- Замыкающие клетки устьиц

Содержание
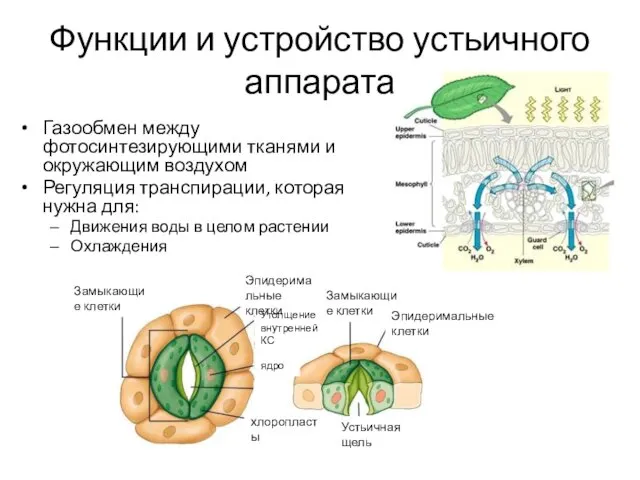
- 2. Функции и устройство устьичного аппарата Газообмен между фотосинтезирующими тканями и окружающим воздухом Регуляция транспирации, которая нужна
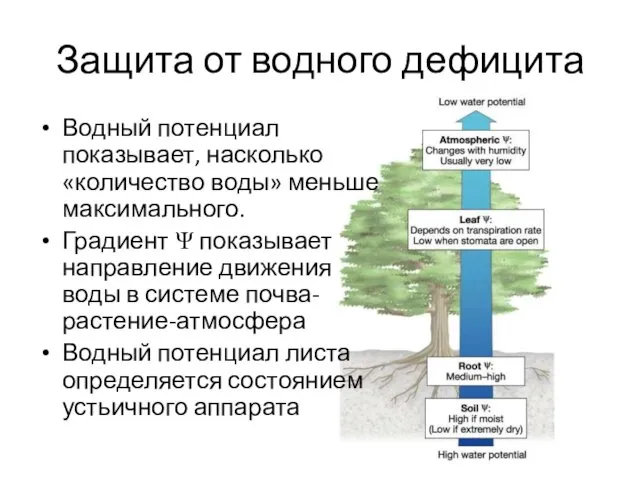
- 3. Защита от водного дефицита Водный потенциал показывает, насколько «количество воды» меньше максимального. Градиент Ψ показывает направление
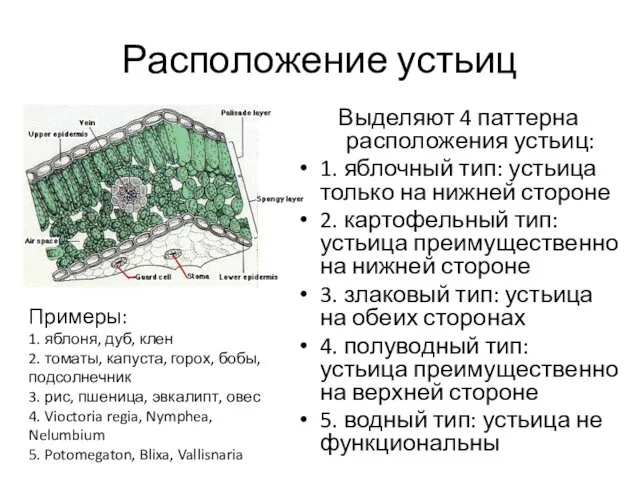
- 4. Расположение устьиц Выделяют 4 паттерна расположения устьиц: 1. яблочный тип: устьица только на нижней стороне 2.
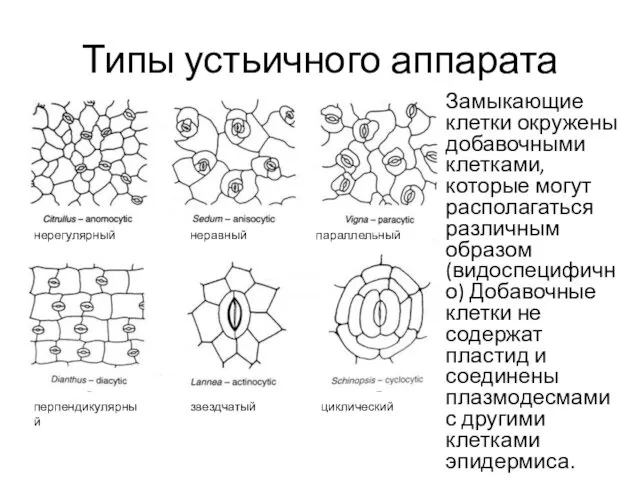
- 5. Типы устьичного аппарата Замыкающие клетки окружены добавочными клетками, которые могут располагаться различным образом (видоспецифично) Добавочные клетки
- 6. Какие факторы регулируют работу устьиц Относительная влажность атмосферы Содержание влаги в почве СВЕТ (суточная ритмичность) Температура
- 7. Какие сигналы чувствуют устьичные клетки? Световой сигнал Гормональный сигнал рецепторы вторичные мессенджеры
- 8. Механизм закрытия Ингибирование Н+-АТФазы и активация анионных каналов приводят к деполяризации. Анионные каналы быстрого типа (R-type)
- 9. Механизм открытия Активация Н-АТФазы ПМ. Ее активируют синий свет и ауксин, ингибируют кальция и АБК. Выход
- 10. Восприятие устьицами светового сигнала Было обнаружено, что синий свет вызывает открывание устьиц. При этом достаточно короткого
- 11. Фототропин Фототропин – белок, содержащий 2 LOV-домена (light, oxygen, voltage), а также киназный домен. На синем
- 12. Механизм передачи сигнала Ключевым элементом регуляции является Н+-АТФаза плазмалеммы. Ее активность регулируется путем фосфорилирования и присоединения
- 14. Восприятие устьицами гормонального сигнала АБК является центральным регулятором устьичного движения. Функционирование сигнального пути зависит от концентрации
- 15. Транспорт АБК Поскольку АБК может транспортироваться на дальние расстояния, она служит надежным стресс-мессенджером. В ЗК экспрессируется
- 16. Рецепция АБК Внутри ЗК АБК связывается с рецептором PYR/PYL/RCAR. Небольшой белок находится в цитоплазме и в
- 17. Запуск ответа на АБК Киназы SNRK регулируют активность ионных каналов и протонной помпы: Они ингибируют Н+-АТФазу,
- 20. Скачать презентацию
Функции и устройство устьичного аппарата
Газообмен между фотосинтезирующими тканями и окружающим воздухом
Регуляция
Функции и устройство устьичного аппарата
Газообмен между фотосинтезирующими тканями и окружающим воздухом
Регуляция
Защита от водного дефицита
Водный потенциал показывает, насколько «количество воды» меньше максимального.
Градиент
Защита от водного дефицита
Водный потенциал показывает, насколько «количество воды» меньше максимального.
Градиент
Расположение устьиц
Выделяют 4 паттерна расположения устьиц:
1. яблочный тип: устьица только на
Расположение устьиц
Выделяют 4 паттерна расположения устьиц:
1. яблочный тип: устьица только на
Типы устьичного аппарата
Замыкающие клетки окружены добавочными клетками, которые могут располагаться различным
Типы устьичного аппарата
Замыкающие клетки окружены добавочными клетками, которые могут располагаться различным
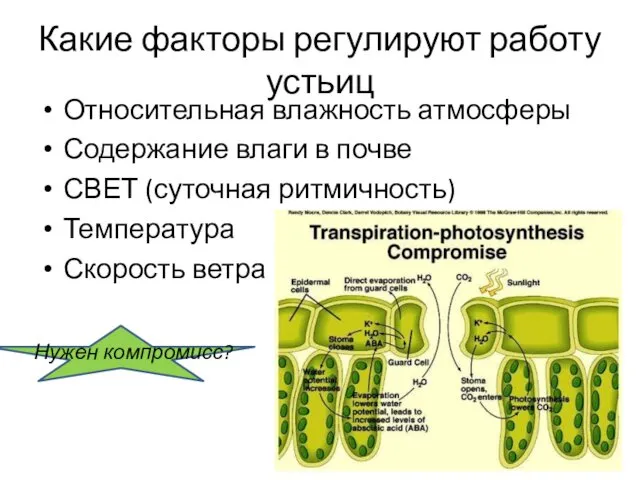
Какие факторы регулируют работу устьиц
Относительная влажность атмосферы
Содержание влаги в почве
СВЕТ (суточная
Какие факторы регулируют работу устьиц
Относительная влажность атмосферы
Содержание влаги в почве
СВЕТ (суточная
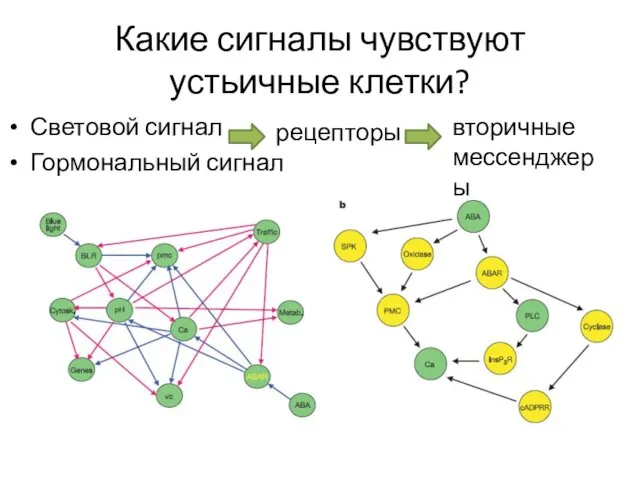
Какие сигналы чувствуют устьичные клетки?
Световой сигнал
Гормональный сигнал
рецепторы
вторичные мессенджеры
Какие сигналы чувствуют устьичные клетки?
Световой сигнал
Гормональный сигнал
рецепторы
вторичные мессенджеры
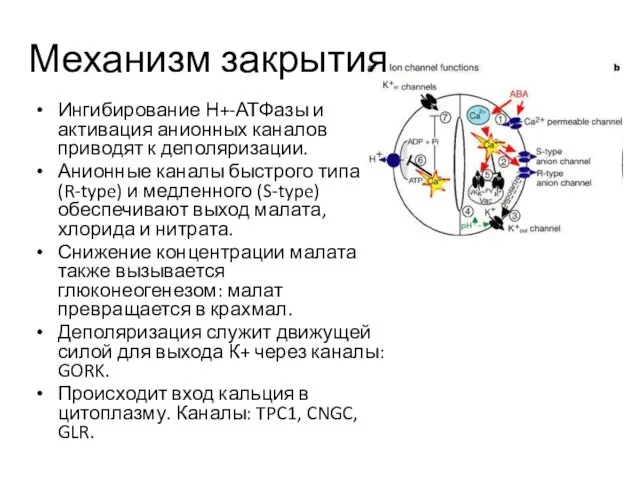
Механизм закрытия
Ингибирование Н+-АТФазы и активация анионных каналов приводят к деполяризации.
Анионные каналы
Механизм закрытия
Ингибирование Н+-АТФазы и активация анионных каналов приводят к деполяризации.
Анионные каналы
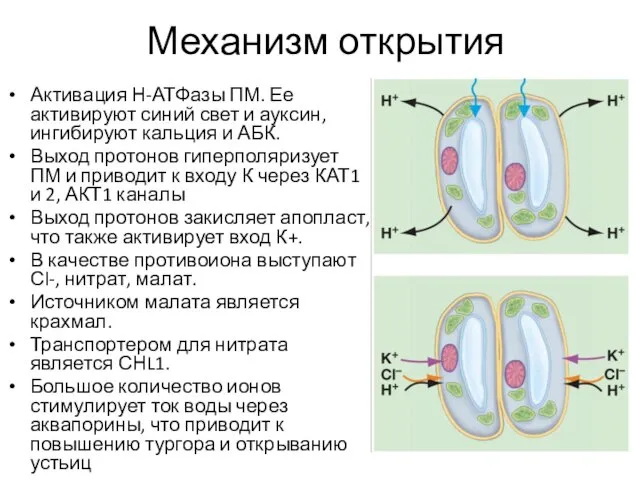
Механизм открытия
Активация Н-АТФазы ПМ. Ее активируют синий свет и ауксин, ингибируют
Механизм открытия
Активация Н-АТФазы ПМ. Ее активируют синий свет и ауксин, ингибируют

Восприятие устьицами светового сигнала
Было обнаружено, что синий свет вызывает открывание устьиц.
При
Восприятие устьицами светового сигнала
Было обнаружено, что синий свет вызывает открывание устьиц.
При

Фототропин
Фототропин – белок, содержащий 2 LOV-домена (light, oxygen, voltage), а также
Фототропин
Фототропин – белок, содержащий 2 LOV-домена (light, oxygen, voltage), а также

Механизм передачи сигнала
Ключевым элементом регуляции является Н+-АТФаза плазмалеммы.
Ее активность регулируется путем
Механизм передачи сигнала
Ключевым элементом регуляции является Н+-АТФаза плазмалеммы.
Ее активность регулируется путем
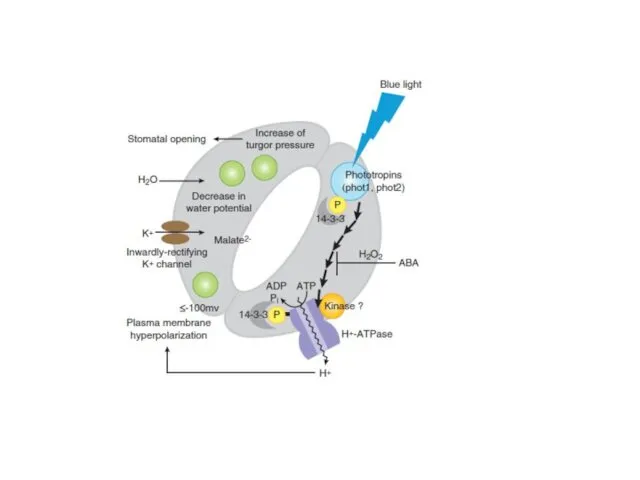
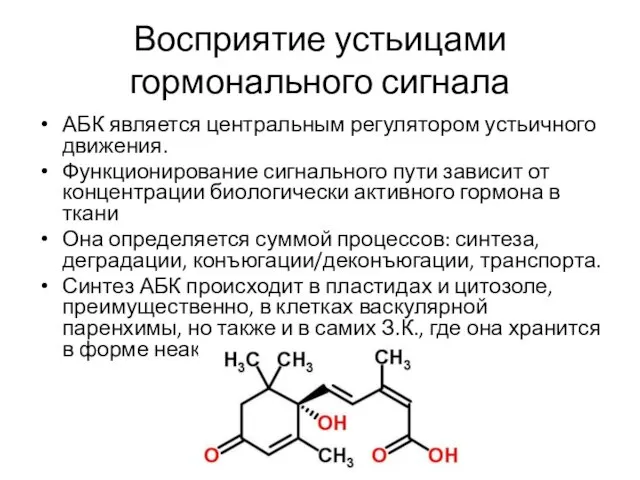
Восприятие устьицами гормонального сигнала
АБК является центральным регулятором устьичного движения.
Функционирование сигнального пути
Восприятие устьицами гормонального сигнала
АБК является центральным регулятором устьичного движения.
Функционирование сигнального пути

Транспорт АБК
Поскольку АБК может транспортироваться на дальние расстояния, она служит надежным
Транспорт АБК
Поскольку АБК может транспортироваться на дальние расстояния, она служит надежным
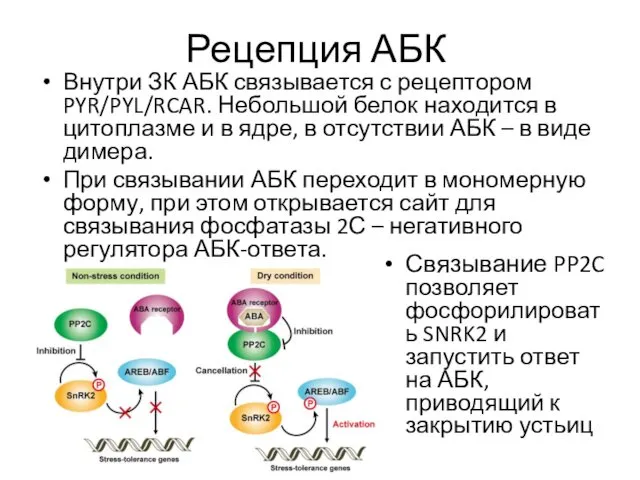
Рецепция АБК
Внутри ЗК АБК связывается с рецептором PYR/PYL/RCAR. Небольшой белок находится
Рецепция АБК
Внутри ЗК АБК связывается с рецептором PYR/PYL/RCAR. Небольшой белок находится
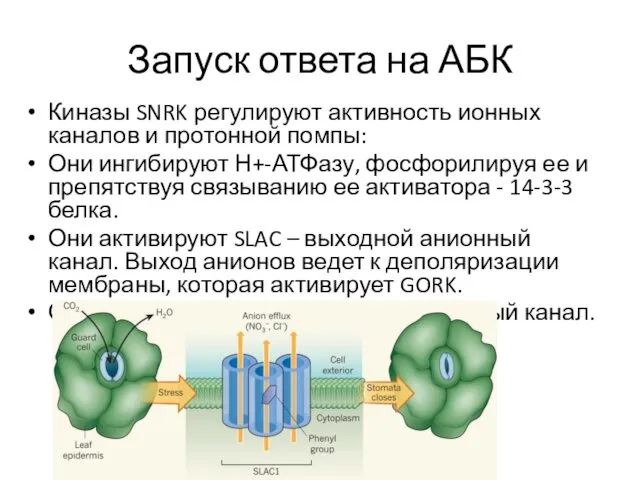
Запуск ответа на АБК
Киназы SNRK регулируют активность ионных каналов и протонной
Запуск ответа на АБК
Киназы SNRK регулируют активность ионных каналов и протонной
 Распознавание объектов на местности с помощью нейронных сетей
Распознавание объектов на местности с помощью нейронных сетей Методика подбора и анализа моделей аналогов
Методика подбора и анализа моделей аналогов Контрольна робота з теми Життя людей за первісних часів
Контрольна робота з теми Життя людей за первісних часів Основы ландшафтной колористики
Основы ландшафтной колористики Коррозия металлов и способы защиты
Коррозия металлов и способы защиты Лекция-презентация. Формирования позиции ответственного родительства.
Лекция-презентация. Формирования позиции ответственного родительства. Алкоголизм- это страшная беда современного общества
Алкоголизм- это страшная беда современного общества Жизнь и быт поволжских татар
Жизнь и быт поволжских татар История миссии Российской Православной Церкви на Американском континенте
История миссии Российской Православной Церкви на Американском континенте практические работы
практические работы Создание на территории субъекта РФ площадок для обучения и творчества в сфере информационных технологий для детей ИТ-cube
Создание на территории субъекта РФ площадок для обучения и творчества в сфере информационных технологий для детей ИТ-cube Умножение дробей
Умножение дробей Вводная презентация к проекту Я - петербуржец.
Вводная презентация к проекту Я - петербуржец. Ремонт автомобильных дорог Чайковского муниципального района. 2018 год
Ремонт автомобильных дорог Чайковского муниципального района. 2018 год Доклад для воспитателей ДОУ на тему :Роль сказок в развитии детей дошкольного возраста
Доклад для воспитателей ДОУ на тему :Роль сказок в развитии детей дошкольного возраста Қаттылықты анықтау тәсілдер
Қаттылықты анықтау тәсілдер Великая Отечественная война.
Великая Отечественная война. Правила безопасного сексуального поведения
Правила безопасного сексуального поведения Презентация Листопадничек
Презентация Листопадничек Астраханский Кремль
Астраханский Кремль Food and fertilizers. Outline
Food and fertilizers. Outline Домашняя экономика
Домашняя экономика Константин Григорьевич Паустовский (1892-1968)
Константин Григорьевич Паустовский (1892-1968) Пассажирские сервисы
Пассажирские сервисы Вращающаяся печь для обжига портландцемента
Вращающаяся печь для обжига портландцемента PEShIE_EKSKURSII_2022
PEShIE_EKSKURSII_2022 Устройства ввода и вывода звука
Устройства ввода и вывода звука Интеллектуальная игра. Где логика
Интеллектуальная игра. Где логика