- AKAPs. Общий признак всех AKAPs

Содержание
- 2. Общий признак всех AKAPs – домен связывания RI/II субъединиц сАК. Тканевая экспрессия: AKAP75/AKAP79/AKAP150 в плазматической мембране
- 3. Schematic view of AKAP250 C.C. Malbon, J. Tao, E. Shumay and H.-Y. Wang Biochemical Society Transactions
- 4. Carmen W. Dessauer Molecular Pharmacology 2009 Изоформы аденилатциклазы и AKAPs с которыми они образуют комплексы smallest
- 6. Berridge, M. J. 1984. Inositol triphosphate and diacylglycerol as second messengers. Biochem. J. 220:345-360. Ca2+ controlled
- 7. Cleavage of PIP2, catalyzed by Phospholipase C, yields 2 second messengers: inositol-1,4,5-trisphosphate (IP3) diacylglycerol (DG). Diacylglycerol,
- 8. Структура кальцисомы
- 9. Hypertonicity stimulates UT-A1 phosphorylation, urea permeability through protein kinase C (PKC) and intracellular calcium. PKC-α is
- 10. classical PKCs (PKCα, βI, βII, and γ) are regulated by both Ca2+ and by diacylglycerol (DAG)
- 11. Domain structure of PKC
- 12. osmosis is
- 13. Закон Фика Движущая сила при диффузии Осмос, осмотическое давление
- 14. Основные механизмы трансмембранного транспорта, обеспечивающие регуляторное снижение объема клетки (RVD) (Hoffmann et al., 2009).
- 15. Fi - сила(поле); Pi - потенциал скорость дрейфа пропорциональна движущей силе B – подвижность единичный поток
- 16. URT=D молекулярный перенос Закон Фика Химический потенциал для электронейтральных частиц где градиент потенциала - движущая сила
- 17. Осмотический поток воды: (Вант-Гофф )
- 18. осмотический перенос van't Hoff equation: П = iMRT где П (pi) осмотическое давление (kPa), M молярная
- 20. = μ0 + RTlnC + ZFϕ электрохимический потенциал Z – заряд, F - число Фарадея, ϕ
- 21. The Nobel Prize in Chemistry 2003
- 22. AQP0 AQP1 AqpZ AQP0
- 25. A schematic for structure of AQP-2 Conservative NPA sequences are illustrated as open boxes Potential N-glycosylation
- 27. AQP0 AQP1
- 33. “Luckily, physiological mechanisms allow animals to escape. . . .”
- 35. Y. Noda and S. Sasaki 2005
- 36. Jason D. Hoffert, Chung-Lin Chou, and Mark A. Knepper 2009
- 37. Ca+2/calmodulin plays a critical role in regulation of renal collecting duct water permeability by vasopressin Vasopressin-stimulated
- 38. Kuniaki Takata · Toshiyuki Matsuzaki · Yuki Tajika · Abduxukur Ablimit · Takahiro Hasegawa; Histochem Cell
- 39. в почках везикулы содержащие AQP2 для встраивания требуют SNARE (The soluble N-ethylmaleimide-sensitive factor attachment protein target
- 40. MSc Faculty of Natural Sciences Novosibirsk State University Novosibirsk, Russia PhD Institute of Cytology and Genetics
- 41. Lecturer at Cell Physics Department of Applied Physics School of Engineering Sciences Royal Institute of Technology
- 42. M.D. Professor Tokyo Medical & Dental University
- 47. Равновесие достигается когда соотношение вероятностей нахождения ионов («концентрация» в макро системе) в области высокой энергии –
- 48. Поток ионов через каналы: (Goldman 1943; Hodgkin and Katz 1949)
- 49. Электрохимический потенциал иона Х Где: - стандартный химический потенциал иона Х Z – валентность иона Х
- 52. Goldman-Hodgkin-Katz
- 53. Jens Christian Skou Born October 8, 1918 ) (age 91) Nationality Danish Aarhus University 1997, Nobel
- 56. ионные каналы обладают селективностью и периодически меняют состояние открыто-закрыто
- 57. selectivity filter
- 58. a, TRPC1 TRPC5 heteromer; b, TRPM5; c, TRPV5, TRPV6
- 59. Образование клатриновых везикул, содержащих комплекс рецептора V2 с вазопрессином, повидимому, одно из звеньев механизма действия гормона.
- 60. Десенситизация рецептора - фосфорилирование GRK ( G-protein Receptor Kinase). - связывание с b-arrestin и механизмом эндоцитоза
- 61. GRKs/arrestins – роль в десенситизации GPCRs
- 62. Demonstration of VP receptor endocytosis in LLC-PK1a cells expressing V2R–GFP, visualized by spinning disc confocal microscopy
- 63. Confocal images showing localization of AQP2 (red) and V2R–GFP (green) in transfected cells expressing both Proteins
- 64. Различные функции и механизмы эндоцитоза
- 66. В настоящее время нет однозначного ответа на вопрос, что служит стимулом к началу формирования клатриновых везикул,
- 67. Эндоцитоз GPCRрецепторов связан с фосфорилированием их C-концевой области рядом киназ. Известно, что GPCR фосфорилируются такими киназами,
- 68. Образование везикулы, окаймленной клатрином, происходит благодаря формированию связанного с плазматической мембраной белкового комплекса эндоцитоз рецепторов, сопряженных
- 69. Эндоцитоз начинается с формирования изгиба мембраны, направленной внутрь клетки. Значительную роль в создании и стабилизации кривизны
- 70. BAR_белки образует суперсемейство, в котором выделяют белки : BAR/N_BAR, проявляющие сродство к мембранам с высокой положительной
- 71. BAR домены формируют гомодимер (BAR модуль) серповидной формы, состоящий из двух суперспирализованных мономеров, при этом угол,
- 72. amphiphysin (BAR) Uncoating to produce naked CCV Endocytosis of V2R
- 73. формирование инвагинации путем внедрения в мембрану амфипатической спирали
- 74. адаптерный комплекс AP2 AP2 – это узловой белок формирования динамичной сети низкоаффинных взаимодействий. AP2 инициирует сборку
- 75. формирование изгиба мембраны Marcus J. Taylor1, David Perrais2,3, Christien J. Merrifield, PLoS Biology 2011 H. T.
- 77. отделение клатриновой везикулы от плазматической мембраны. Полимеризация клатрина способствует искривлению мембраны и приводит к вытеснению некоторых
- 78. в регуляцию кривизны мембраны вовлечен актиновый цитоскелет. На высокоразрешающих электронных изображениях можно увидеть формирование актинового комплекса
- 79. Везикулы отщепляются главным образом динамином. В клетке динамин формирует спираль вокруг шейки образующейся везикулы, что при
- 80. Динамин – белок размером порядка 100 кДа, обладающий GTPазной активностью и способный образовывать спиральные олигомеры, размер
- 81. Praefcke, G.J.K. and McMahon, H.T. (2004) 2013 г. А. В. Иляскин и др.
- 83. Последний этап клатринзависимого эндоцитоза – разрушение клатриновой оболочки ATP азой Hsc70, ее кофактром ауксилином или циклин
- 85. Интернализованные везикулы сливаются с ранними эндосомами , затем карго-белок рециклизуется в мембрану или направляется в лизосомы.
- 86. пять основных стадий формирования клатриновой везикулы: инициация, селекция интернализуемого белка, сборка клатриновой оболочки, отщепление везикулы разрушение
- 88. Cellular fate of V2R mutants in nephrogenic diabetes insipidus (NDI) and their rescue.
- 91. PTEN (название гена) (“phosphatase and tensin homolog”) семейство тирозиновых фосфатаз. Участвует в торможении клеточного деления, активатор
- 92. PH domain: pleckstrin homology domain; C2 domain: домен связи с мембраной; PDZ: a common structural domain
- 93. phosphatidylinositol-3 kinase (PI3K) участвует в регуляции клеточного роста и метаболизма. Активные мутантные формы PI3K участвуют в
- 94. PIP3 (продукт PI3K) связывается с АКТ , что приводит к перемещению на мембрану, где образуется связь
- 95. Суперсемейство EF-hand белков CaM кальмодулин MW 16,700 ; 148 aa; Kd = 4 – 18 10-6
- 97. The soluble N-ethylmaleimide-sensitive factor attachment protein target receptor (SNARE) предполагается, участвуют в присоединении/слиянии пузырьков на всех
- 98. (synaptosome-associated protein) SNAP23 В почке его находят в апикальной мембране и везикулах AQP2 главных клеток. участвует
- 101. Effect of VP treatment on Lysotracker and V2R-GFP distribution in LLC-V2R-GFP cells. At 37°C, Lysotracker, a
- 103. Скачать презентацию

Общий признак всех AKAPs – домен связывания RI/II субъединиц сАК.
Тканевая
Общий признак всех AKAPs – домен связывания RI/II субъединиц сАК.
Тканевая
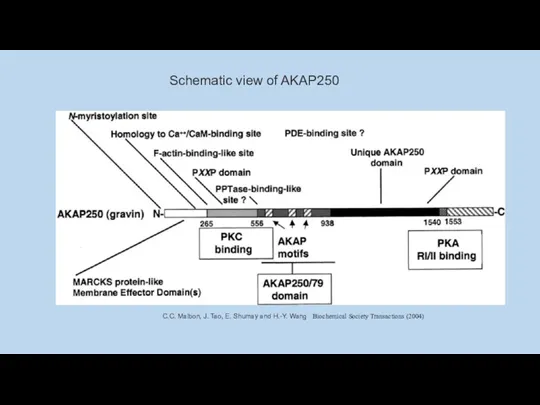
Schematic view of AKAP250
C.C. Malbon, J. Tao, E. Shumay and H.-Y.
Schematic view of AKAP250
C.C. Malbon, J. Tao, E. Shumay and H.-Y.
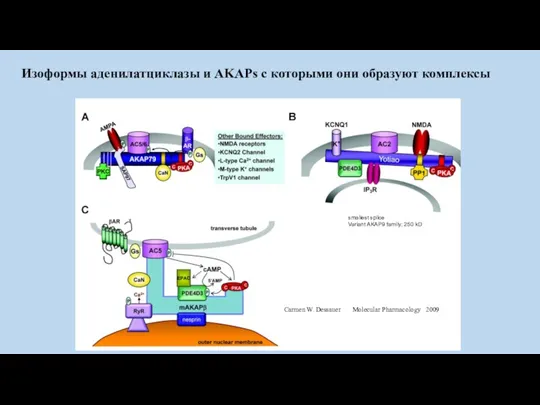
Carmen W. Dessauer Molecular Pharmacology 2009
Изоформы аденилатциклазы и AKAPs с которыми
Carmen W. Dessauer Molecular Pharmacology 2009
Изоформы аденилатциклазы и AKAPs с которыми

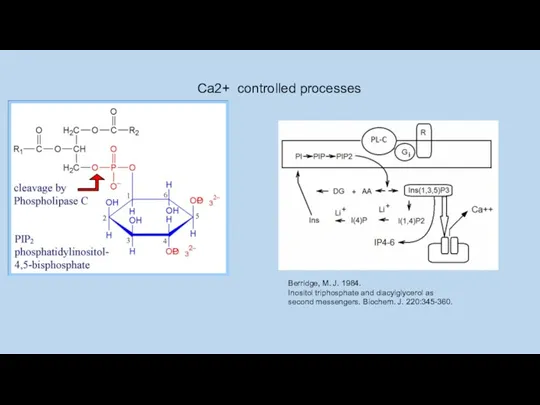
Berridge, M. J. 1984.
Inositol triphosphate and diacylglycerol as
second messengers. Biochem.
Berridge, M. J. 1984.
Inositol triphosphate and diacylglycerol as
second messengers. Biochem.
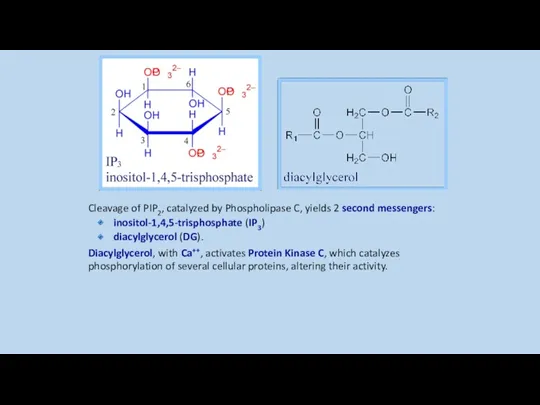
Cleavage of PIP2, catalyzed by Phospholipase C, yields 2 second messengers:
Cleavage of PIP2, catalyzed by Phospholipase C, yields 2 second messengers:
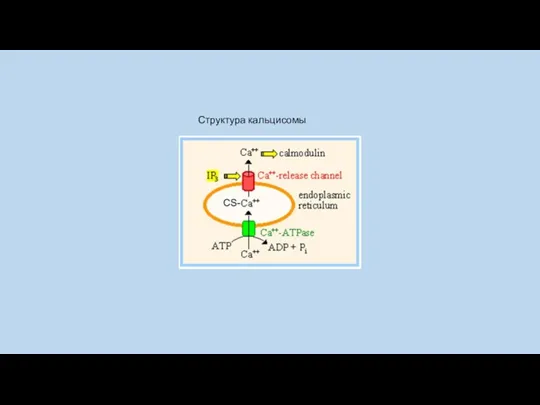
Структура кальцисомы
Структура кальцисомы

Hypertonicity stimulates UT-A1 phosphorylation, urea permeability through protein kinase C (PKC)

classical PKCs (PKCα, βI, βII, and γ)
are regulated by both
classical PKCs (PKCα, βI, βII, and γ)
are regulated by both

Domain structure of PKC
Domain structure of PKC

osmosis is
osmosis is

Закон Фика
Движущая сила при диффузии
Осмос, осмотическое давление
Закон Фика
Движущая сила при диффузии
Осмос, осмотическое давление
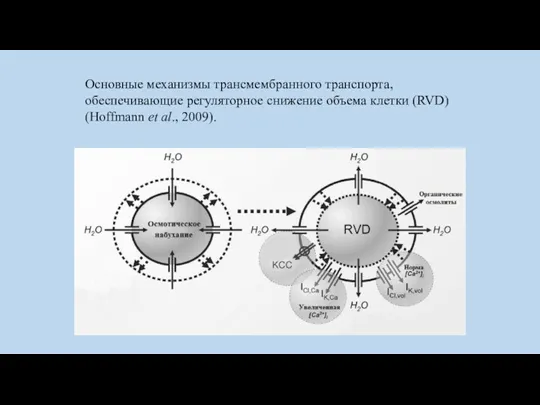
Основные механизмы трансмембранного транспорта, обеспечивающие регуляторное снижение объема клетки (RVD) (Hoffmann
Основные механизмы трансмембранного транспорта, обеспечивающие регуляторное снижение объема клетки (RVD) (Hoffmann

Fi - сила(поле); Pi - потенциал
скорость дрейфа пропорциональна движущей силе
B –
Fi - сила(поле); Pi - потенциал
скорость дрейфа пропорциональна движущей силе
B –

URT=D
молекулярный перенос
Закон Фика
Химический потенциал для электронейтральных частиц
где
градиент потенциала -
URT=D
молекулярный перенос
Закон Фика
Химический потенциал для электронейтральных частиц
где
градиент потенциала -

Осмотический поток воды:
(Вант-Гофф )
Осмотический поток воды:
(Вант-Гофф )

осмотический перенос
van't Hoff equation:
П = iMRT
где П (pi) осмотическое
осмотический перенос
van't Hoff equation:
П = iMRT
где П (pi) осмотическое

= μ0 + RTlnC + ZFϕ электрохимический потенциал
Z – заряд,
= μ0 + RTlnC + ZFϕ электрохимический потенциал
Z – заряд,
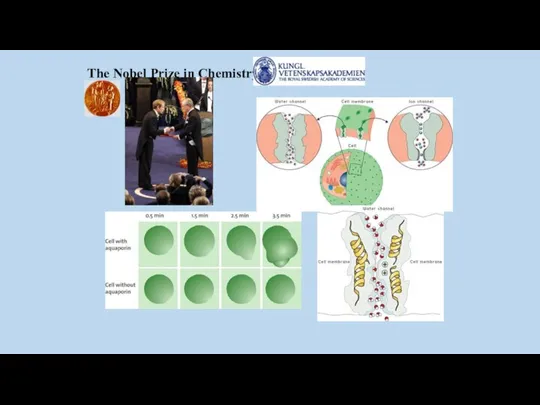
The Nobel Prize in Chemistry 2003
The Nobel Prize in Chemistry 2003

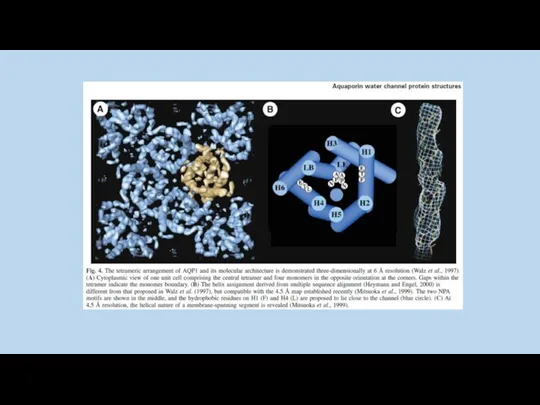
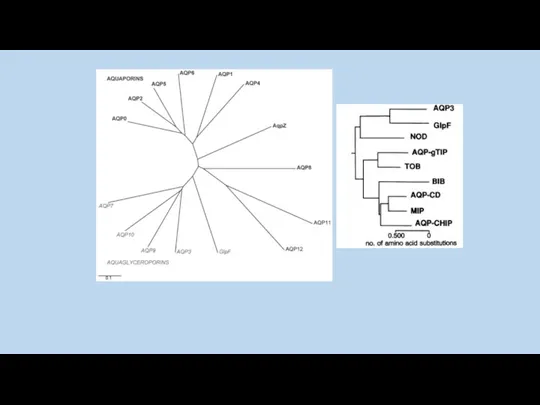
AQP0
AQP1
AqpZ
AQP0
AQP0
AQP1
AqpZ
AQP0

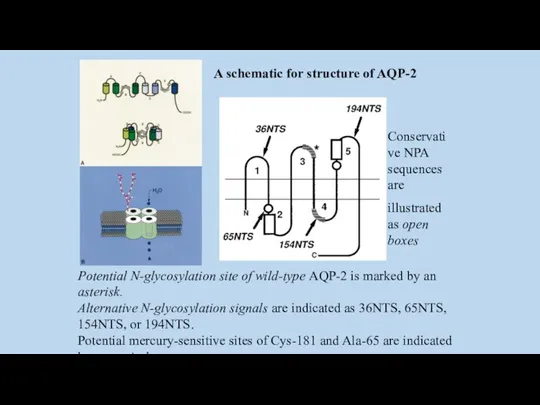
A schematic for structure of AQP-2
Conservative NPA sequences are
illustrated as open
A schematic for structure of AQP-2
Conservative NPA sequences are
illustrated as open
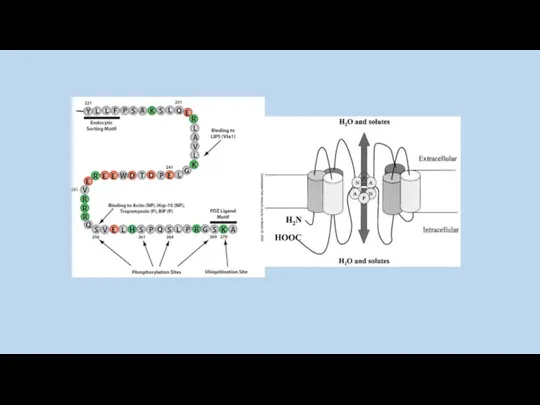
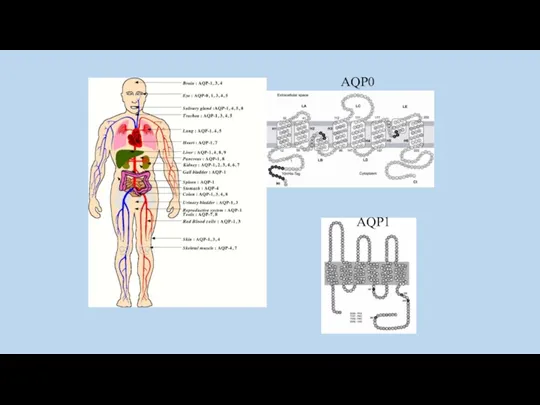
AQP0
AQP1
AQP0
AQP1
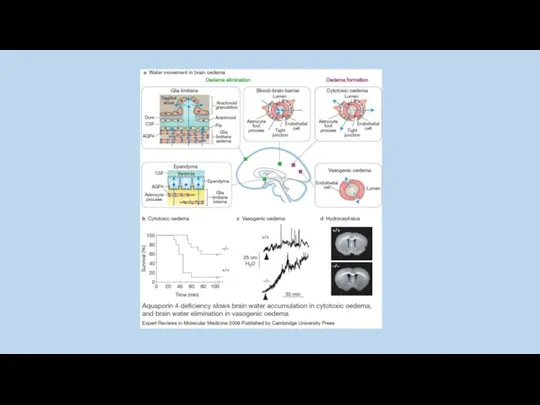
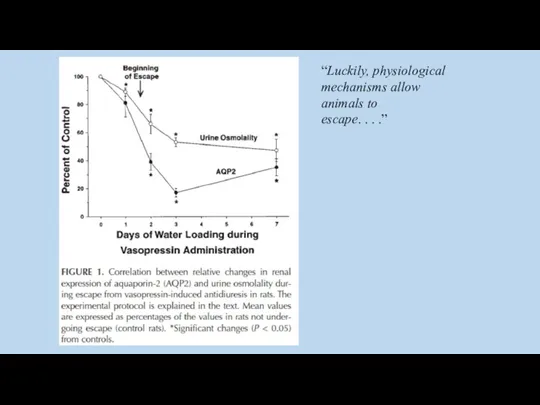
“Luckily, physiological
mechanisms allow
animals to
escape. . . .”
“Luckily, physiological
mechanisms allow
animals to
escape. . . .”
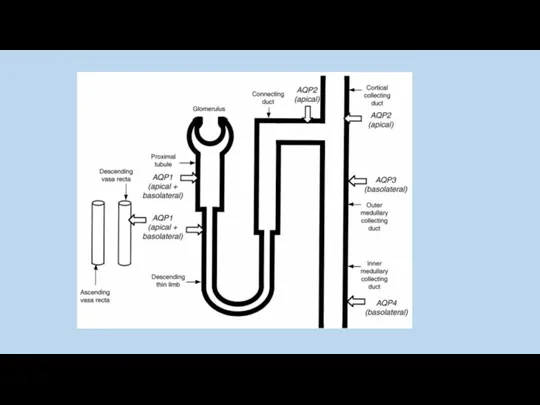

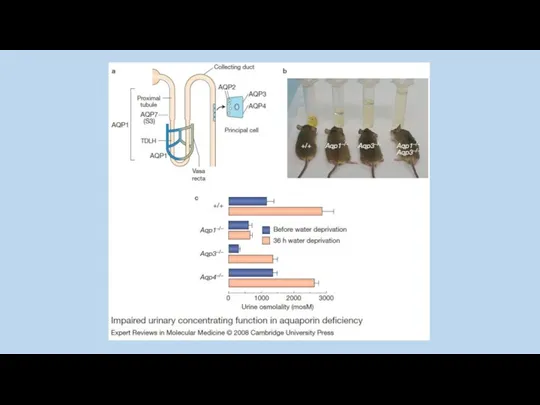
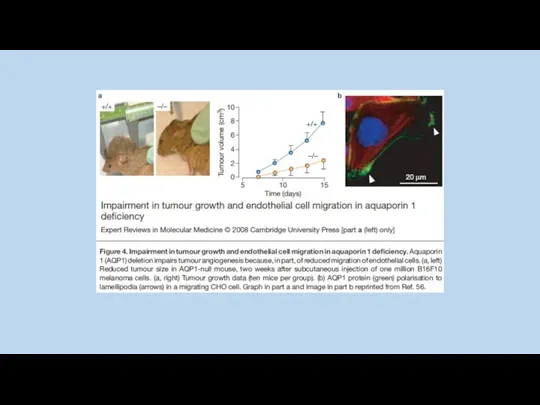
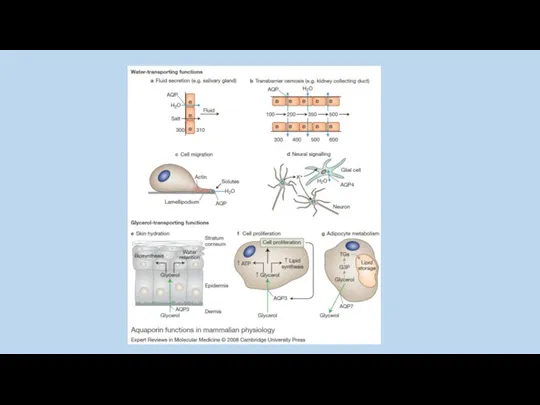
Y. Noda and S. Sasaki 2005
Y. Noda and S. Sasaki 2005
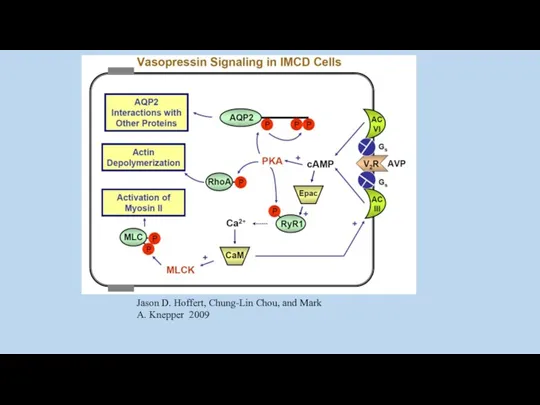
Jason D. Hoffert, Chung-Lin Chou, and Mark A. Knepper 2009
Jason D. Hoffert, Chung-Lin Chou, and Mark A. Knepper 2009
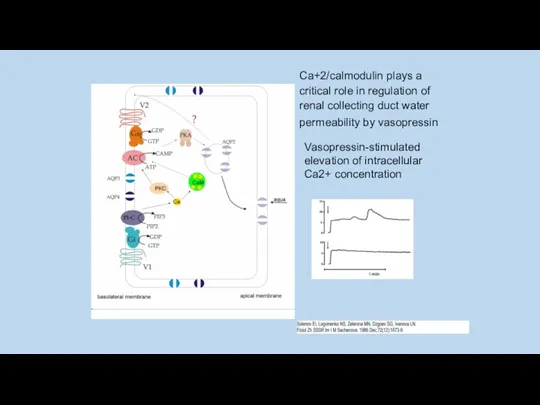
Ca+2/calmodulin plays a critical role in regulation of renal collecting duct
Ca+2/calmodulin plays a critical role in regulation of renal collecting duct
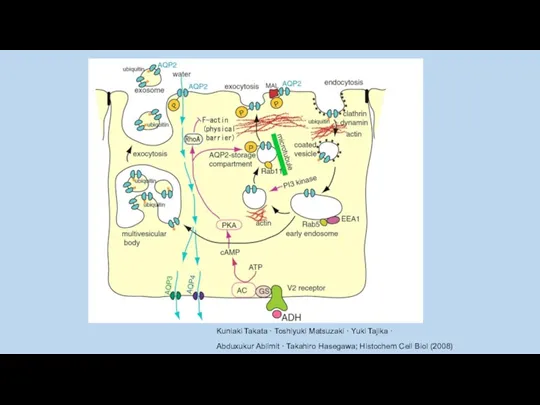
Kuniaki Takata · Toshiyuki Matsuzaki · Yuki Tajika ·
Abduxukur Ablimit ·
Kuniaki Takata · Toshiyuki Matsuzaki · Yuki Tajika ·
Abduxukur Ablimit ·

в почках везикулы содержащие AQP2 для встраивания требуют SNARE (The soluble
в почках везикулы содержащие AQP2 для встраивания требуют SNARE (The soluble

MSc
Faculty of Natural Sciences
Novosibirsk State University
Novosibirsk, Russia
PhD
Institute of Cytology and
MSc
Faculty of Natural Sciences
Novosibirsk State University
Novosibirsk, Russia
PhD
Institute of Cytology and
Lecturer at Cell Physics
Department of Applied Physics
School of Engineering Sciences
Royal Institute
Lecturer at Cell Physics
Department of Applied Physics
School of Engineering Sciences
Royal Institute

M.D. Professor
Tokyo Medical & Dental University
M.D. Professor
Tokyo Medical & Dental University



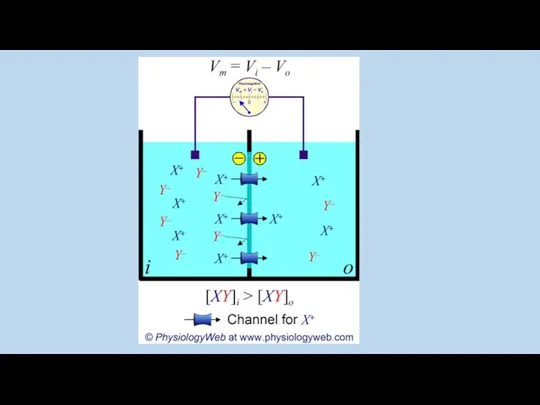

Равновесие достигается когда соотношение вероятностей нахождения ионов («концентрация» в макро системе)
Равновесие достигается когда соотношение вероятностей нахождения ионов («концентрация» в макро системе)

Поток ионов через каналы:
(Goldman 1943; Hodgkin and Katz 1949)
Поток ионов через каналы:
(Goldman 1943; Hodgkin and Katz 1949)

Электрохимический потенциал иона Х
Где:
- стандартный химический потенциал иона Х
Электрохимический потенциал иона Х
Где:
- стандартный химический потенциал иона Х



Goldman-Hodgkin-Katz
Goldman-Hodgkin-Katz
Jens Christian Skou
Born October 8, 1918 ) (age 91)
Nationality Danish
Aarhus University
Jens Christian Skou
Born October 8, 1918 ) (age 91)
Nationality Danish
Aarhus University
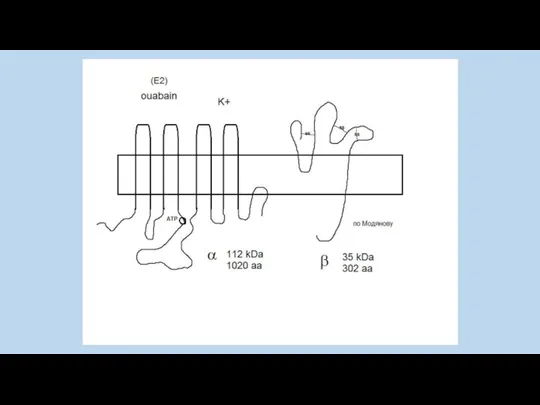
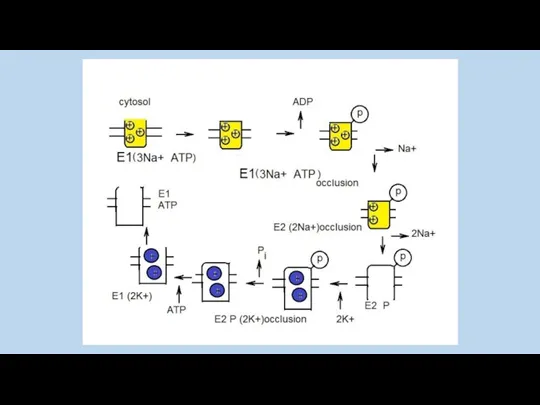
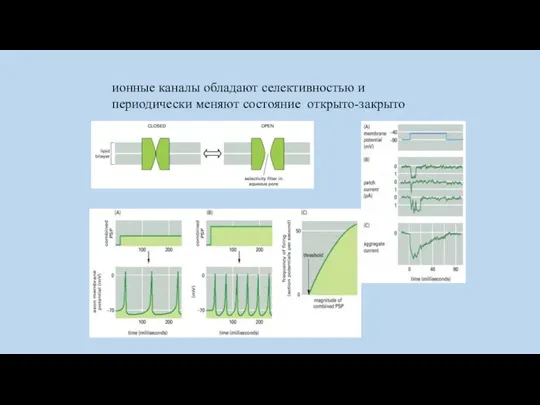
ионные каналы обладают селективностью и периодически меняют состояние открыто-закрыто
ионные каналы обладают селективностью и периодически меняют состояние открыто-закрыто
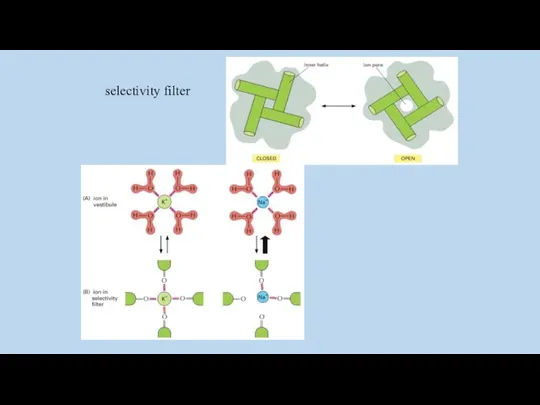
selectivity filter
selectivity filter
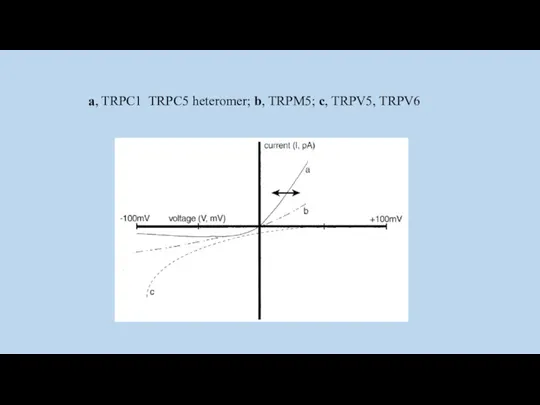
a, TRPC1 TRPC5 heteromer; b, TRPM5; c, TRPV5, TRPV6
a, TRPC1 TRPC5 heteromer; b, TRPM5; c, TRPV5, TRPV6

Образование клатриновых везикул, содержащих комплекс рецептора V2 с вазопрессином, повидимому, одно
Образование клатриновых везикул, содержащих комплекс рецептора V2 с вазопрессином, повидимому, одно

Десенситизация рецептора
- фосфорилирование GRK ( G-protein Receptor Kinase).
- связывание с
Десенситизация рецептора
- фосфорилирование GRK ( G-protein Receptor Kinase).
- связывание с
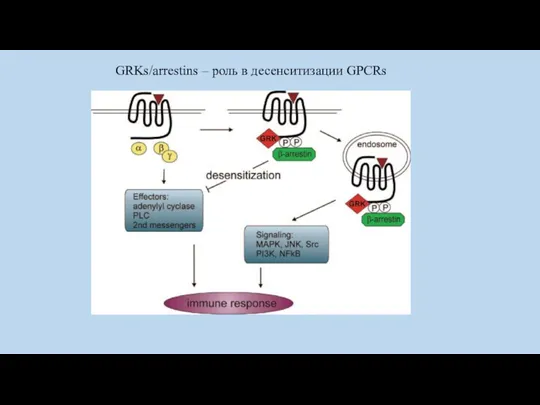
GRKs/arrestins – роль в десенситизации GPCRs
GRKs/arrestins – роль в десенситизации GPCRs
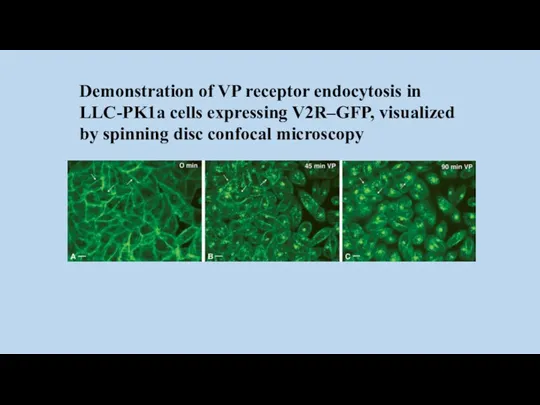
Demonstration of VP receptor endocytosis in LLC-PK1a cells expressing V2R–GFP, visualized
Demonstration of VP receptor endocytosis in LLC-PK1a cells expressing V2R–GFP, visualized
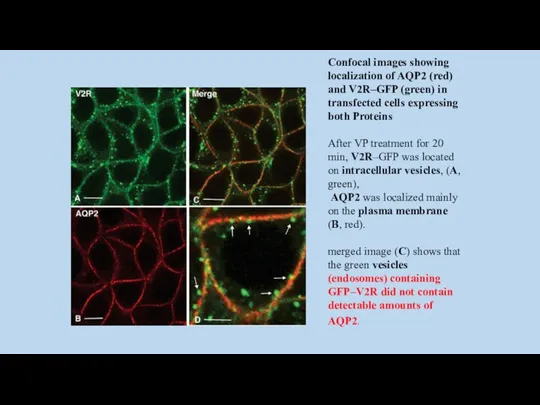
Confocal images showing localization of AQP2 (red) and V2R–GFP (green) in
Confocal images showing localization of AQP2 (red) and V2R–GFP (green) in
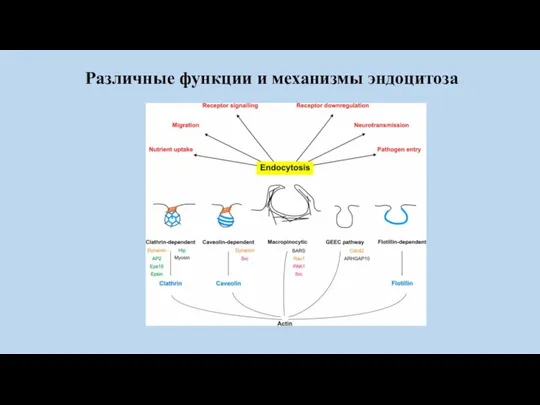
Различные функции и механизмы эндоцитоза
Различные функции и механизмы эндоцитоза
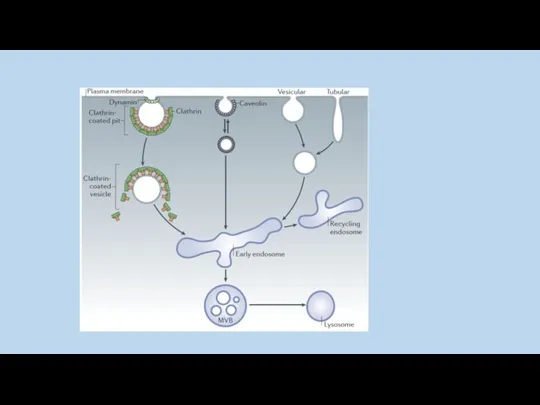

В настоящее время нет однозначного ответа на вопрос, что служит стимулом
В настоящее время нет однозначного ответа на вопрос, что служит стимулом
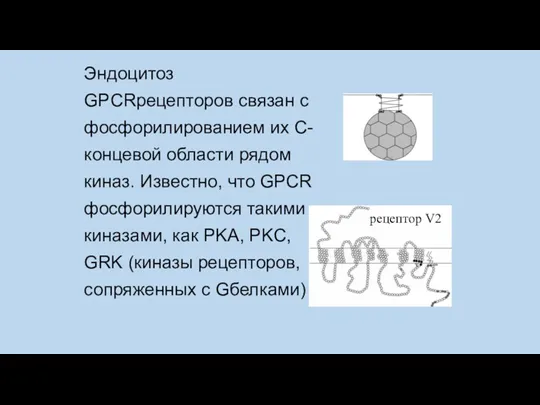
Эндоцитоз GPCRрецепторов связан с фосфорилированием их C-концевой области рядом киназ. Известно,
Эндоцитоз GPCRрецепторов связан с фосфорилированием их C-концевой области рядом киназ. Известно,

Образование везикулы, окаймленной клатрином, происходит благодаря формированию связанного с плазматической мембраной
Образование везикулы, окаймленной клатрином, происходит благодаря формированию связанного с плазматической мембраной

Эндоцитоз начинается с формирования изгиба мембраны, направленной внутрь клетки. Значительную роль
Эндоцитоз начинается с формирования изгиба мембраны, направленной внутрь клетки. Значительную роль

BAR_белки образует суперсемейство, в котором выделяют белки :
BAR/N_BAR, проявляющие сродство к
BAR_белки образует суперсемейство, в котором выделяют белки :
BAR/N_BAR, проявляющие сродство к

BAR домены формируют гомодимер (BAR модуль) серповидной формы, состоящий из двух
BAR домены формируют гомодимер (BAR модуль) серповидной формы, состоящий из двух
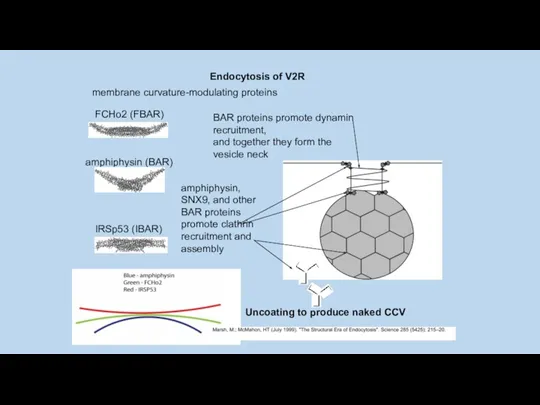
amphiphysin (BAR)
Uncoating to produce naked CCV
Endocytosis of V2R
amphiphysin (BAR)
Uncoating to produce naked CCV
Endocytosis of V2R
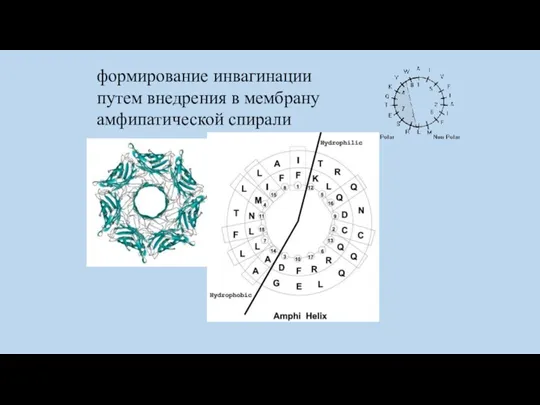
формирование инвагинации путем внедрения в мембрану амфипатической спирали
формирование инвагинации путем внедрения в мембрану амфипатической спирали

адаптерный комплекс AP2
AP2 – это узловой белок формирования динамичной
адаптерный комплекс AP2
AP2 – это узловой белок формирования динамичной
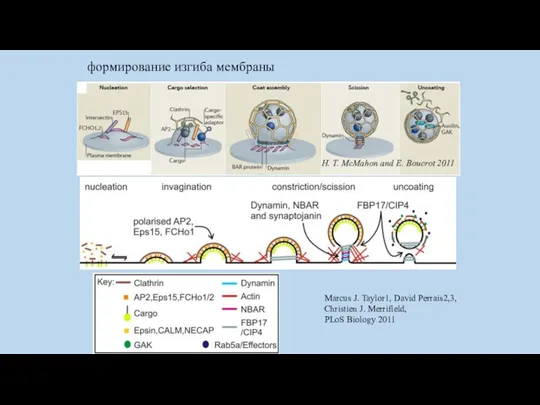
формирование изгиба мембраны
Marcus J. Taylor1, David Perrais2,3, Christien J. Merrifield,
PLoS
формирование изгиба мембраны
Marcus J. Taylor1, David Perrais2,3, Christien J. Merrifield,
PLoS
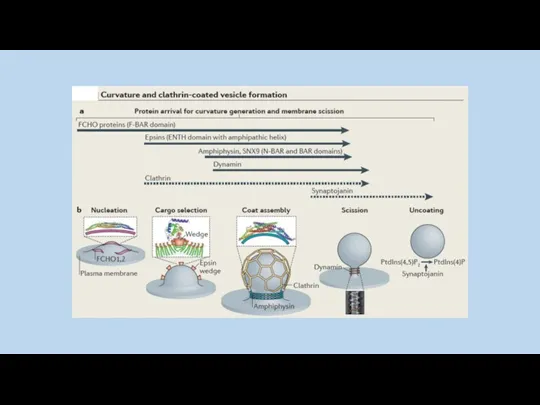

отделение клатриновой везикулы от плазматической мембраны.
Полимеризация клатрина способствует искривлению
отделение клатриновой везикулы от плазматической мембраны.
Полимеризация клатрина способствует искривлению

в регуляцию кривизны мембраны вовлечен актиновый цитоскелет.
На высокоразрешающих электронных изображениях
в регуляцию кривизны мембраны вовлечен актиновый цитоскелет.
На высокоразрешающих электронных изображениях

Везикулы отщепляются главным образом динамином.
В клетке динамин формирует спираль вокруг
Везикулы отщепляются главным образом динамином.
В клетке динамин формирует спираль вокруг

Динамин – белок размером порядка 100 кДа, обладающий GTPазной активностью и
Динамин – белок размером порядка 100 кДа, обладающий GTPазной активностью и
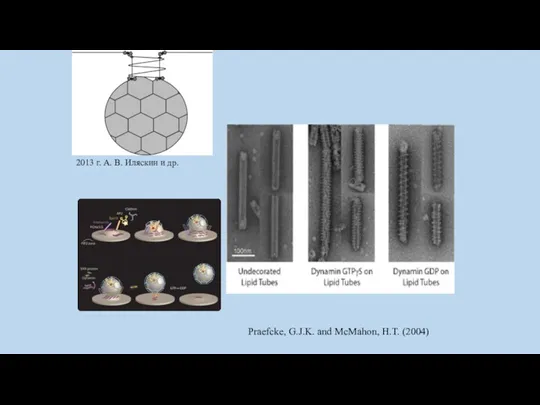
Praefcke, G.J.K. and McMahon, H.T. (2004)
2013 г. А. В. Иляскин
Praefcke, G.J.K. and McMahon, H.T. (2004)
2013 г. А. В. Иляскин
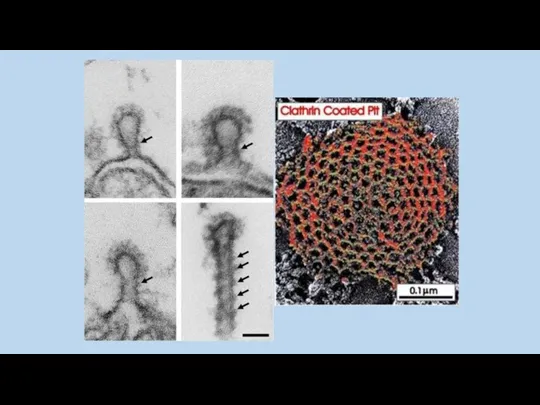

Последний этап клатринзависимого эндоцитоза – разрушение клатриновой оболочки ATP азой Hsc70,
Последний этап клатринзависимого эндоцитоза – разрушение клатриновой оболочки ATP азой Hsc70,
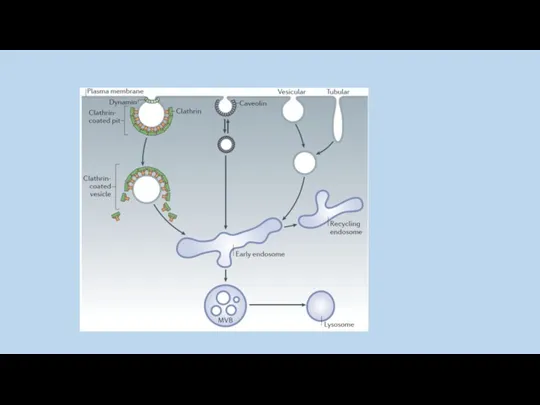

Интернализованные везикулы сливаются с ранними эндосомами , затем карго-белок рециклизуется в
Интернализованные везикулы сливаются с ранними эндосомами , затем карго-белок рециклизуется в

пять основных стадий формирования клатриновой везикулы:
инициация,
селекция интернализуемого белка,
сборка клатриновой
пять основных стадий формирования клатриновой везикулы:
инициация,
селекция интернализуемого белка,
сборка клатриновой
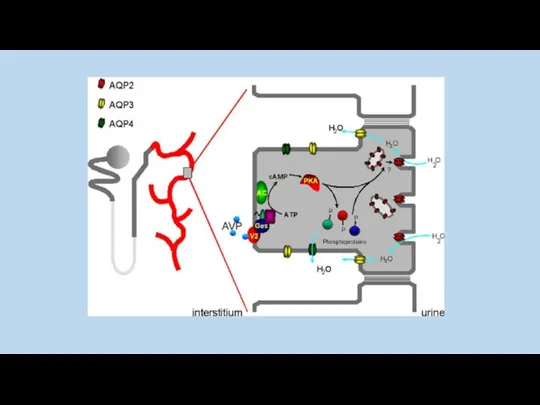
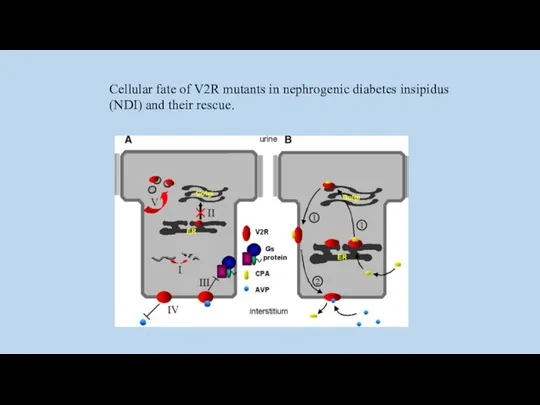
Cellular fate of V2R mutants in nephrogenic diabetes insipidus (NDI) and
Cellular fate of V2R mutants in nephrogenic diabetes insipidus (NDI) and

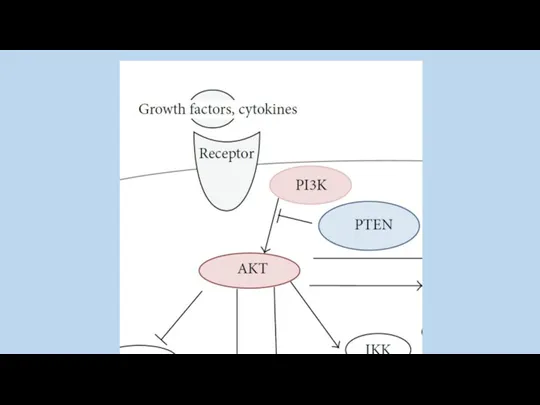

PTEN (название гена) (“phosphatase and tensin homolog”) семейство тирозиновых фосфатаз.
Участвует
PTEN (название гена) (“phosphatase and tensin homolog”) семейство тирозиновых фосфатаз.
Участвует
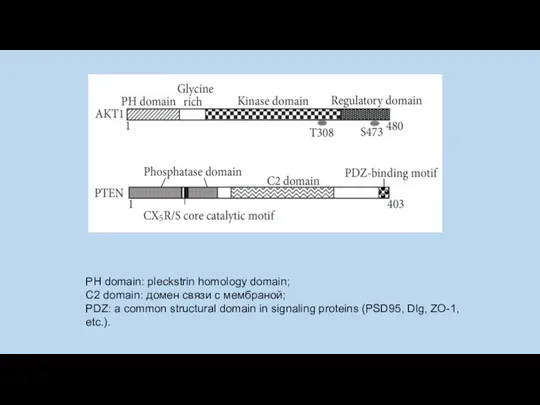
PH domain: pleckstrin homology domain;
C2 domain: домен связи с мембраной;
PH domain: pleckstrin homology domain;
C2 domain: домен связи с мембраной;

phosphatidylinositol-3 kinase (PI3K) участвует в регуляции клеточного роста и метаболизма. Активные
phosphatidylinositol-3 kinase (PI3K) участвует в регуляции клеточного роста и метаболизма. Активные

PIP3 (продукт PI3K) связывается с АКТ , что приводит к перемещению
PIP3 (продукт PI3K) связывается с АКТ , что приводит к перемещению

Суперсемейство EF-hand белков
CaM кальмодулин
MW 16,700 ; 148 aa; Kd
Суперсемейство EF-hand белков
CaM кальмодулин
MW 16,700 ; 148 aa; Kd


The soluble N-ethylmaleimide-sensitive factor attachment protein target receptor (SNARE) предполагается, участвуют
The soluble N-ethylmaleimide-sensitive factor attachment protein target receptor (SNARE) предполагается, участвуют

(synaptosome-associated protein) SNAP23
В почке его находят в апикальной мембране
(synaptosome-associated protein) SNAP23
В почке его находят в апикальной мембране
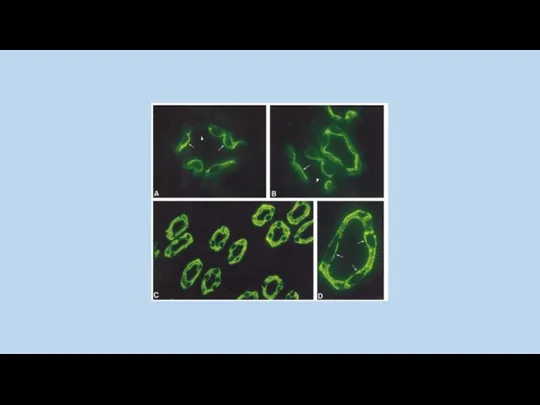

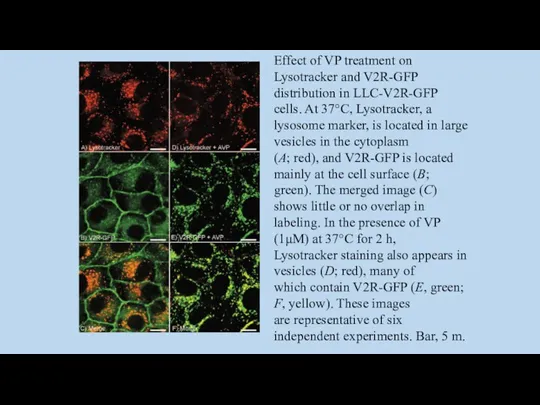
Effect of VP treatment on Lysotracker and V2R-GFP
distribution in LLC-V2R-GFP cells.
Effect of VP treatment on Lysotracker and V2R-GFP
distribution in LLC-V2R-GFP cells.
 Хранение дачного урожая
Хранение дачного урожая Penguins
Penguins Деление клеток
Деление клеток Прокариоты и эукариоты
Прокариоты и эукариоты Физиология синапсов
Физиология синапсов Животные с длинными ушами
Животные с длинными ушами В гости к весне
В гости к весне Антропология. Ранние этапы эволюции человека
Антропология. Ранние этапы эволюции человека Регуляция дыхания
Регуляция дыхания Половое размножение
Половое размножение Голонасінні рослини
Голонасінні рослини Дыхательная система человека
Дыхательная система человека Биотические связи в природе. 9 класс
Биотические связи в природе. 9 класс Генетическое картирование. Методы картирования генов
Генетическое картирование. Методы картирования генов Тип Хордовые
Тип Хордовые Ядовитые животные и растения
Ядовитые животные и растения Клетки прокариотических организмов
Клетки прокариотических организмов Sample Kit Kemira 3f. External
Sample Kit Kemira 3f. External Спинной мозг и ствол мозга
Спинной мозг и ствол мозга Микробиологиялық синтездегі өнімдерді шоғырландыру және анықтау тәсілдері
Микробиологиялық синтездегі өнімдерді шоғырландыру және анықтау тәсілдері презентация по биологии 7 класс Типы развития насекомых
презентация по биологии 7 класс Типы развития насекомых Бактериальная палеонтология. Бактериальная клетка и ее основные структуры
Бактериальная палеонтология. Бактериальная клетка и ее основные структуры Органаминералды тыңайтқыштар мен биотопырақ өндірісі
Органаминералды тыңайтқыштар мен биотопырақ өндірісі Листериоз. Род Listeria
Листериоз. Род Listeria Ткани человека
Ткани человека Роль белков в фомировании потребительских свойств продовольственных товаров
Роль белков в фомировании потребительских свойств продовольственных товаров Дыхание растений и животных
Дыхание растений и животных Семейство Куньи
Семейство Куньи