланктонных цианобактерий Prochlorococcus и

Лимитирование недостатком серы ведет к снижению скорости синтеза фотосинтетических пигментов и
Лимитирование недостатком серы ведет к снижению скорости синтеза фотосинтетических пигментов и

Лимитирование недостатком железа:
разобщение антенны ФС2 с РЦ ФС2: накопление неактивных РЦ
Лимитирование недостатком железа:
разобщение антенны ФС2 с РЦ ФС2: накопление неактивных РЦ
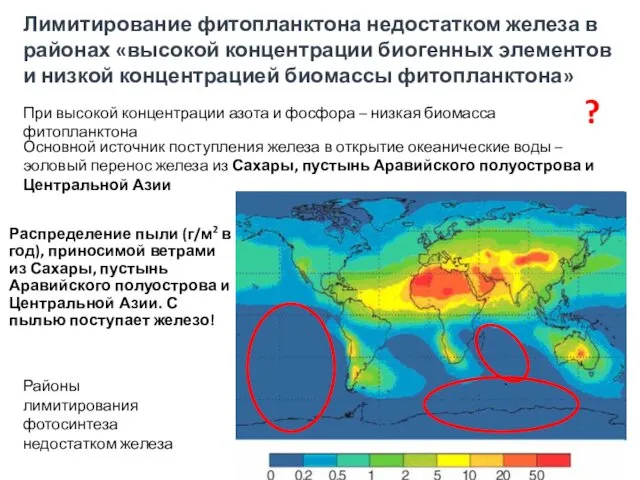
Распределение пыли (г/м2 в год), приносимой ветрами из Сахары, пустынь Аравийского
Распределение пыли (г/м2 в год), приносимой ветрами из Сахары, пустынь Аравийского
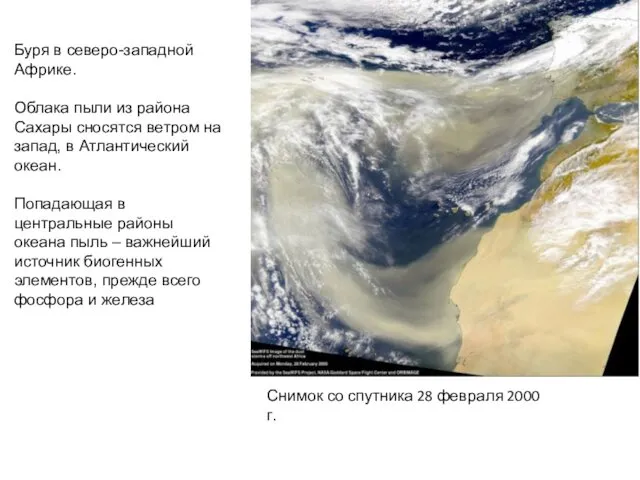
Буря в северо-западной Африке.
Облака пыли из района Сахары сносятся ветром
Буря в северо-западной Африке.
Облака пыли из района Сахары сносятся ветром

Фиксация молекулярного азота
Чтобы разорвать прочную тройную связь в молекуле N2, нужны
Фиксация молекулярного азота
Чтобы разорвать прочную тройную связь в молекуле N2, нужны
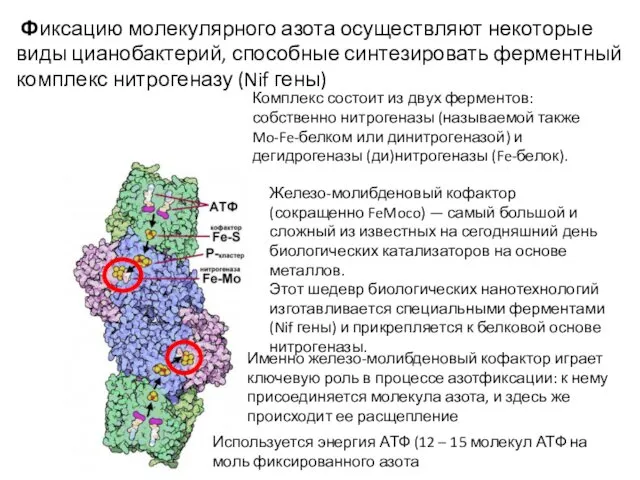
Комплекс состоит из двух ферментов: собственно нитрогеназы (называемой также Mo-Fe-белком или
Комплекс состоит из двух ферментов: собственно нитрогеназы (называемой также Mo-Fe-белком или

Новые данные (на основе молекулярно-генетических методов):
Процесс азотфиксации «стартовал» по крайней мере
Новые данные (на основе молекулярно-генетических методов):
Процесс азотфиксации «стартовал» по крайней мере

Гетероцисты – результат клеточной дифференциации.
Дезактивация ФС2: хл-белковые комплексы и компоненты ФС2
Гетероцисты – результат клеточной дифференциации.
Дезактивация ФС2: хл-белковые комплексы и компоненты ФС2
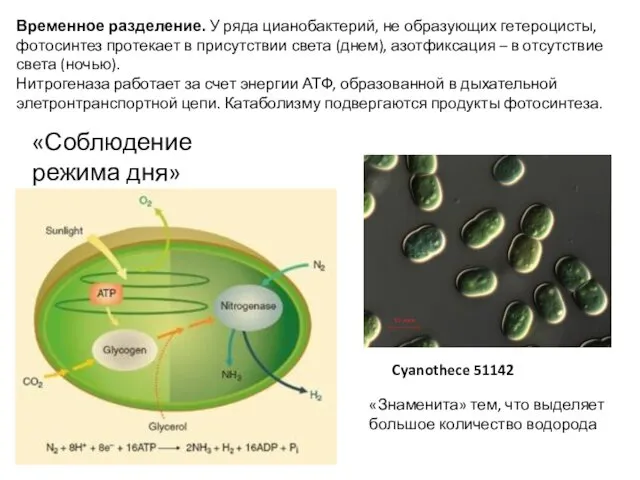
Cyanothece 51142
Временное разделение. У ряда цианобактерий, не образующих гетероцисты, фотосинтез протекает
Cyanothece 51142
Временное разделение. У ряда цианобактерий, не образующих гетероцисты, фотосинтез протекает
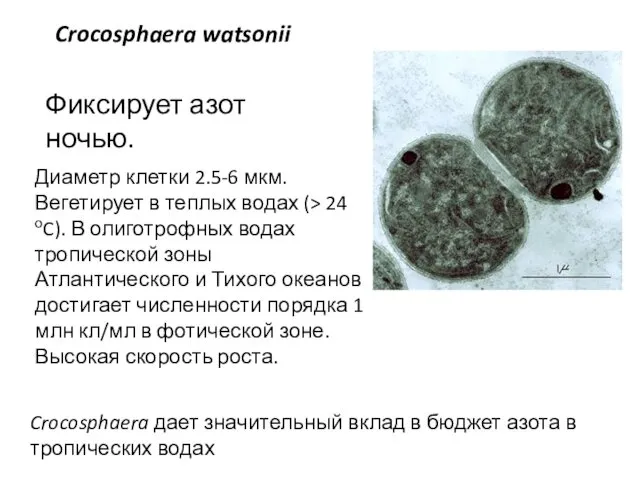
Диаметр клетки 2.5-6 мкм. Вегетирует в теплых водах (> 24 оC).
Диаметр клетки 2.5-6 мкм. Вегетирует в теплых водах (> 24 оC).

Trichodesmium spp.
Цианобактерии Trichodesmium фиксируют азот ТОЛЬКО на свету и при этом
Trichodesmium spp.
Цианобактерии Trichodesmium фиксируют азот ТОЛЬКО на свету и при этом
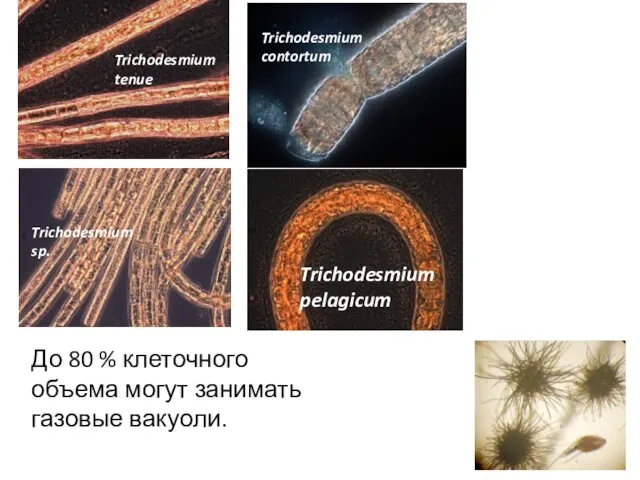
Trichodesmium tenue
Trichodesmium sp.
Trichodesmium contortum
Trichodesmium pelagicum
До 80 % клеточного объема могут занимать
Trichodesmium tenue
Trichodesmium sp.
Trichodesmium contortum
Trichodesmium pelagicum
До 80 % клеточного объема могут занимать

Как же осуществляется защита нитрогеназ от фотосинтетически выделяемого кислорода у Trichodesmium?
Как
Как же осуществляется защита нитрогеназ от фотосинтетически выделяемого кислорода у Trichodesmium?
Как

Разделение азотфиксации и фотосинтеза в пространстве у Trichodesmium
Окраска специфическим красителем Nif
Разделение азотфиксации и фотосинтеза в пространстве у Trichodesmium
Окраска специфическим красителем Nif

Разделение азотфиксации и фотосинтеза во времени у Trichodesmium
Суточный ритм регулирует транскрипцию
Разделение азотфиксации и фотосинтеза во времени у Trichodesmium
Суточный ритм регулирует транскрипцию

Способность к фиксации СО2 также снижается в диазоцитах Trichodesmium во время
Способность к фиксации СО2 также снижается в диазоцитах Trichodesmium во время

Механизмы предотвращения блокирования нитрогеназ фотосинтетически выделяемым кислородом, ПРЕДПОЛОЖИТЕЛЬНО, также включают:
тетразол
Формирование в
Механизмы предотвращения блокирования нитрогеназ фотосинтетически выделяемым кислородом, ПРЕДПОЛОЖИТЕЛЬНО, также включают:
тетразол
Формирование в
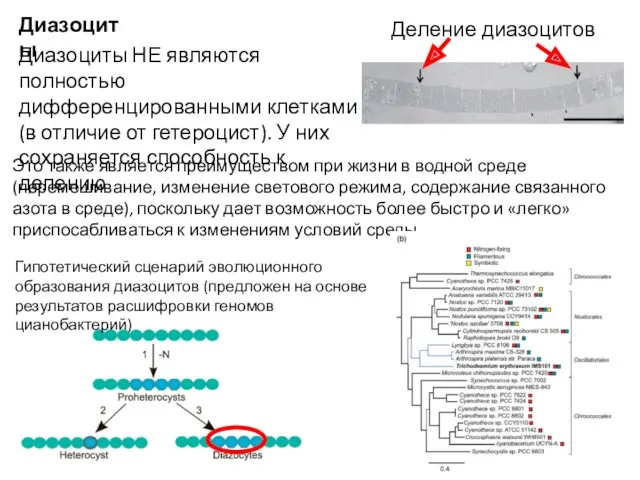
Гипотетический сценарий эволюционного образования диазоцитов (предложен на основе результатов расшифровки геномов
Гипотетический сценарий эволюционного образования диазоцитов (предложен на основе результатов расшифровки геномов

Дальнейшая ассимиляция глутамата, образованного при азотфиксации, происходит в глутаминсинтетазо - глутамин
Дальнейшая ассимиляция глутамата, образованного при азотфиксации, происходит в глутаминсинтетазо - глутамин
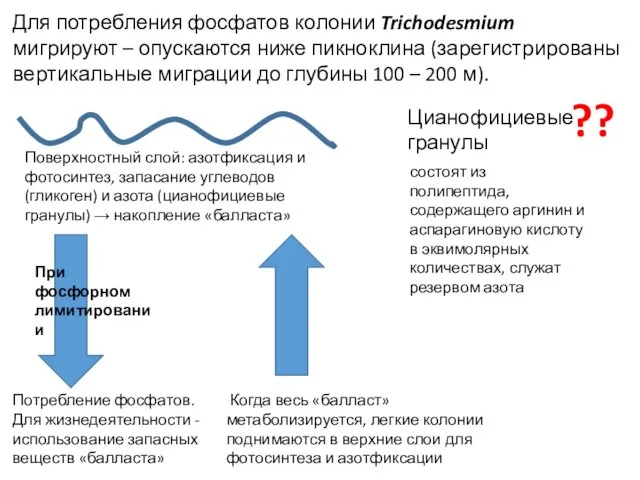
Для потребления фосфатов колонии Trichodesmium мигрируют – опускаются ниже пикноклина (зарегистрированы
Для потребления фосфатов колонии Trichodesmium мигрируют – опускаются ниже пикноклина (зарегистрированы
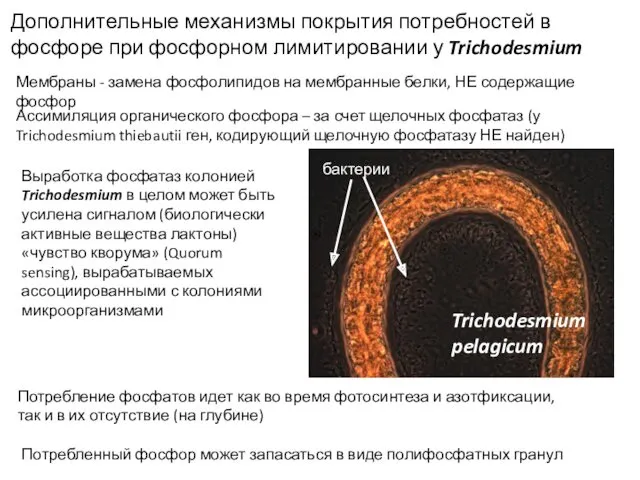
Дополнительные механизмы покрытия потребностей в фосфоре при фосфорном лимитировании у Trichodesmium
Мембраны
Дополнительные механизмы покрытия потребностей в фосфоре при фосфорном лимитировании у Trichodesmium
Мембраны

Обеспечение потребностей в железе у Trichodesmium
В клетках Trichodesmium высокое содержание
Обеспечение потребностей в железе у Trichodesmium
В клетках Trichodesmium высокое содержание

Лимитирование недостатком железа ведет к снижению числа диазоцитов и скорости азофиксации
А
Лимитирование недостатком железа ведет к снижению числа диазоцитов и скорости азофиксации
А

Экосистемная роль Trichodesmium
Пределы температурной толерантности для роста и азотфиксации у
Экосистемная роль Trichodesmium
Пределы температурной толерантности для роста и азотфиксации у

Цветение Trichodesmium.
Большой барьерный риф побережья Австралии.
Снимок NASA
Цветение Trichodesmium.
Красное море
Снимок
Цветение Trichodesmium.
Большой барьерный риф побережья Австралии.
Снимок NASA
Цветение Trichodesmium.
Красное море
Снимок

Для цветений Trichodesmium характерно как прижизненное выделение значительно количества органического вещества,
Для цветений Trichodesmium характерно как прижизненное выделение значительно количества органического вещества,

Это довольно НЕОЖИДАННЫЙ результат, поскольку Trichodesmium синтезирует токсины, вызывающие гибель копепод,
Это довольно НЕОЖИДАННЫЙ результат, поскольку Trichodesmium синтезирует токсины, вызывающие гибель копепод,

Прогнозы динамики Trichodesmium при наблюдаемом климатическом тренде
Есть все основания полагать, что
Прогнозы динамики Trichodesmium при наблюдаемом климатическом тренде
Есть все основания полагать, что
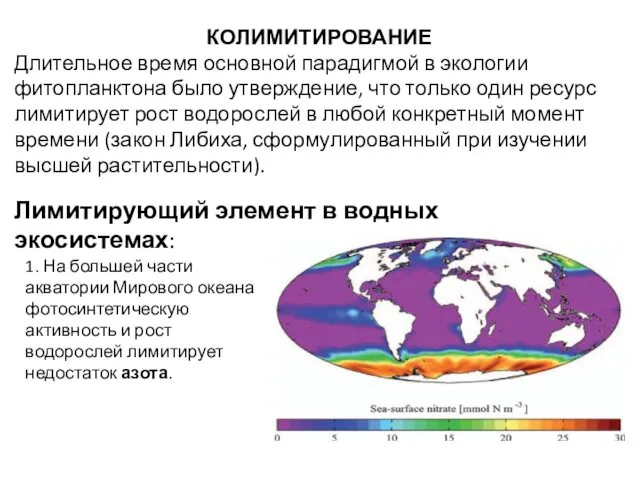
КОЛИМИТИРОВАНИЕ
Длительное время основной парадигмой в экологии фитопланктона было утверждение, что только
КОЛИМИТИРОВАНИЕ
Длительное время основной парадигмой в экологии фитопланктона было утверждение, что только
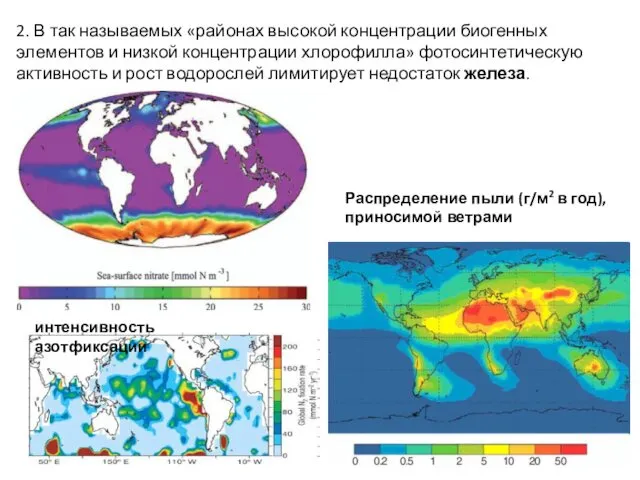
2. В так называемых «районах высокой концентрации биогенных элементов и низкой
2. В так называемых «районах высокой концентрации биогенных элементов и низкой

3. В ряде районов Мирового океана (Северная Атлантика, Средиземное море, Бискайский
3. В ряде районов Мирового океана (Северная Атлантика, Средиземное море, Бискайский

Мультиресурсное ко-лимитирование.
Два ресурса в такой низкой концентрации (ниже порогового уровня),
Мультиресурсное ко-лимитирование.
Два ресурса в такой низкой концентрации (ниже порогового уровня),

Колимитирование на уровне сообщества. Добавка ресурса А стимулирует рост одной популяции,
Колимитирование на уровне сообщества. Добавка ресурса А стимулирует рост одной популяции,
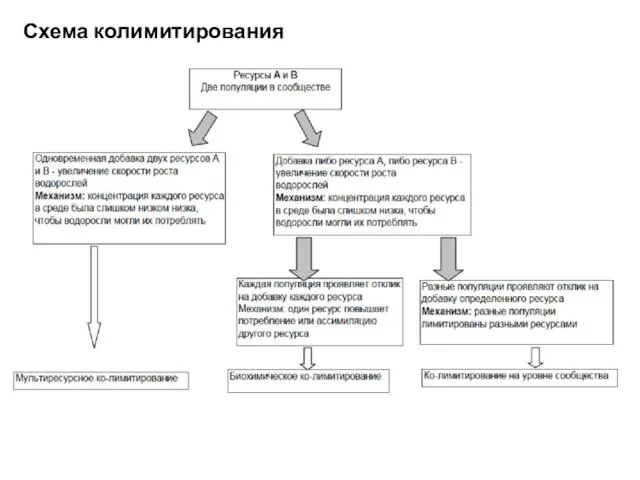
Схема колимитирования
Схема колимитирования

На следующей лекции:
Пространственно-временная изменчивость первичной продукции в Мировом океане
Вопросы?
На следующей лекции:
Пространственно-временная изменчивость первичной продукции в Мировом океане
Вопросы?
 История развития экологии как науки
История развития экологии как науки Призентация приложения к программе Доброморье
Призентация приложения к программе Доброморье Повышение эффективности ТЭУ
Повышение эффективности ТЭУ I wish a had a magic vacuum cleaner, instead of my old broom!
I wish a had a magic vacuum cleaner, instead of my old broom! Приложение 1 к уроку по теме Озёра. Презентация. часть 3
Приложение 1 к уроку по теме Озёра. Презентация. часть 3 Интегративный подход к организации развивающих центров активности детей
Интегративный подход к организации развивающих центров активности детей Интерактивная игра Своя игра. ОРКСЭ. Основы православной культуры.
Интерактивная игра Своя игра. ОРКСЭ. Основы православной культуры. Ливонская война
Ливонская война Концептуальные аспекты управления инновациями
Концептуальные аспекты управления инновациями История русской вышивки. Вышивка крестом
История русской вышивки. Вышивка крестом Геодезическое обеспечение геологоразведочных работ
Геодезическое обеспечение геологоразведочных работ Консультация для родителей второклассников.
Консультация для родителей второклассников. Поздравления с 8 марта
Поздравления с 8 марта Исследовательская работа: Прекрасные и опасные комнатные растения
Исследовательская работа: Прекрасные и опасные комнатные растения Усилители СВЧ
Усилители СВЧ SWOT-анализ
SWOT-анализ Западно-Сибирская низменность
Западно-Сибирская низменность Политическая карта мира
Политическая карта мира Педагогическая академия последипломного образования
Педагогическая академия последипломного образования Презентация кружка Уроки здоровья
Презентация кружка Уроки здоровья Многолучевые и сканирующие антенны. (Лекция 14)
Многолучевые и сканирующие антенны. (Лекция 14) Голландское искусство XVII в
Голландское искусство XVII в правила пожарной безопасности
правила пожарной безопасности Кислородные соединения углерода Диск
Кислородные соединения углерода Диск Метрологическое оборудование от калибратора до лаборатории
Метрологическое оборудование от калибратора до лаборатории Отморожения. Классификация
Отморожения. Классификация Последние дни земной жизни Господа нашего Иисуса Христа. Вход Господень в Иерусалим
Последние дни земной жизни Господа нашего Иисуса Христа. Вход Господень в Иерусалим Принтерлермен жұмыс істеу
Принтерлермен жұмыс істеу