- Аэробное и анаэробное дыхание прокариот

Содержание
- 2. Вопросы: 1. Аэробное дыхание. 2. Хемосинтез. Группы аэробных хемолитотрофных бактерий. 3. Группы аэробных хеморганотрофных бактерий. 4.
- 3. Дыхание – это окислительно-восстановительный процесс, идущий с образованием АТФ.
- 4. В качестве донора электронов и протонов прокариоты могут использовать: органические вещества (органотрофные бактерии) неорганические вещества (литотрофные
- 5. Аэробное дыхание - это тип дыхания, при котором конечным акцептором электронов является О2
- 6. Анаэробное дыхание - тип дыхания при котором конечным акцептором электронов могут служит: «связанный кислород» в форме
- 7. 1. Аэробное дыхание Аэробное дыхание – это процесс, обратный «нормальному» фотосинтезу, т. е. органическое вещество (СН2О)
- 8. Аэробные организмы обладают особым аппаратом: дыхательной (электронтранспортной) цепью ферментом АТФ-синтетазой. оба компонента у прокариот расположены в
- 9. Дыхательная цепь - это система переносчиков электронов и протонов от донора (окисляемого вещества) к акцептору (при
- 10. Протоны и электроны, извлеченные при окислении субстрата (при помощи дегидрогеназ), акцептируются НАД или ФАД (это переносчики
- 11. Вследствие чего возникает трансмембранный электрохимический протонный градиент, который создает протондвижущую силу. Протоны возвращаются, «перетекают», обратно с
- 12. Синтез АТФ осуществляется при помощи мембранного фермента АТФ-синтетазы путем конверсии энергии трансмембранного электрохимического градиента протонов в
- 13. 2. Хемосинтез. Группы аэробных хемолитотрофных прокариот Хемосинтез – это способность некоторых прокариот синтезировать клеточные компоненты, используя
- 14. Явление хемосинтеза было открыто С. Н. Виноградским в 1887-1890 гг. Хемосинтез осуществляют только прокариоты – хемолитоавтотрофные
- 15. Дыхательная цепь хемолитоавтотрофных бактерий
- 16. В большинстве случаев включение электронов в дыхательную цепь при окислении неорганических субстратов (NH4+, NO2‾, Н2S, S,
- 17. Для образования восстановителя НАД∙Н2 у таких бактерий работает система обратного переноса электронов, т. е. «лифт», поднимающий
- 18. Группы аэробных хемолитотрофных прокариот Нитрифицирующие бактерии Аэробные Грам(-) бактерии различной морфологии. Окисляют восстановленные соединения азота (аммоний,
- 19. Процесс нитрификации идет в два этапа: 1. Окисление аммония до нитрита: NH4+ + 1,5 O2 →
- 20. 2. Окисление нитрита (NO2 -) до нитрата: NO2‾ + ½ O2 → NO3‾ Осуществляют бактерии родов:
- 21. Между двумя группами нитрифицирующих бактерий особый тип симбиоза – метабиоз – взаимоотношения, когда один вид используют
- 22. Значение: Участвуют в круговороте азота, осуществляя нитрификацию. В хорошо аэрируемых почвах могут приводить к подкислению почвы.
- 23. Тионовые бактерии К ним относятся бактерии родов: Thiobacillus, Thiomicrospora, Thiosphаera, Thermothrix и др. Окисляют восстановленные соединения
- 24. Встречаются в водоемах при наличии Н2S и S, в серных источниках, в сульфидных и серных месторождениях,
- 25. Железобактерии Истинные железобактерии (Thiobacillus ferrooxidans, Leptospirillum ferrooxidans) окисляют Fe2+ до Fe3+ с целью получения энергии. Окисление
- 26. Железобактерии встречаются в кислых рудничных стоках, в подземных водах сульфидных месторождений, кислых железистых источниках и озерах.
- 27. Водородные бактерии Облигатные водородные бактерии входят в состав родов Сalderobacterium и Aquifex (экстремальные термофилы, развивающиеся при
- 28. Энергию получают при окислении водорода: Н2 + ½ О2 → Н2О Имеют гидрогеназы – ферменты, катализирующие
- 29. Карбоксидобактерии Род Carboxydomonas, некоторые представители р. Pseudomonas СО окисляют для получения энергии: 2СО + О2 →
- 30. 3. Аэробные хемоорганотрофные бактерии Типичные хемоорганогетеротрофы - бактерии семейства Enterobacteriaceae, например, Escherichia coli. Энтеробактерии предпочитают использовать
- 31. Окисление глюкозы происходит в гликолитическом или окислительном пентозофосфатном пути, в результате образуется ПВК, которая после окислительного
- 32. К хемоорганогетеротрофным бактериям также относятся узкоспециализированные группы: Метанотрофы и метилотрофы – окисляют метан СН4, метанол СН3ОН,
- 33. 4. Анаэробное дыхание Нитратное дыхание Конечным акцептором электронов является нитрат NO3-, который восстанавливается до нитрита NO2-:
- 34. Сульфатное дыхание Конечным акцептором электронов является сульфат SO42-, который восстанавливается через сульфит (SO32 ‾) до сероводорода:
- 35. Серное дыхание В качестве конечного акцептора электронов используется молекулярная сера S, которая восстанавливается до сероводорода. Характерно
- 36. Карбонатное дыхание Конечным акцептором электронов является СО2. Характерно для анаэробных ацетогенных бактерий (р. Acetobacterium и др.),
- 37. У метанобразующих бактерий в процессе карбонатного дыхания при окислении Н2 образуется метан СН4: 4Н2 + СО2
- 39. Скачать презентацию

Вопросы:
1. Аэробное дыхание.
2. Хемосинтез. Группы аэробных хемолитотрофных бактерий.
3. Группы аэробных
Вопросы:
1. Аэробное дыхание.
2. Хемосинтез. Группы аэробных хемолитотрофных бактерий.
3. Группы аэробных

Дыхание – это окислительно-восстановительный процесс, идущий с образованием АТФ.
Дыхание – это окислительно-восстановительный процесс, идущий с образованием АТФ.

В качестве донора электронов и протонов прокариоты могут использовать:
органические вещества (органотрофные
В качестве донора электронов и протонов прокариоты могут использовать:
органические вещества (органотрофные

Аэробное дыхание - это тип дыхания, при котором конечным акцептором электронов
Аэробное дыхание - это тип дыхания, при котором конечным акцептором электронов

Анаэробное дыхание - тип дыхания при котором конечным акцептором электронов могут
Анаэробное дыхание - тип дыхания при котором конечным акцептором электронов могут

1. Аэробное дыхание
Аэробное дыхание – это процесс, обратный «нормальному» фотосинтезу,
1. Аэробное дыхание
Аэробное дыхание – это процесс, обратный «нормальному» фотосинтезу,

Аэробные организмы обладают особым аппаратом:
дыхательной (электронтранспортной) цепью
ферментом АТФ-синтетазой.
оба
Аэробные организмы обладают особым аппаратом:
дыхательной (электронтранспортной) цепью
ферментом АТФ-синтетазой.
оба

Дыхательная цепь - это система переносчиков электронов и протонов от донора
Дыхательная цепь - это система переносчиков электронов и протонов от донора

Протоны и электроны, извлеченные при окислении субстрата (при помощи дегидрогеназ), акцептируются
Протоны и электроны, извлеченные при окислении субстрата (при помощи дегидрогеназ), акцептируются

Вследствие чего возникает трансмембранный электрохимический протонный градиент, который создает протондвижущую силу.
Вследствие чего возникает трансмембранный электрохимический протонный градиент, который создает протондвижущую силу.

Синтез АТФ осуществляется при помощи мембранного фермента АТФ-синтетазы путем конверсии энергии
Синтез АТФ осуществляется при помощи мембранного фермента АТФ-синтетазы путем конверсии энергии

2. Хемосинтез. Группы аэробных хемолитотрофных прокариот
Хемосинтез – это способность некоторых прокариот
2. Хемосинтез. Группы аэробных хемолитотрофных прокариот
Хемосинтез – это способность некоторых прокариот

Явление хемосинтеза было открыто С. Н. Виноградским в 1887-1890 гг.
Хемосинтез
Явление хемосинтеза было открыто С. Н. Виноградским в 1887-1890 гг.
Хемосинтез
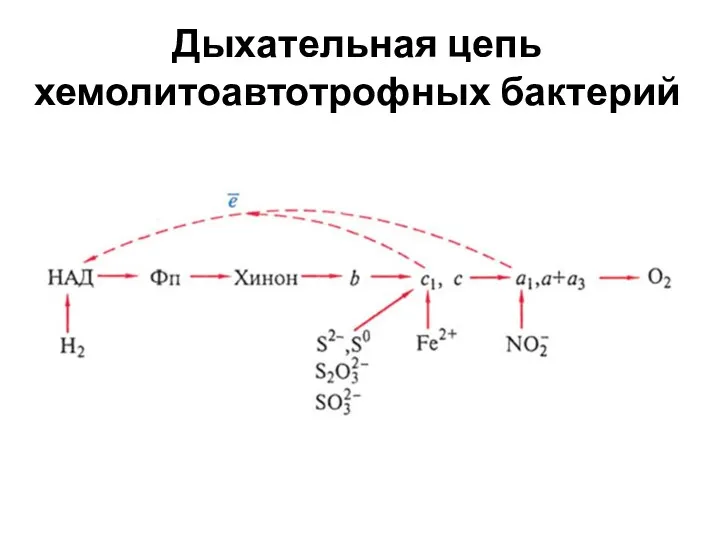
Дыхательная цепь хемолитоавтотрофных бактерий
Дыхательная цепь хемолитоавтотрофных бактерий

В большинстве случаев включение электронов в дыхательную цепь при окислении неорганических
В большинстве случаев включение электронов в дыхательную цепь при окислении неорганических

Для образования восстановителя НАД∙Н2 у таких бактерий работает система обратного переноса
Для образования восстановителя НАД∙Н2 у таких бактерий работает система обратного переноса

Группы аэробных хемолитотрофных прокариот
Нитрифицирующие бактерии
Аэробные Грам(-) бактерии различной морфологии.
Окисляют восстановленные
Группы аэробных хемолитотрофных прокариот
Нитрифицирующие бактерии
Аэробные Грам(-) бактерии различной морфологии.
Окисляют восстановленные

Процесс нитрификации идет в два этапа:
1. Окисление аммония до нитрита:
NH4+ +
Процесс нитрификации идет в два этапа:
1. Окисление аммония до нитрита:
NH4+ +

2. Окисление нитрита (NO2 -) до нитрата:
NO2‾ + ½ O2
2. Окисление нитрита (NO2 -) до нитрата:
NO2‾ + ½ O2

Между двумя группами нитрифицирующих бактерий особый тип симбиоза – метабиоз –
Между двумя группами нитрифицирующих бактерий особый тип симбиоза – метабиоз –

Значение:
Участвуют в круговороте азота, осуществляя нитрификацию.
В хорошо аэрируемых почвах
Значение:
Участвуют в круговороте азота, осуществляя нитрификацию.
В хорошо аэрируемых почвах

Тионовые бактерии
К ним относятся бактерии родов: Thiobacillus, Thiomicrospora, Thiosphаera, Thermothrix и
Тионовые бактерии
К ним относятся бактерии родов: Thiobacillus, Thiomicrospora, Thiosphаera, Thermothrix и

Встречаются в водоемах при наличии Н2S и S, в серных источниках,
Встречаются в водоемах при наличии Н2S и S, в серных источниках,

Железобактерии
Истинные железобактерии (Thiobacillus ferrooxidans, Leptospirillum ferrooxidans) окисляют Fe2+ до Fe3+ с
Железобактерии
Истинные железобактерии (Thiobacillus ferrooxidans, Leptospirillum ferrooxidans) окисляют Fe2+ до Fe3+ с

Железобактерии встречаются в кислых рудничных стоках, в подземных водах сульфидных месторождений,
Железобактерии встречаются в кислых рудничных стоках, в подземных водах сульфидных месторождений,

Водородные бактерии
Облигатные водородные бактерии входят в состав родов Сalderobacterium и
Водородные бактерии
Облигатные водородные бактерии входят в состав родов Сalderobacterium и

Энергию получают при окислении водорода:
Н2 + ½ О2 → Н2О
Имеют гидрогеназы
Энергию получают при окислении водорода:
Н2 + ½ О2 → Н2О
Имеют гидрогеназы

Карбоксидобактерии
Род Carboxydomonas, некоторые представители р. Pseudomonas
СО окисляют для получения энергии:
Карбоксидобактерии
Род Carboxydomonas, некоторые представители р. Pseudomonas
СО окисляют для получения энергии:

3. Аэробные хемоорганотрофные бактерии
Типичные хемоорганогетеротрофы - бактерии семейства Enterobacteriaceae, например, Escherichia
3. Аэробные хемоорганотрофные бактерии
Типичные хемоорганогетеротрофы - бактерии семейства Enterobacteriaceae, например, Escherichia

Окисление глюкозы происходит в гликолитическом или окислительном пентозофосфатном пути, в результате
Окисление глюкозы происходит в гликолитическом или окислительном пентозофосфатном пути, в результате

К хемоорганогетеротрофным бактериям также относятся узкоспециализированные группы:
Метанотрофы и метилотрофы – окисляют
К хемоорганогетеротрофным бактериям также относятся узкоспециализированные группы:
Метанотрофы и метилотрофы – окисляют

4. Анаэробное дыхание
Нитратное дыхание
Конечным акцептором электронов является нитрат NO3-, который восстанавливается
4. Анаэробное дыхание
Нитратное дыхание
Конечным акцептором электронов является нитрат NO3-, который восстанавливается

Сульфатное дыхание
Конечным акцептором электронов является сульфат SO42-, который восстанавливается через сульфит
Сульфатное дыхание
Конечным акцептором электронов является сульфат SO42-, который восстанавливается через сульфит

Серное дыхание
В качестве конечного акцептора электронов используется молекулярная сера S, которая
Серное дыхание
В качестве конечного акцептора электронов используется молекулярная сера S, которая

Карбонатное дыхание
Конечным акцептором электронов является СО2.
Характерно для анаэробных ацетогенных бактерий
Карбонатное дыхание
Конечным акцептором электронов является СО2.
Характерно для анаэробных ацетогенных бактерий

У метанобразующих бактерий в процессе карбонатного дыхания при окислении Н2 образуется
У метанобразующих бактерий в процессе карбонатного дыхания при окислении Н2 образуется
 Тип Кишечнополостные. Тип Гребневики. Жизненные циклы кишечнополостных
Тип Кишечнополостные. Тип Гребневики. Жизненные циклы кишечнополостных The endangered animals
The endangered animals Пищеварение в ротовой полости и желудке
Пищеварение в ротовой полости и желудке Постельные клопы
Постельные клопы Мочеполовой аппарат. Органы мочеотделения. (Лекция 5)
Мочеполовой аппарат. Органы мочеотделения. (Лекция 5) Педсовет Современные подходы к преподаванию в условиях ФГОС
Педсовет Современные подходы к преподаванию в условиях ФГОС Размножение организмов. (Лекция 5)
Размножение организмов. (Лекция 5) Оснащение микробиологической лаборатории
Оснащение микробиологической лаборатории Кора большого мозга
Кора большого мозга Венозная система. Лекция № 19
Венозная система. Лекция № 19 Лабораторная диагностика нарушений обмена белков плазмы крови (Преальбумин, С-РБ, Фибронектин, Фибриноген)
Лабораторная диагностика нарушений обмена белков плазмы крови (Преальбумин, С-РБ, Фибронектин, Фибриноген) Теории происхождения жизни. (Лекция 15)
Теории происхождения жизни. (Лекция 15) Становление наук о человеке
Становление наук о человеке Рябина обыкновенная
Рябина обыкновенная Строение сердечно-сосудистой системы
Строение сердечно-сосудистой системы Қызыл балдырлардың түрлерінің жалпы
Қызыл балдырлардың түрлерінің жалпы Различия в строении клеток эукариот и прокариот
Различия в строении клеток эукариот и прокариот Объекты дендрологии. Понятие о жизненной форме у древесных растений
Объекты дендрологии. Понятие о жизненной форме у древесных растений Мышцы туловища, головы, шеи
Мышцы туловища, головы, шеи презентация Орган слуха
презентация Орган слуха Різноманітність ссавців
Різноманітність ссавців Высшие жирные кислоты. Липиды
Высшие жирные кислоты. Липиды Вирусы, вирусология.
Вирусы, вирусология. Обмін білків
Обмін білків Симпатический отдел автономной нервной системы
Симпатический отдел автономной нервной системы Биотические связи
Биотические связи Углеводы. 9 класс
Углеводы. 9 класс Строение клетки растения и животного
Строение клетки растения и животного