Водоросли. Особенности строения, размножения и экология. Принципы классификации. Эволюция презентация
- Водоросли. Особенности строения, размножения и экология. Принципы классификации. Эволюция

Содержание
- 2. Водоросли— гетерогенная экологическая группа преимущественно фототрофных одноклеточных, колониальных или многоклеточных организмов, обитающих, как правило, в водной
- 3. Особенности водорослей наличие хлорофилла и фотоавтотрофного питания (некоторые водоросли способны к гетеротрофии: жгутиконосцы, эвгленовые, динофитовые). у
- 4. Типы дифференциации таллома Монадный ТДТ имеют постоянную форму тела, жгутик(и), часто стигму, а пресноводные — сократительную
- 5. Нитчатый (трихальный) ТДТ Клетки соединены в нить, простую или разветвлённую. Нити могут свободно плавать в толще
- 6. Строение клетки эукариотических водорослей Клетки водорослей — вполне типичные для эукариот. Основные отличия — на биохимическом
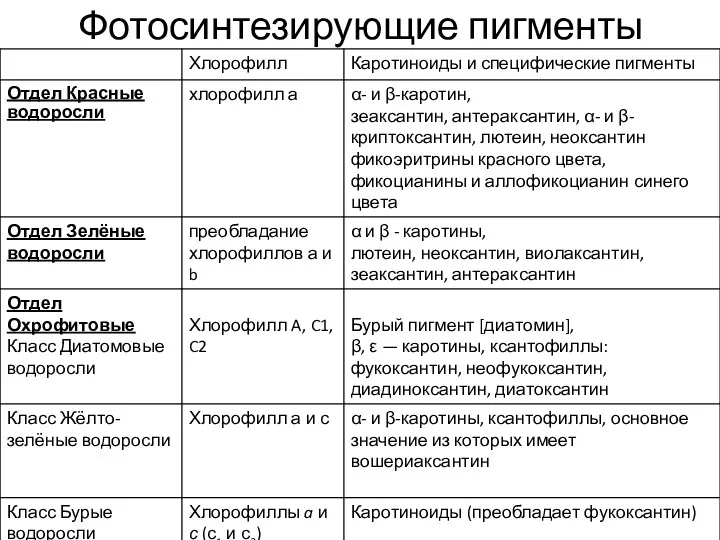
- 7. Фотосинтезирующие пигменты находятся в особых пластидах — хлоропластах. Хлоропласт имеет несколько мембран: две (красные, зелёные, харовые
- 8. Фотосинтезирующие пигменты
- 9. Жгутики Жгутиков у подвижных клеток чаще всего 2, реже — 1, 4 или много (до 120);
- 10. Размножение Бесполое Вегетативное: Бинарное деление клетки (хламидмонада) Акинеты – специализированные толстостенные клетки с запасом питательных веществ
- 11. Половое размножение: Саматогамия-слияние вегетативных клеток: Хологамия- слияние двух однокл. организмов (хламидомонада) Коньюгация- слияние содержимого двух вегетатив.
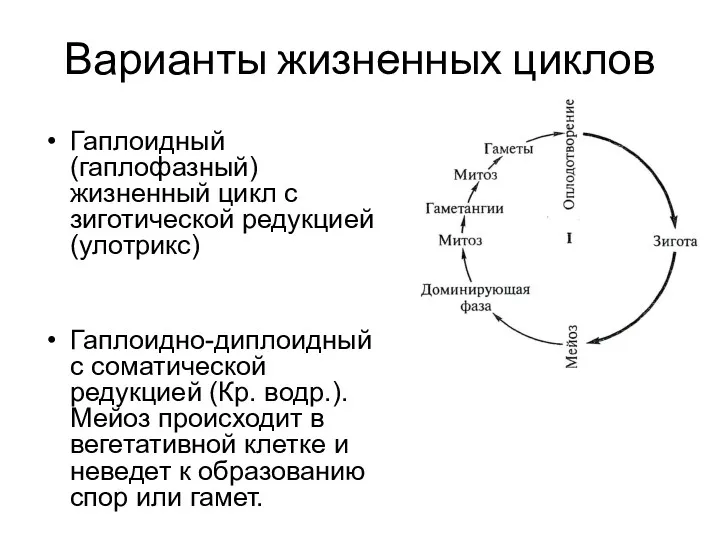
- 12. Варианты жизненных циклов Гаплоидный (гаплофазный) жизненный цикл с зиготической редукцией (улотрикс) Гаплоидно-диплоидный с соматической редукцией (Кр.
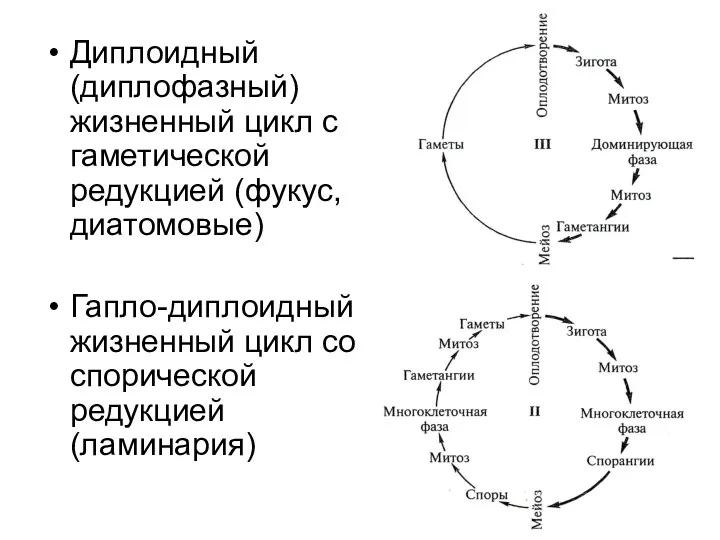
- 13. Диплоидный (диплофазный) жизненный цикл с гаметической редукцией (фукус, диатомовые) Гапло-диплоидный жизненный цикл со спорической редукцией (ламинария)
- 14. Экология Водоросли — главные производители органических веществ в водной среде. Около 80 % всех органических веществ,
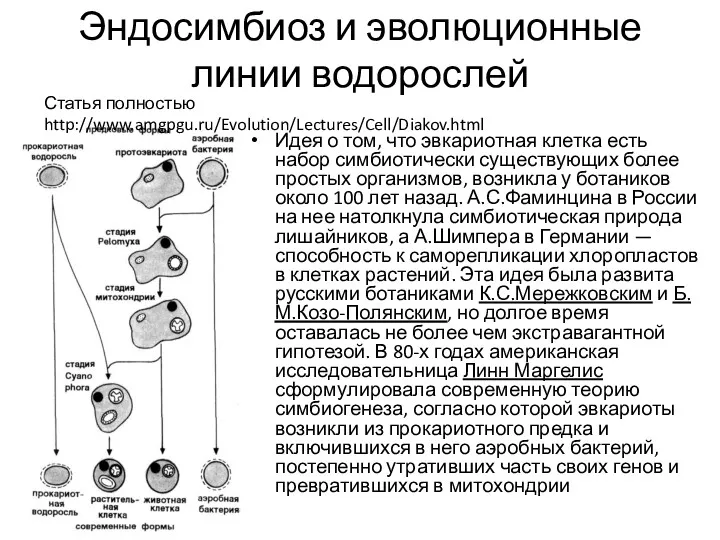
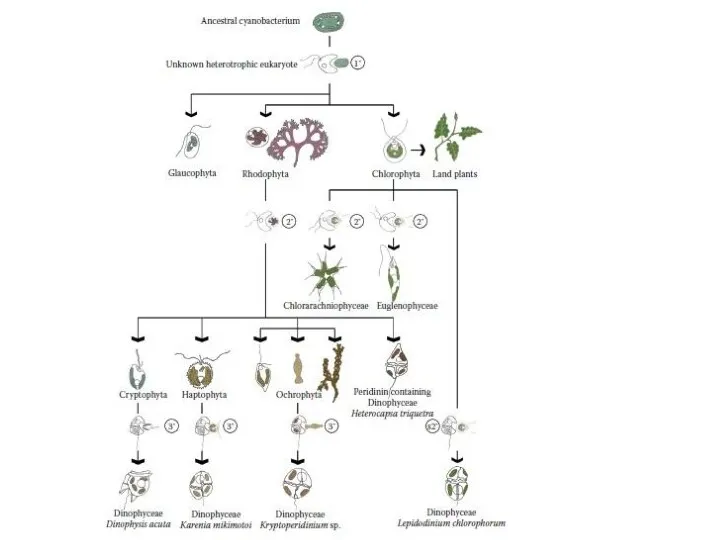
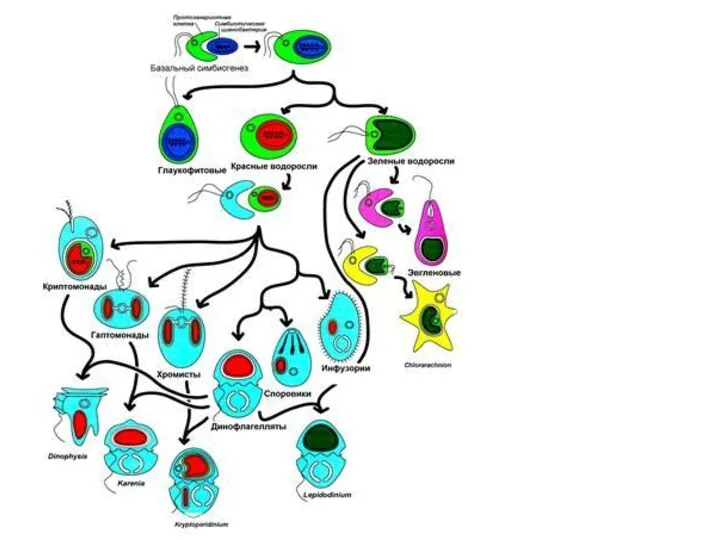
- 15. Эндосимбиоз и эволюционные линии водорослей Идея о том, что эвкариотная клетка есть набор симбиотически существующих более
- 16. Хлоропласты по симбиогенетической теории произошли из организмов, сходных с современными цианобактериями (сине-зелеными водорослями), которых поглотил амебоидный
- 17. Многие эвкариотные водоросли выполняют функции пластид у морских и пресноводных беспозвоночных. Если в клетках живут одноклеточные
- 18. Как известно, большинство отделов водорослей названы по преобладающему цвету их клеток (красные, бурые и т.д.). Это
- 19. Интересные данные получены при изучении оболочек, покрывающих хроматофоры. Только у красных и зеленых водорослей оболочка состоит
- 20. Первая эволюционная линия Сходство строения и пигментного состава фотосинтетического аппарата сине-зеленых и красных водорослей указывает на
- 21. Вторая эволюционная линия Зеленые и эвгленовые водоросли. В 1975 г. была найдена одноклеточная прокариотная морская водоросль
- 22. У эвглен третья мембрана хлоропласта свидетельствует о его вторичном происхождении. По ряду признаков эвгленовые водоросли близки
- 23. Третья эволюционная линия Эволюция линии хромофит связана с приобретением хлорофилла с и специфического набора каротиноидов. Непосредственный
- 24. Согласно гипотетической схеме эндосимбиотической эволюции водорослей, можно выделить три эволюционные линии Родофиты Багрянки — тупиковая ветвь
- 25. Таким образом, у водорослей наблюдается три типа эволюции. Параллельные ряды морфологической эволюции (монадный, пальмеллоидный, коккоидный, нитчатый
- 29. Скачать презентацию
Водоросли—
гетерогенная экологическая группа преимущественно фототрофных одноклеточных, колониальных или многоклеточных организмов, обитающих, как правило,
Водоросли—
гетерогенная экологическая группа преимущественно фототрофных одноклеточных, колониальных или многоклеточных организмов, обитающих, как правило,

Особенности водорослей
наличие хлорофилла и фотоавтотрофного питания (некоторые водоросли способны к гетеротрофии: жгутиконосцы, эвгленовые,
Особенности водорослей
наличие хлорофилла и фотоавтотрофного питания (некоторые водоросли способны к гетеротрофии: жгутиконосцы, эвгленовые,

Типы дифференциации таллома
Монадный ТДТ
имеют постоянную форму тела, жгутик(и), часто стигму, а пресноводные — сократительную
Типы дифференциации таллома
Монадный ТДТ
имеют постоянную форму тела, жгутик(и), часто стигму, а пресноводные — сократительную

Нитчатый (трихальный) ТДТ
Клетки соединены в нить, простую или разветвлённую. Нити могут свободно плавать
Нитчатый (трихальный) ТДТ
Клетки соединены в нить, простую или разветвлённую. Нити могут свободно плавать

Строение клетки эукариотических водорослей
Клетки водорослей — вполне типичные для эукариот.
Основные отличия — на биохимическом
Строение клетки эукариотических водорослей
Клетки водорослей — вполне типичные для эукариот.
Основные отличия — на биохимическом

Фотосинтезирующие пигменты находятся в особых пластидах — хлоропластах.
Хлоропласт имеет несколько мембран:
две (красные, зелёные,
Фотосинтезирующие пигменты находятся в особых пластидах — хлоропластах.
Хлоропласт имеет несколько мембран:
две (красные, зелёные,

Фотосинтезирующие пигменты
Фотосинтезирующие пигменты
Жгутики
Жгутиков у подвижных клеток чаще всего 2, реже — 1, 4 или много
Жгутики
Жгутиков у подвижных клеток чаще всего 2, реже — 1, 4 или много

Размножение
Бесполое
Вегетативное:
Бинарное деление клетки (хламидмонада)
Акинеты – специализированные толстостенные клетки с запасом питательных веществ
Размножение
Бесполое
Вегетативное:
Бинарное деление клетки (хламидмонада)
Акинеты – специализированные толстостенные клетки с запасом питательных веществ

Половое размножение:
Саматогамия-слияние вегетативных клеток:
Хологамия- слияние двух однокл. организмов (хламидомонада)
Коньюгация- слияние содержимого двух вегетатив.
Половое размножение:
Саматогамия-слияние вегетативных клеток:
Хологамия- слияние двух однокл. организмов (хламидомонада)
Коньюгация- слияние содержимого двух вегетатив.

Варианты жизненных циклов
Гаплоидный (гаплофазный) жизненный цикл с зиготической редукцией (улотрикс)
Гаплоидно-диплоидный с соматической редукцией
Варианты жизненных циклов
Гаплоидный (гаплофазный) жизненный цикл с зиготической редукцией (улотрикс)
Гаплоидно-диплоидный с соматической редукцией
Диплоидный (диплофазный) жизненный цикл с гаметической редукцией (фукус, диатомовые)
Гапло-диплоидный жизненный цикл со спорической
Диплоидный (диплофазный) жизненный цикл с гаметической редукцией (фукус, диатомовые)
Гапло-диплоидный жизненный цикл со спорической
Экология
Водоросли — главные производители органических веществ в водной среде. Около 80 % всех органических веществ,
Экология
Водоросли — главные производители органических веществ в водной среде. Около 80 % всех органических веществ,

Эндосимбиоз и эволюционные линии водорослей
Идея о том, что эвкариотная клетка есть набор симбиотически
Эндосимбиоз и эволюционные линии водорослей
Идея о том, что эвкариотная клетка есть набор симбиотически
Хлоропласты по симбиогенетической теории произошли из организмов, сходных с современными цианобактериями (сине-зелеными водорослями),
Хлоропласты по симбиогенетической теории произошли из организмов, сходных с современными цианобактериями (сине-зелеными водорослями),

Многие эвкариотные водоросли выполняют функции пластид у морских и пресноводных беспозвоночных. Если в
Многие эвкариотные водоросли выполняют функции пластид у морских и пресноводных беспозвоночных. Если в

Как известно, большинство отделов водорослей названы по преобладающему цвету их клеток (красные, бурые
Как известно, большинство отделов водорослей названы по преобладающему цвету их клеток (красные, бурые

Интересные данные получены при изучении оболочек, покрывающих хроматофоры.
Только у красных и зеленых
Интересные данные получены при изучении оболочек, покрывающих хроматофоры.
Только у красных и зеленых

Первая эволюционная линия
Сходство строения и пигментного состава фотосинтетического аппарата сине-зеленых и красных водорослей
Первая эволюционная линия
Сходство строения и пигментного состава фотосинтетического аппарата сине-зеленых и красных водорослей

Вторая эволюционная линия
Зеленые и эвгленовые водоросли.
В 1975 г. была найдена одноклеточная прокариотная
Вторая эволюционная линия
Зеленые и эвгленовые водоросли.
В 1975 г. была найдена одноклеточная прокариотная

У эвглен третья мембрана хлоропласта свидетельствует о его вторичном происхождении. По ряду признаков эвгленовые
У эвглен третья мембрана хлоропласта свидетельствует о его вторичном происхождении. По ряду признаков эвгленовые

Третья эволюционная линия
Эволюция линии хромофит связана с приобретением хлорофилла с и специфического набора
Третья эволюционная линия
Эволюция линии хромофит связана с приобретением хлорофилла с и специфического набора

Согласно гипотетической схеме эндосимбиотической эволюции водорослей, можно выделить три эволюционные линии
Родофиты
Багрянки — тупиковая
Согласно гипотетической схеме эндосимбиотической эволюции водорослей, можно выделить три эволюционные линии
Родофиты
Багрянки — тупиковая

Таким образом, у водорослей наблюдается три типа эволюции.
Параллельные ряды морфологической эволюции (монадный, пальмеллоидный, коккоидный, нитчатый
Таким образом, у водорослей наблюдается три типа эволюции.
Параллельные ряды морфологической эволюции (монадный, пальмеллоидный, коккоидный, нитчатый

 Внесение изменений в РД и ПД. Оформление. Памятка
Внесение изменений в РД и ПД. Оформление. Памятка Электронды техника ќўраушылары. Заттыѕ ќўрылысы туралы негізгі тїсініктер
Электронды техника ќўраушылары. Заттыѕ ќўрылысы туралы негізгі тїсініктер Прессовые формовочные машины
Прессовые формовочные машины Павловопосадские платки. Статья и презентация
Павловопосадские платки. Статья и презентация Презентация здравствуй сказка.ppt
Презентация здравствуй сказка.ppt Основы позирования
Основы позирования Developing toys
Developing toys Самый умный пятиклассник
Самый умный пятиклассник Адаптированные общеобразовательные программы для детей с ТНР
Адаптированные общеобразовательные программы для детей с ТНР Теория литературы. Авторская позиция
Теория литературы. Авторская позиция Расцвет Руси при Ярославе Мудром
Расцвет Руси при Ярославе Мудром Патріотизм – дань моді чи самовідданість нації?
Патріотизм – дань моді чи самовідданість нації? Досуг для группы среднего возраста путешествие кленового листочка.
Досуг для группы среднего возраста путешествие кленового листочка. Создание коллажей с помощью Microsoft Research AutoCollage
Создание коллажей с помощью Microsoft Research AutoCollage Recognized an object on graphics which taken from street camers
Recognized an object on graphics which taken from street camers Моря России
Моря России Профессиональный союз работников народного образования и науки Российской Федерации
Профессиональный союз работников народного образования и науки Российской Федерации Викторина Корней Чуковский
Викторина Корней Чуковский Интерактивное ТВ
Интерактивное ТВ Utilajul frigorific din sala de comerț
Utilajul frigorific din sala de comerț “Мое кофе”. Отдых в горных условиях
“Мое кофе”. Отдых в горных условиях Диагностика беременности
Диагностика беременности Регулирование частоты вращения асинхронных двигателей с фазным ротором
Регулирование частоты вращения асинхронных двигателей с фазным ротором Сущность поликультурного образования в Европе
Сущность поликультурного образования в Европе Каркасная система зданий. Лекция № 6
Каркасная система зданий. Лекция № 6 Бродячие собаки
Бродячие собаки Старый мост (Староволжский мост)
Старый мост (Староволжский мост) Выявление отношения спортсменов к употреблению допинга
Выявление отношения спортсменов к употреблению допинга