- Активный транспорт

Содержание
- 2. ОБЩИЕ СВОЙСТВА: ТРАНСПОРТ СОПРЯЖЕН С ГИДРОЛИЗОМ АТФ ПЕРЕНОС ИОНОВ ОСУЩЕСТВЛЯЮТ ТРАНСПОРТНЫЕ АТФАЗЫ ТРАНСПОРТ ОСУЩЕСТВЛЯЕТСЯ ПРОТИВ ГРАДИЕНТА
- 3. ТИПЫ АТФАЗ АТФаза V-типа АТФаза F-типа АТФаза Р-типа
- 4. АТФазы V типа ЛОКАЛИЗАЦИЯ: МЕМБРАНЫ ВАКОЛЕЙ ДРОЖЖЕЙ ТОНОПЛАСТЫ РАСТЕНИЙ ЛИЗОСОМЫ ЭНДОСОМЫ СЕКРЕТОРНЫЕ ГРАНУЛЫ
- 5. V0 – ГИДРОФОБНАЯ ЧАСТЬ (УЧАСТВУЕТ В ТРАНСЛОКАЦИИ ПРОТОНОВ) V1 – ВОДОРАСТВОРИМАЯ ЧАСТЬ ОБЛАДАЕТ КАТАЛИТИЧЕСКОЙ АКТИВНОСТЬЮ, КАТАЛИЗИРУЕТ
- 6. ФУНКЦИИ ПЕРЕНОСЯТ ПРОТОНЫ УЧАСТВУЮТ В ТРАНСПОРТЕ АНИОНОВ, АМИНОКИСЛОТ И РЕПАРАЦИИ МЕМБРАН ПРИ ЭНДО- И ЭКЗОЦИТОЗЕ ИНГИБИТОРЫ:
- 7. ТРАНСПОРТ ПРОТОНОВ В ТОНОПЛАСТЕ КЛЕТОК РАСТЕНИЙ ПРОТОНЫ ПОСТУПАЮТ ВНУТРЬ ВАКУОЛИ И СОЗДАЮТ ЭЛЕКТРИЧЕСКИЙ (от +20 до
- 8. АТФаза F ТИПА ЛОКАЛИЗАЦИЯ: МЕМБРАНЫ БАКТЕРИЙ, ХЛОРОПЛАСТОВ И МИТОХОНДРИЙ СТРОЕНИЕ: F0 – ГИДРОФОБНАЯ ЧАСТЬ (ТРАНСЛОКАЦИЯ ПРОТОНОВ)
- 9. АТФазы Р ТИПА ОБЩЕЕ СВОЙСТВО: ОБРАЗОВАНИЕ ФОСФОРИЛИРОВАННОГО ПРОДУКТА, КОТОРЫЙ УЧАСТВУЕТ В РЕАКЦИОННОЙ ЦИКЛЕ ПРИМЕРЫ: Na/K-АТФаза, Ca-АТФаза,
- 10. Na/K АТФаза ЛОКАЛИЗОВАНА НА ЦИТОПЛАЗМАТИЧЕСКОЙ МЕМБРАНЕ СОЗДАЕТ ГРАДИЕНТ ИОНОВ НАТРИЯ И КАЛИЯ
- 11. СТРУКТУРА Na+/K+ АТФазы: состоит из 2 полипептидных цепей α и β 1 – участок связывания ионов
- 12. УАБАИН – ингибитор Na,K-АТФазы Строфант
- 13. РЕАКЦИОННЫЙ ЦИКЛ Na/K АТФазы
- 14. 2 - изменение конформации транслоказы, вызванное присоединением 3Na+, приводит к активации каталитической субъединицы и увеличению сродства
- 15. 3 - аутофосфорилирование изменяет заряд и конформацию транслоказы, она закрывается с внутренней стороны мембраны и открывается
- 16. 5 - дефосфорилирование изменяет заряд и конформацию транслоказы, она закрывается с наружной стороны мембраны и открывается
- 17. РЕГУЛЯЦИЯ АКТИВНОСТИ Na/K АТФазы СООТНОШЕНИЕ Na/K И СОДЕРЖАНИЕ АТФ (факторы краткосрочной регуляции) ФОСФОРИЛИРОВАНИЕ ПРОТЕИНКИНАЗАМИ, ЧТО ПРИВОДИТ
- 18. Са2+ АТФаза ЛОКАЛИЗАЦИЯ: САРКО- (ЭНДО)-ПЛАЗМАТИЧЕСКИЙ РЕТИКУЛУМ ЦИТОПЛАЗМАТИЧЕСКАЯ МЕМБРАНА
- 19. ВСЕ Са2+ АТФазы – МОНОМЕРНЫЕ БЕЛКИ, Т.Е. СОСТОЯТ ИЗ ОДНОЙ ПОЛИПЕПТИДНОЙ ЦЕПИ Са2+ АТФаза СПР И
- 20. ФЛБ – фосфоламбан (у Са АТФазы саркоплазматического ретикулума),
- 21. В фосфорилированном состоянии фосфоламбан стимулирует кальциевый насос ретикулума миоцитов, ускоряя восстановление кальциевых градиентов.
- 22. КМ – кальмодулин (у Са АТФазы плазматической мембраны )
- 23. ЦИКЛ РАБОТЫ Са2+АТФазы СТАДИИ ГИДРОЛИЗА АТФ ЧЕРЕДУЮТСЯ СО СТАДИЯМИ ПЕРЕНОСА ИОНОВ КАЛЬЦИЯ
- 24. ЛОКАЛИЗАЦИЯ: ПЛАЗМАТИЧЕСКИЕ МЕМБРАНЫ РАСТИТЕЛЬНЫХ КЛЕТОК Н+АТФаза
- 25. Н+-АТФаза – это интегральный белок, полипептидная цепь которого десять раз пересекает поверхностную (плазматическую) мембрану. Полагают, что
- 27. Роль Н+-АТФазы: поддерживает рН цитоплазмы близкий к нейтральному создает на мембране разность потенциалов (∆ψ), во многом
- 28. Механизм возникновения ПД в проводящих пучках высших растений имеет большое сходство с таковым в нервах животных.
- 29. Благодаря вторичному активному транспорту клетка активно поглощает (или удаляет) многие вещества (ионы, углеводы, аминокислоты и др.).
- 30. ТРАНСПОРТ ВЕЩЕСТВ ЧЕРЕЗ МНОГОМЕМБРАННЫЕ СИСТЕМЫ ВТОРИЧНЫЙ АКТИВНЫЙ ТРАНСПОРТ
- 31. МНОГОМЕМБРАННЫЕ СИСТЕМЫ гликокаликс + коллаген микроворсинки базолатеральная поверхность
- 32. БАЗАЛЬНАЯ МЕМБРАНА – дополнительная мембрана за пределами плазмалеммы эпителиоцита ЕЕ СОСТАВ: гликопротеидный матрикс (гликокаликс) + коллагеновый
- 33. Базолатеральная мембрана несет межклеточные контакты разного типа
- 34. ЭКСПЕРИМЕНТЫ УССИНГА Установка Уссинга Объект исследований
- 35. ЭКСПЕРИМЕНТ УССИНГА: ИЗУЧЕНИЕ АСИММЕТРИЧНЫХ СВОЙСТВ ЭПИТЕЛИЯ 1 – кожа лягушки; 2 – вольтметр; 3 и 4
- 36. МОДЕЛЬ УССИНГА Мукозная (наружная)поверхность кожи Серозная (внутренняя)поверхность кожи
- 37. ВТОРИЧНЫЙ АКТИВНЫЙ ТРАНСПОРТ
- 38. ВТОРИЧНЫЙ АКТИВНЫЙ ТРАНСПОРТ ТРАНСПОРТ САХАРОВ И АМИНОКИСЛОТ ЗА СЧЕТ ЭНЕРГИИ ГРАДИЕНТА Na+ , КОТОРЫЙ СОЗДАЕТСЯ БЛАГОДАРЯ
- 39. Уравнение для транспорта сахаров Jmax = 12 мкмоль / м2 с – одинакова для всех моносахаридов
- 40. Графики Лайнуивера – Берка для транспорта 6-дезокси-D-глюкозы через эпителий кишки, показывающие зависимость транспорта сахара от концентрации
- 41. ТРАНСПОРТ ГЛЮКОЗЫ, СОПРЯЖЕННЫЙ С ИОНАМИ НАТРИЯ
- 42. ТРАНСПОРТ АМИНОКИСЛОТ, СОПРЯЖЕННЫЙ С ИОНАМИ НАТРИЯ
- 44. В настоящее время выделяют 5 транспортных систем: для крупных нейтральных, в том числе алифатических и ароматических
- 45. Глутатионовая система транспорта Второй способ переноса аминокислот внутрь клетки происходит в комплексе с глутатионом при помощи
- 47. Скачать презентацию

ОБЩИЕ СВОЙСТВА:
ТРАНСПОРТ СОПРЯЖЕН С ГИДРОЛИЗОМ АТФ
ПЕРЕНОС ИОНОВ ОСУЩЕСТВЛЯЮТ ТРАНСПОРТНЫЕ АТФАЗЫ
ТРАНСПОРТ
ОБЩИЕ СВОЙСТВА:
ТРАНСПОРТ СОПРЯЖЕН С ГИДРОЛИЗОМ АТФ
ПЕРЕНОС ИОНОВ ОСУЩЕСТВЛЯЮТ ТРАНСПОРТНЫЕ АТФАЗЫ
ТРАНСПОРТ
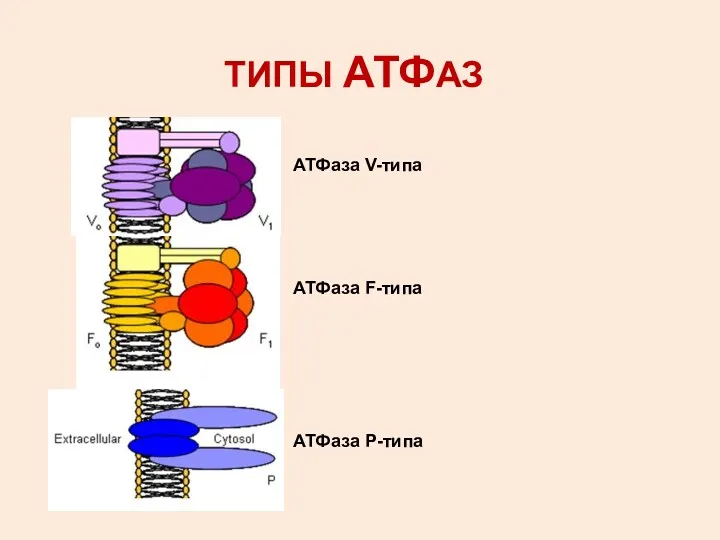
ТИПЫ АТФАЗ
АТФаза V-типа
АТФаза F-типа
АТФаза Р-типа
ТИПЫ АТФАЗ
АТФаза V-типа
АТФаза F-типа
АТФаза Р-типа
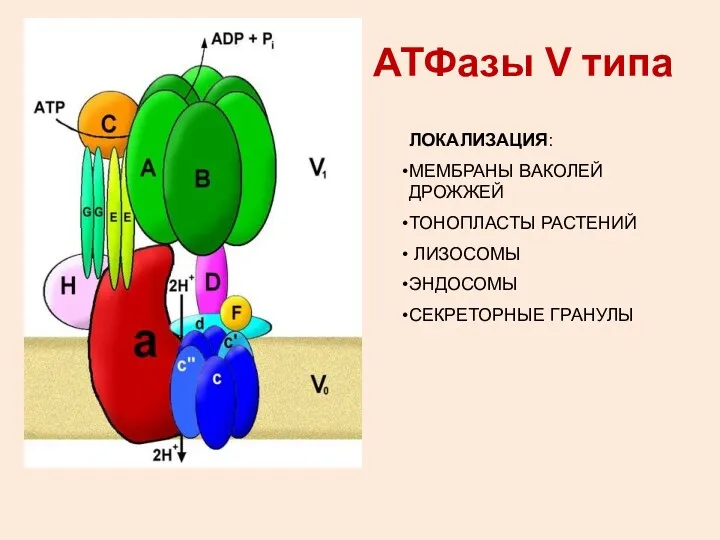
АТФазы V типа
ЛОКАЛИЗАЦИЯ:
МЕМБРАНЫ ВАКОЛЕЙ ДРОЖЖЕЙ
ТОНОПЛАСТЫ РАСТЕНИЙ
ЛИЗОСОМЫ
ЭНДОСОМЫ
СЕКРЕТОРНЫЕ ГРАНУЛЫ
АТФазы V типа
ЛОКАЛИЗАЦИЯ:
МЕМБРАНЫ ВАКОЛЕЙ ДРОЖЖЕЙ
ТОНОПЛАСТЫ РАСТЕНИЙ
ЛИЗОСОМЫ
ЭНДОСОМЫ
СЕКРЕТОРНЫЕ ГРАНУЛЫ
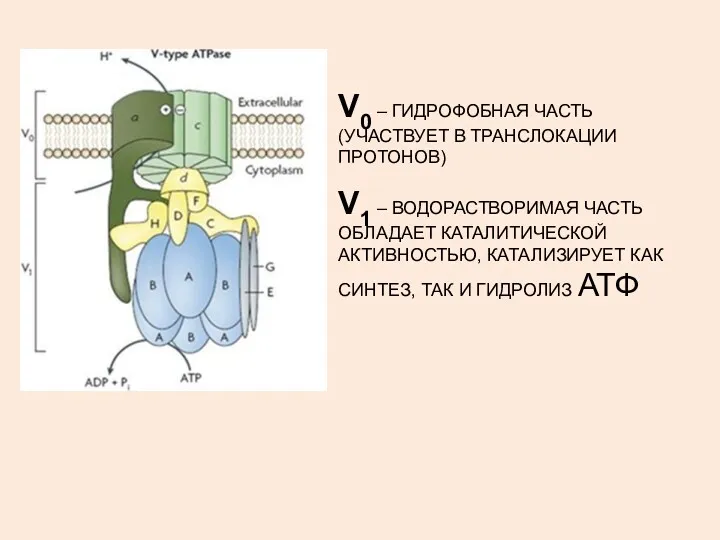
V0 – ГИДРОФОБНАЯ ЧАСТЬ (УЧАСТВУЕТ В ТРАНСЛОКАЦИИ ПРОТОНОВ)
V1 – ВОДОРАСТВОРИМАЯ ЧАСТЬ
V0 – ГИДРОФОБНАЯ ЧАСТЬ (УЧАСТВУЕТ В ТРАНСЛОКАЦИИ ПРОТОНОВ)
V1 – ВОДОРАСТВОРИМАЯ ЧАСТЬ
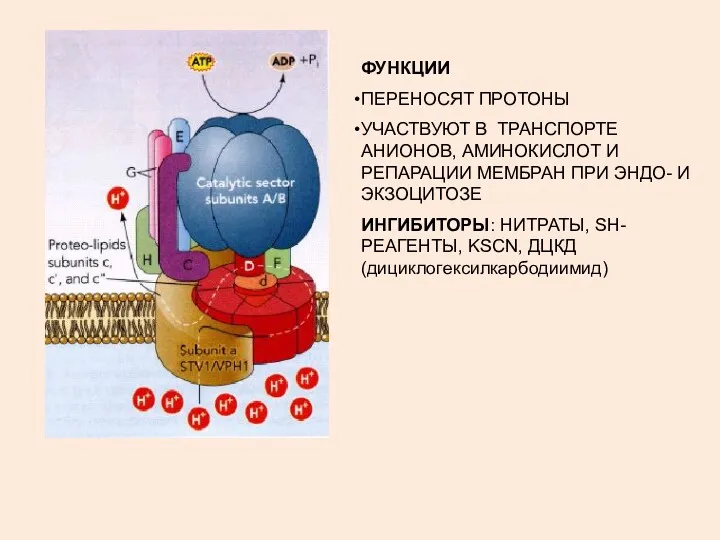
ФУНКЦИИ
ПЕРЕНОСЯТ ПРОТОНЫ
УЧАСТВУЮТ В ТРАНСПОРТЕ АНИОНОВ, АМИНОКИСЛОТ И РЕПАРАЦИИ МЕМБРАН ПРИ ЭНДО-
ФУНКЦИИ
ПЕРЕНОСЯТ ПРОТОНЫ
УЧАСТВУЮТ В ТРАНСПОРТЕ АНИОНОВ, АМИНОКИСЛОТ И РЕПАРАЦИИ МЕМБРАН ПРИ ЭНДО-
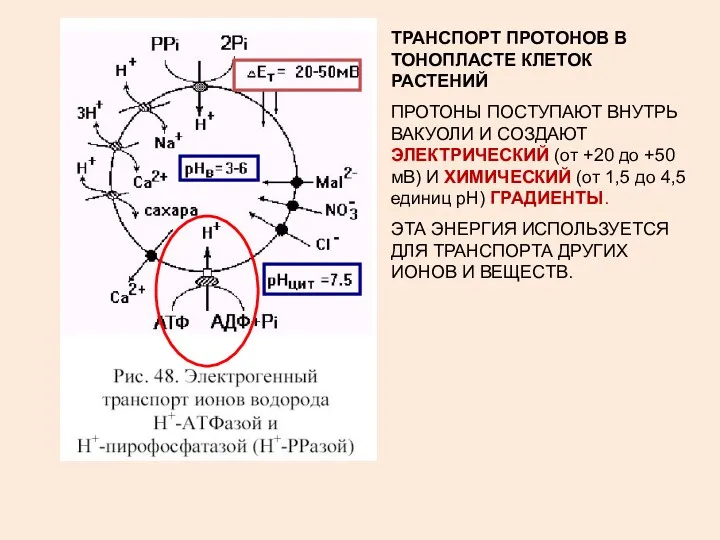
ТРАНСПОРТ ПРОТОНОВ В ТОНОПЛАСТЕ КЛЕТОК РАСТЕНИЙ
ПРОТОНЫ ПОСТУПАЮТ ВНУТРЬ ВАКУОЛИ И СОЗДАЮТ
ТРАНСПОРТ ПРОТОНОВ В ТОНОПЛАСТЕ КЛЕТОК РАСТЕНИЙ
ПРОТОНЫ ПОСТУПАЮТ ВНУТРЬ ВАКУОЛИ И СОЗДАЮТ
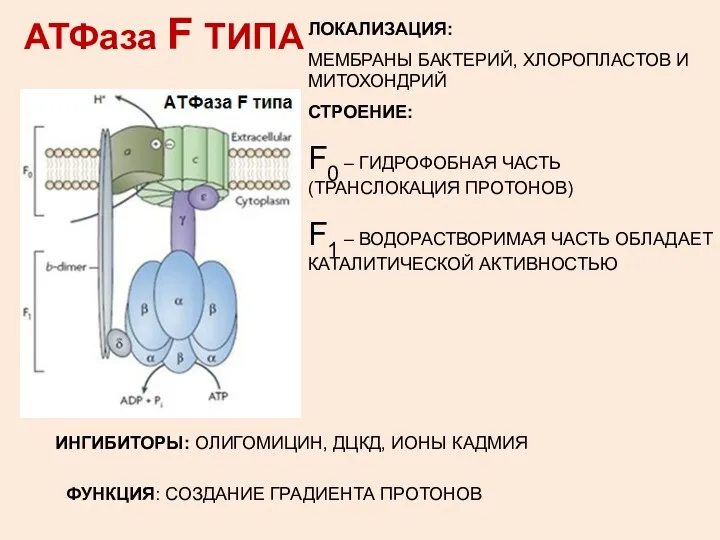
АТФаза F ТИПА
ЛОКАЛИЗАЦИЯ:
МЕМБРАНЫ БАКТЕРИЙ, ХЛОРОПЛАСТОВ И МИТОХОНДРИЙ
СТРОЕНИЕ:
F0 – ГИДРОФОБНАЯ ЧАСТЬ (ТРАНСЛОКАЦИЯ
АТФаза F ТИПА
ЛОКАЛИЗАЦИЯ:
МЕМБРАНЫ БАКТЕРИЙ, ХЛОРОПЛАСТОВ И МИТОХОНДРИЙ
СТРОЕНИЕ:
F0 – ГИДРОФОБНАЯ ЧАСТЬ (ТРАНСЛОКАЦИЯ

АТФазы Р ТИПА
ОБЩЕЕ СВОЙСТВО: ОБРАЗОВАНИЕ ФОСФОРИЛИРОВАННОГО ПРОДУКТА, КОТОРЫЙ УЧАСТВУЕТ В РЕАКЦИОННОЙ
АТФазы Р ТИПА
ОБЩЕЕ СВОЙСТВО: ОБРАЗОВАНИЕ ФОСФОРИЛИРОВАННОГО ПРОДУКТА, КОТОРЫЙ УЧАСТВУЕТ В РЕАКЦИОННОЙ
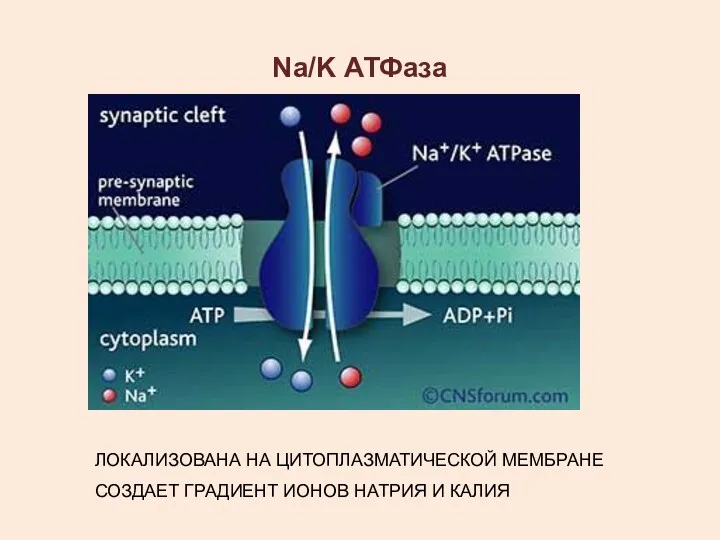
Na/K АТФаза
ЛОКАЛИЗОВАНА НА ЦИТОПЛАЗМАТИЧЕСКОЙ МЕМБРАНЕ
СОЗДАЕТ ГРАДИЕНТ ИОНОВ НАТРИЯ И КАЛИЯ
Na/K АТФаза
ЛОКАЛИЗОВАНА НА ЦИТОПЛАЗМАТИЧЕСКОЙ МЕМБРАНЕ
СОЗДАЕТ ГРАДИЕНТ ИОНОВ НАТРИЯ И КАЛИЯ
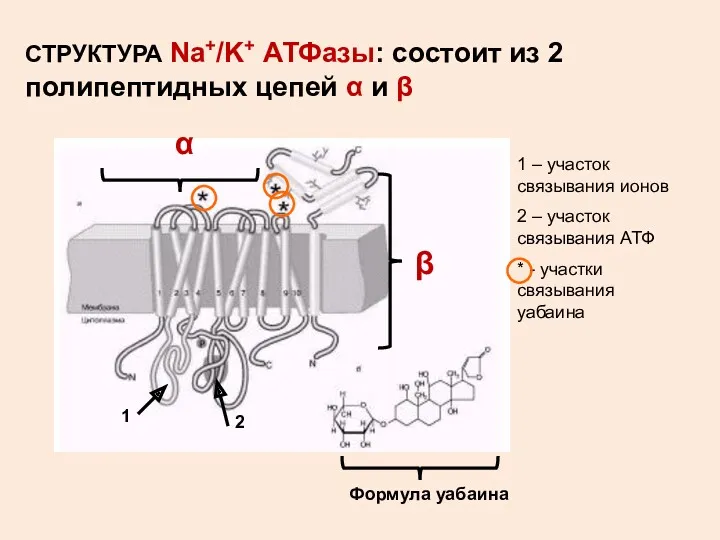
СТРУКТУРА Na+/K+ АТФазы: состоит из 2 полипептидных цепей α и β
1
СТРУКТУРА Na+/K+ АТФазы: состоит из 2 полипептидных цепей α и β
1
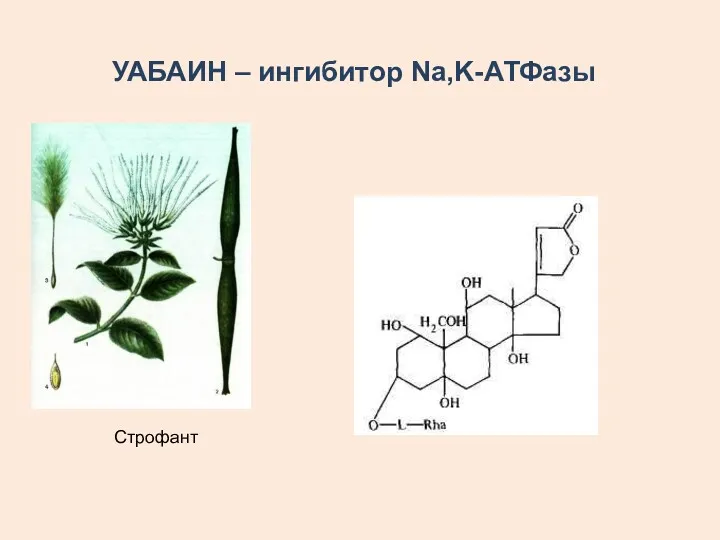
УАБАИН – ингибитор Na,K-АТФазы
Строфант
УАБАИН – ингибитор Na,K-АТФазы
Строфант
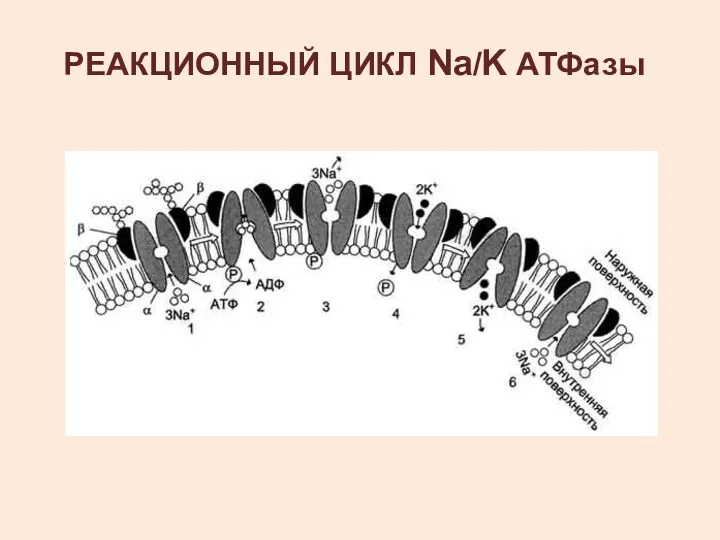
РЕАКЦИОННЫЙ ЦИКЛ Na/K АТФазы
РЕАКЦИОННЫЙ ЦИКЛ Na/K АТФазы
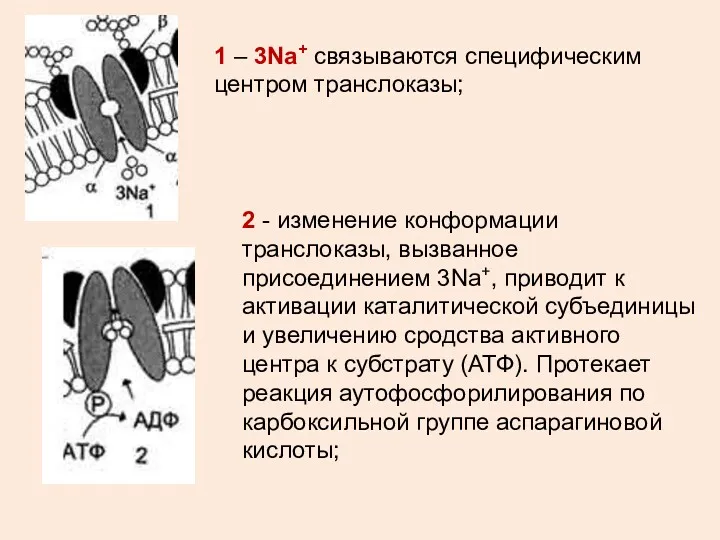
2 - изменение конформации транслоказы, вызванное присоединением 3Na+, приводит к активации
2 - изменение конформации транслоказы, вызванное присоединением 3Na+, приводит к активации

3 - аутофосфорилирование изменяет заряд и конформацию транслоказы, она закрывается с
3 - аутофосфорилирование изменяет заряд и конформацию транслоказы, она закрывается с
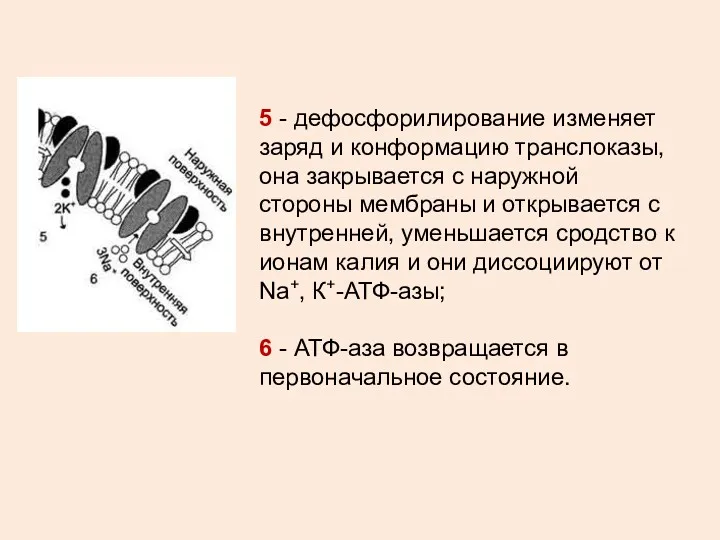
5 - дефосфорилирование изменяет заряд и конформацию транслоказы, она закрывается с
5 - дефосфорилирование изменяет заряд и конформацию транслоказы, она закрывается с

РЕГУЛЯЦИЯ АКТИВНОСТИ Na/K АТФазы
СООТНОШЕНИЕ Na/K И СОДЕРЖАНИЕ АТФ (факторы краткосрочной регуляции)
ФОСФОРИЛИРОВАНИЕ
РЕГУЛЯЦИЯ АКТИВНОСТИ Na/K АТФазы
СООТНОШЕНИЕ Na/K И СОДЕРЖАНИЕ АТФ (факторы краткосрочной регуляции)
ФОСФОРИЛИРОВАНИЕ
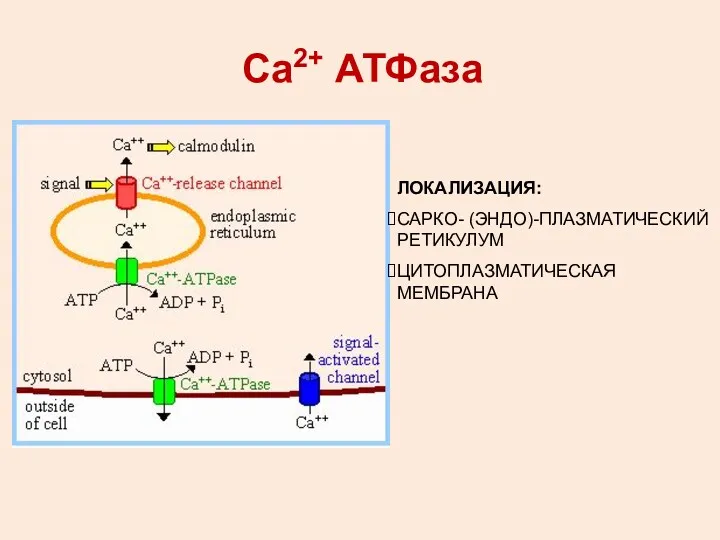
Са2+ АТФаза
ЛОКАЛИЗАЦИЯ:
САРКО- (ЭНДО)-ПЛАЗМАТИЧЕСКИЙ РЕТИКУЛУМ
ЦИТОПЛАЗМАТИЧЕСКАЯ МЕМБРАНА
Са2+ АТФаза
ЛОКАЛИЗАЦИЯ:
САРКО- (ЭНДО)-ПЛАЗМАТИЧЕСКИЙ РЕТИКУЛУМ
ЦИТОПЛАЗМАТИЧЕСКАЯ МЕМБРАНА

ВСЕ Са2+ АТФазы – МОНОМЕРНЫЕ БЕЛКИ, Т.Е. СОСТОЯТ ИЗ ОДНОЙ ПОЛИПЕПТИДНОЙ
ВСЕ Са2+ АТФазы – МОНОМЕРНЫЕ БЕЛКИ, Т.Е. СОСТОЯТ ИЗ ОДНОЙ ПОЛИПЕПТИДНОЙ
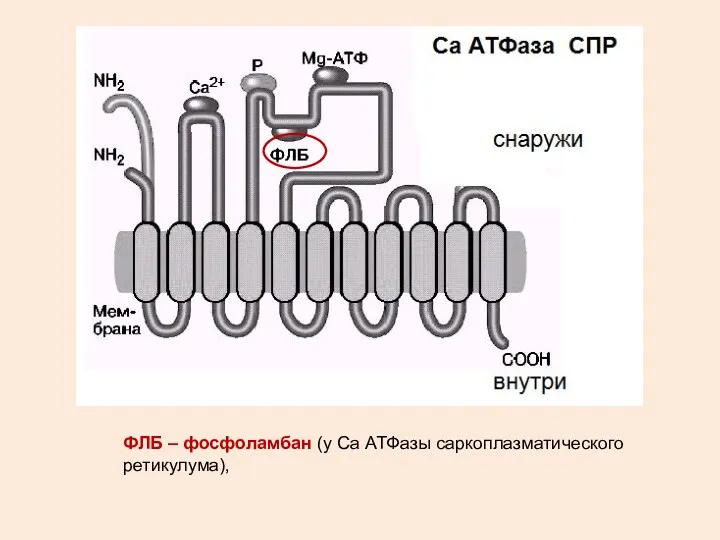
ФЛБ – фосфоламбан (у Са АТФазы саркоплазматического ретикулума),
ФЛБ – фосфоламбан (у Са АТФазы саркоплазматического ретикулума),
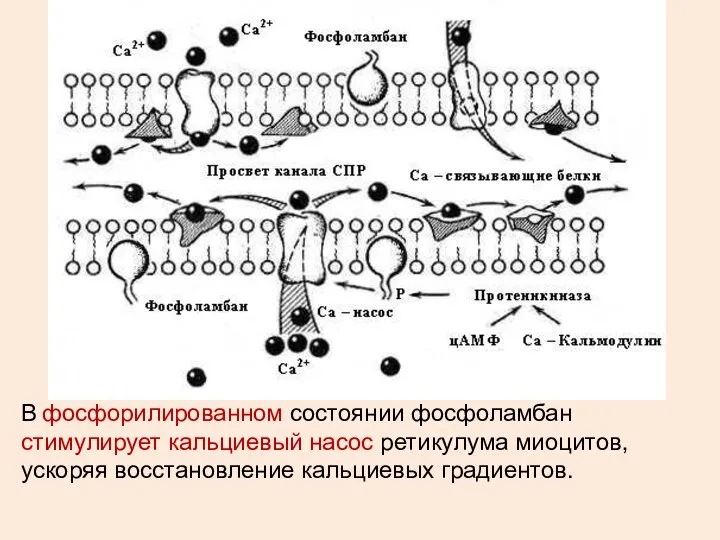
В фосфорилированном состоянии фосфоламбан стимулирует кальциевый насос ретикулума миоцитов, ускоряя восстановление
В фосфорилированном состоянии фосфоламбан стимулирует кальциевый насос ретикулума миоцитов, ускоряя восстановление
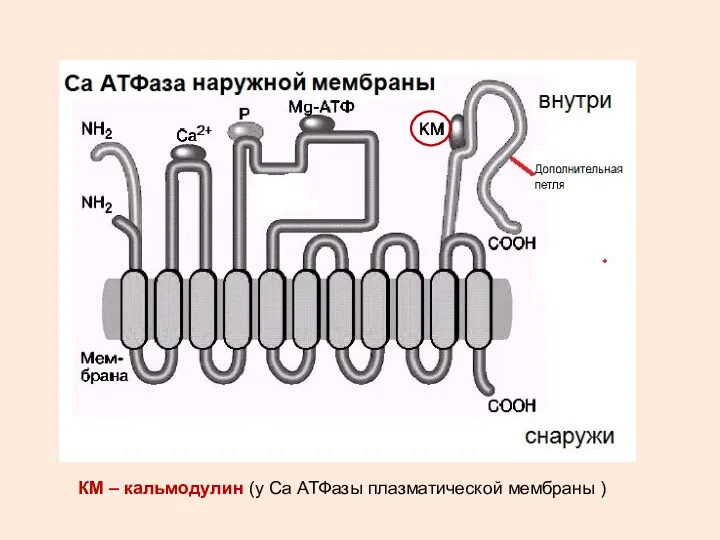
КМ – кальмодулин (у Са АТФазы плазматической мембраны )
КМ – кальмодулин (у Са АТФазы плазматической мембраны )
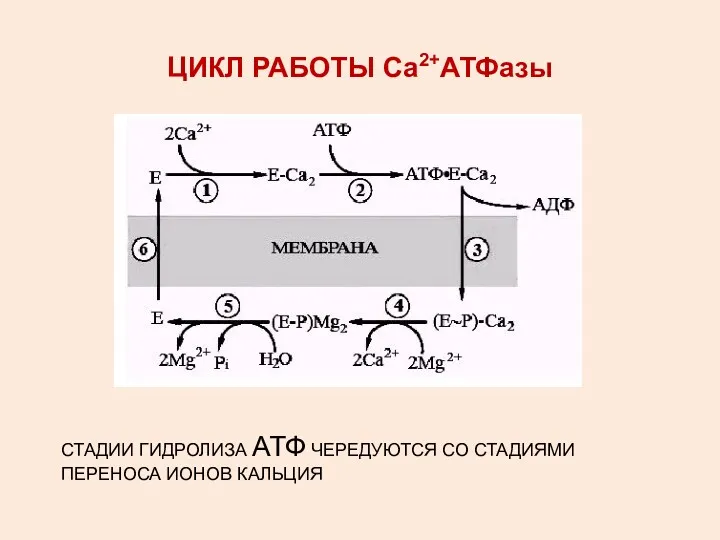
ЦИКЛ РАБОТЫ Са2+АТФазы
СТАДИИ ГИДРОЛИЗА АТФ ЧЕРЕДУЮТСЯ СО СТАДИЯМИ ПЕРЕНОСА ИОНОВ КАЛЬЦИЯ
ЦИКЛ РАБОТЫ Са2+АТФазы
СТАДИИ ГИДРОЛИЗА АТФ ЧЕРЕДУЮТСЯ СО СТАДИЯМИ ПЕРЕНОСА ИОНОВ КАЛЬЦИЯ
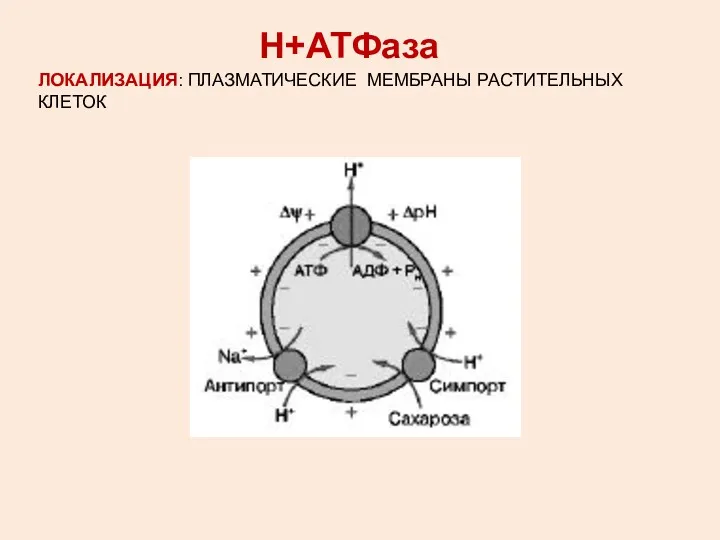
ЛОКАЛИЗАЦИЯ: ПЛАЗМАТИЧЕСКИЕ МЕМБРАНЫ РАСТИТЕЛЬНЫХ КЛЕТОК
Н+АТФаза
ЛОКАЛИЗАЦИЯ: ПЛАЗМАТИЧЕСКИЕ МЕМБРАНЫ РАСТИТЕЛЬНЫХ КЛЕТОК
Н+АТФаза

Н+-АТФаза – это интегральный белок, полипептидная цепь которого десять раз пересекает
Н+-АТФаза – это интегральный белок, полипептидная цепь которого десять раз пересекает


Роль Н+-АТФазы:
поддерживает рН цитоплазмы близкий к нейтральному
создает на мембране разность
Роль Н+-АТФазы:
поддерживает рН цитоплазмы близкий к нейтральному
создает на мембране разность
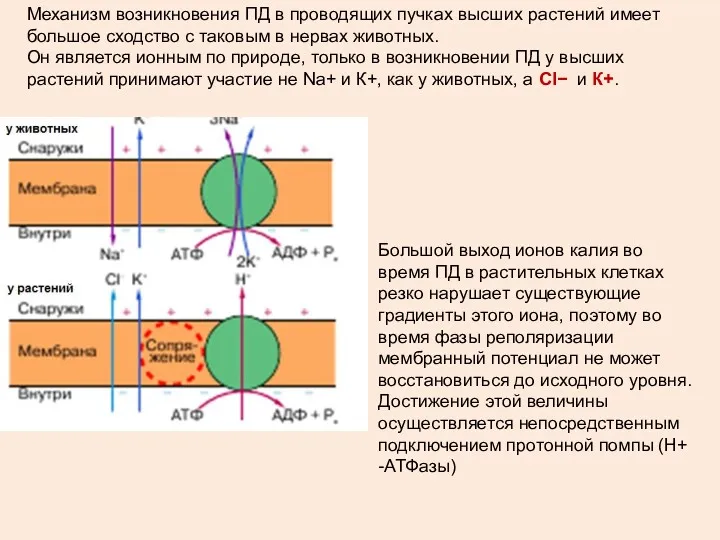
Механизм возникновения ПД в проводящих пучках высших растений имеет большое сходство
Механизм возникновения ПД в проводящих пучках высших растений имеет большое сходство
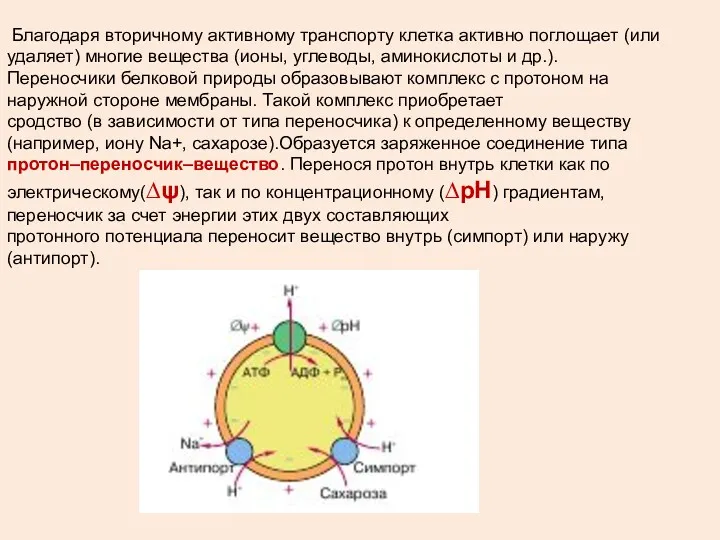
Благодаря вторичному активному транспорту клетка активно поглощает (или удаляет) многие
Благодаря вторичному активному транспорту клетка активно поглощает (или удаляет) многие

ТРАНСПОРТ ВЕЩЕСТВ ЧЕРЕЗ МНОГОМЕМБРАННЫЕ СИСТЕМЫ
ВТОРИЧНЫЙ АКТИВНЫЙ ТРАНСПОРТ
ТРАНСПОРТ ВЕЩЕСТВ ЧЕРЕЗ МНОГОМЕМБРАННЫЕ СИСТЕМЫ
ВТОРИЧНЫЙ АКТИВНЫЙ ТРАНСПОРТ
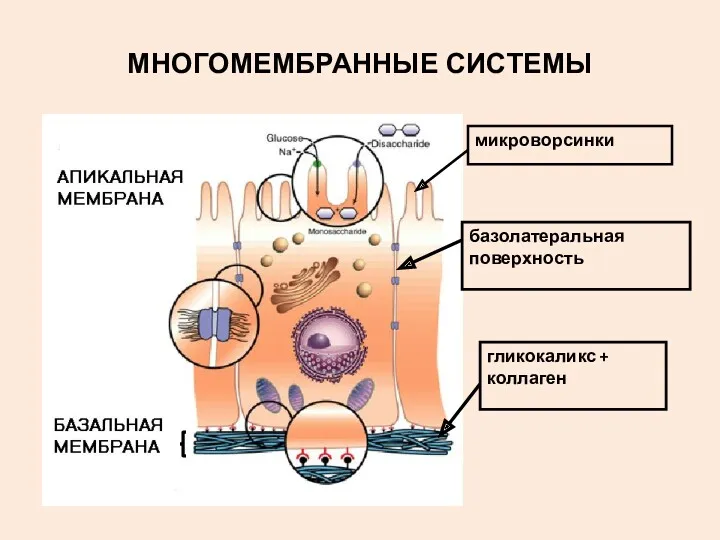
МНОГОМЕМБРАННЫЕ СИСТЕМЫ
гликокаликс + коллаген
микроворсинки
базолатеральная поверхность
МНОГОМЕМБРАННЫЕ СИСТЕМЫ
гликокаликс + коллаген
микроворсинки
базолатеральная поверхность

БАЗАЛЬНАЯ МЕМБРАНА – дополнительная мембрана за пределами плазмалеммы эпителиоцита
ЕЕ СОСТАВ: гликопротеидный
БАЗАЛЬНАЯ МЕМБРАНА – дополнительная мембрана за пределами плазмалеммы эпителиоцита
ЕЕ СОСТАВ: гликопротеидный
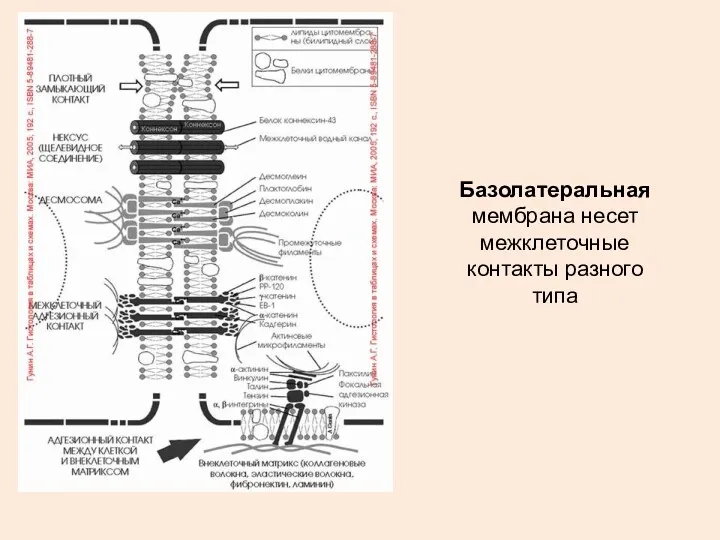
Базолатеральная мембрана несет межклеточные контакты разного типа
Базолатеральная мембрана несет межклеточные контакты разного типа
ЭКСПЕРИМЕНТЫ УССИНГА
Установка Уссинга
Объект исследований
ЭКСПЕРИМЕНТЫ УССИНГА
Установка Уссинга
Объект исследований
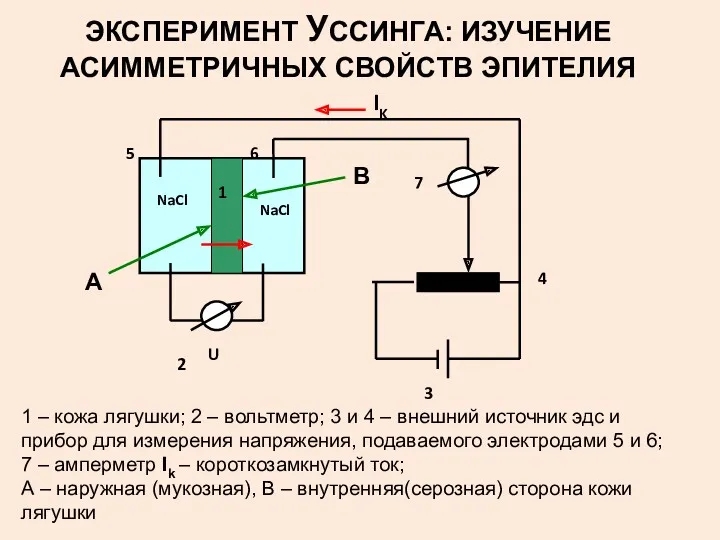
ЭКСПЕРИМЕНТ УССИНГА: ИЗУЧЕНИЕ АСИММЕТРИЧНЫХ СВОЙСТВ ЭПИТЕЛИЯ
1 – кожа лягушки; 2 –
ЭКСПЕРИМЕНТ УССИНГА: ИЗУЧЕНИЕ АСИММЕТРИЧНЫХ СВОЙСТВ ЭПИТЕЛИЯ
1 – кожа лягушки; 2 –
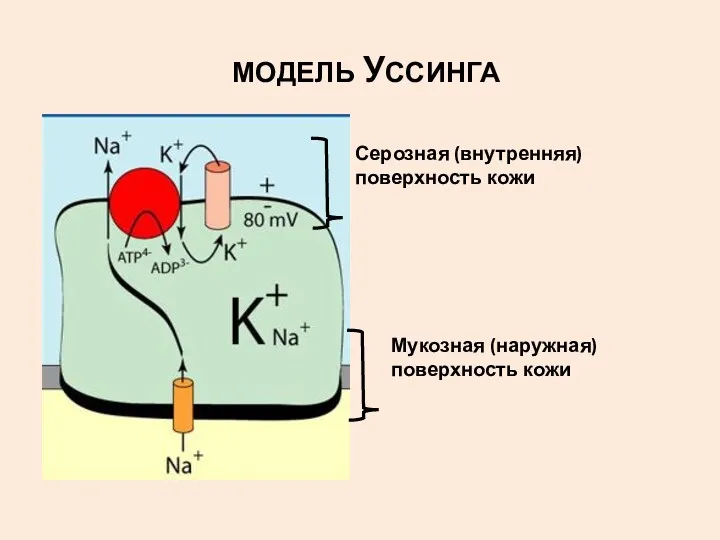
МОДЕЛЬ УССИНГА
Мукозная (наружная)поверхность кожи
Серозная (внутренняя)поверхность кожи
МОДЕЛЬ УССИНГА
Мукозная (наружная)поверхность кожи
Серозная (внутренняя)поверхность кожи
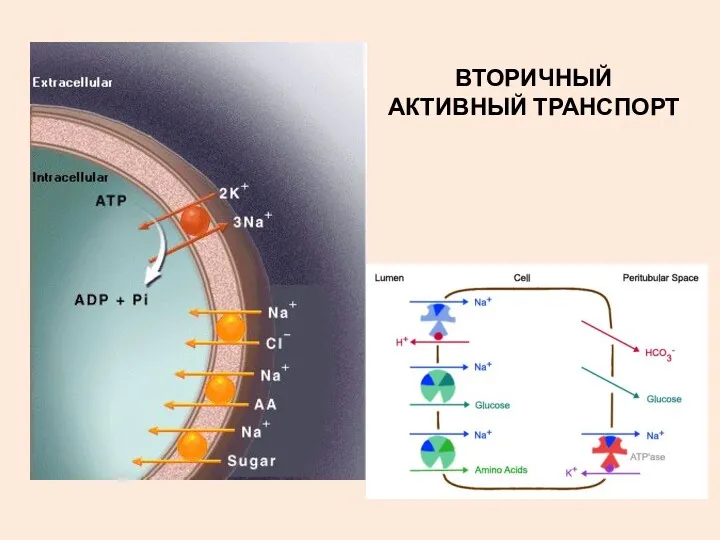
ВТОРИЧНЫЙ АКТИВНЫЙ ТРАНСПОРТ
ВТОРИЧНЫЙ АКТИВНЫЙ ТРАНСПОРТ

ВТОРИЧНЫЙ АКТИВНЫЙ ТРАНСПОРТ
ТРАНСПОРТ САХАРОВ И АМИНОКИСЛОТ ЗА СЧЕТ ЭНЕРГИИ ГРАДИЕНТА Na+
ВТОРИЧНЫЙ АКТИВНЫЙ ТРАНСПОРТ
ТРАНСПОРТ САХАРОВ И АМИНОКИСЛОТ ЗА СЧЕТ ЭНЕРГИИ ГРАДИЕНТА Na+

Уравнение для транспорта сахаров
Jmax = 12 мкмоль / м2 с –
Уравнение для транспорта сахаров
Jmax = 12 мкмоль / м2 с –
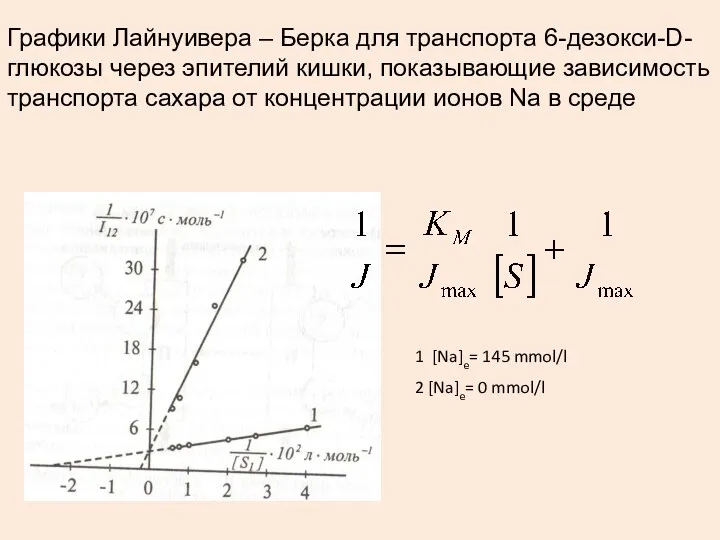
Графики Лайнуивера – Берка для транспорта 6-дезокси-D-глюкозы через эпителий кишки, показывающие
Графики Лайнуивера – Берка для транспорта 6-дезокси-D-глюкозы через эпителий кишки, показывающие
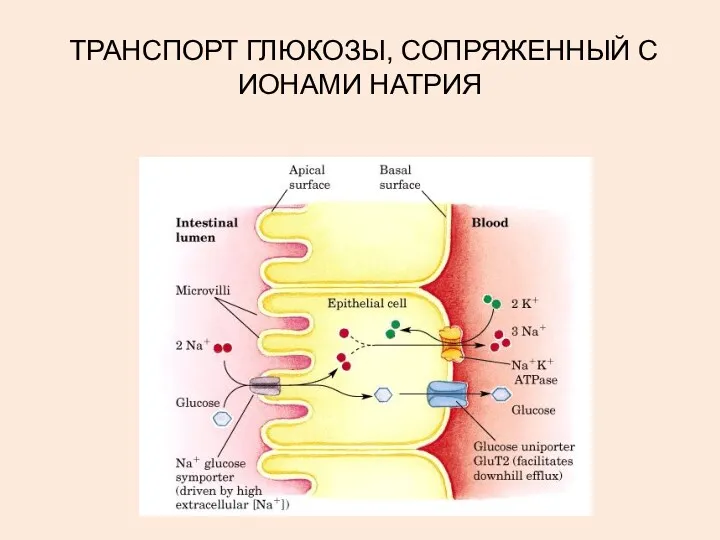
ТРАНСПОРТ ГЛЮКОЗЫ, СОПРЯЖЕННЫЙ С ИОНАМИ НАТРИЯ
ТРАНСПОРТ ГЛЮКОЗЫ, СОПРЯЖЕННЫЙ С ИОНАМИ НАТРИЯ
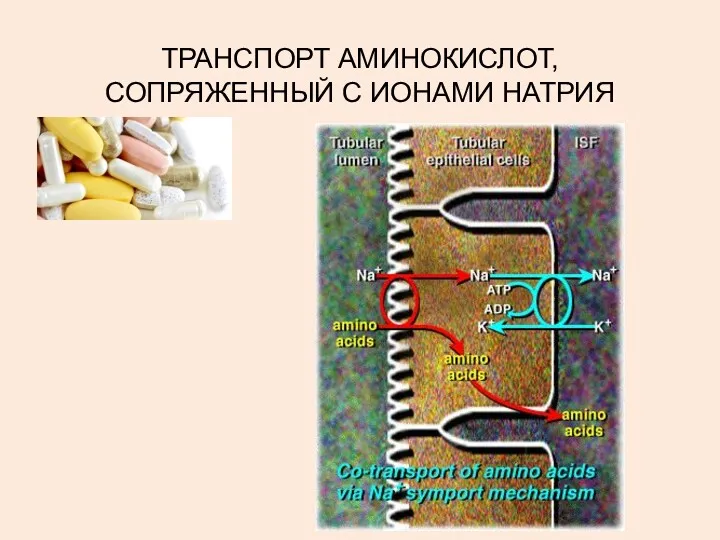
ТРАНСПОРТ АМИНОКИСЛОТ, СОПРЯЖЕННЫЙ С ИОНАМИ НАТРИЯ
ТРАНСПОРТ АМИНОКИСЛОТ, СОПРЯЖЕННЫЙ С ИОНАМИ НАТРИЯ
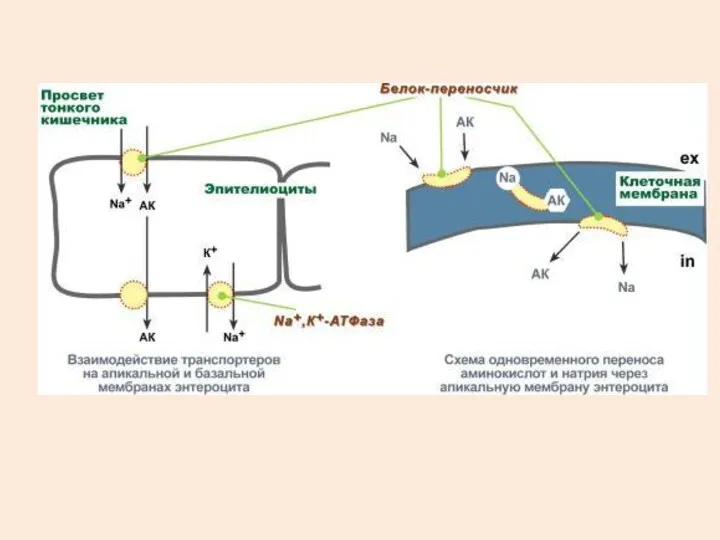

В настоящее время выделяют 5 транспортных систем:
для крупных нейтральных, в том
В настоящее время выделяют 5 транспортных систем:
для крупных нейтральных, в том
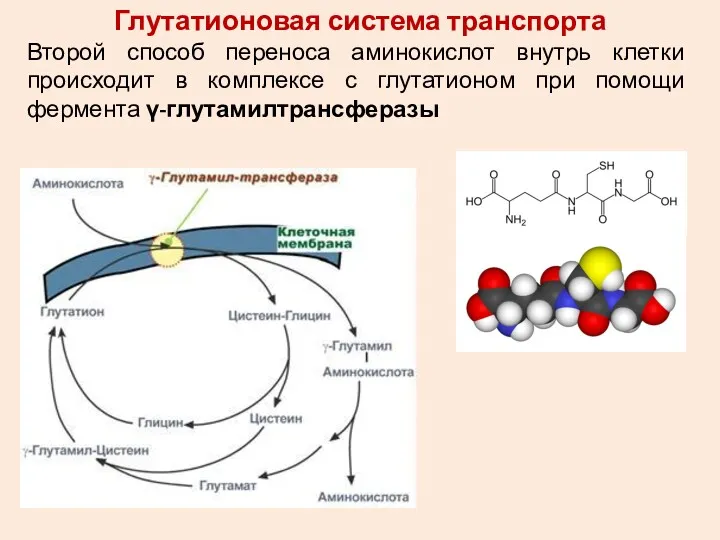
Глутатионовая система транспорта
Второй способ переноса аминокислот внутрь клетки происходит в комплексе
Глутатионовая система транспорта
Второй способ переноса аминокислот внутрь клетки происходит в комплексе
 Комнатные растения в интерьере
Комнатные растения в интерьере Cellular neurophysiology
Cellular neurophysiology Зеленые водоросли
Зеленые водоросли Спинной мозг человека
Спинной мозг человека Апоптоз. Исследования феномена программируемой клеточной смерти
Апоптоз. Исследования феномена программируемой клеточной смерти Ознакомление с фитонцидными растениями и выявление возможности их использования в интерьере. Практическая работа
Ознакомление с фитонцидными растениями и выявление возможности их использования в интерьере. Практическая работа Ритмы здоровья. Витаминно-минеральный комплекс
Ритмы здоровья. Витаминно-минеральный комплекс Чарлз Дарвин об эволюции органического мира
Чарлз Дарвин об эволюции органического мира урок биологии 6 класс по теме Побег
урок биологии 6 класс по теме Побег Эволюция человека
Эволюция человека Корова – домашнее и сельскохозяйственное животное
Корова – домашнее и сельскохозяйственное животное Онтогенез. Размножение организмов
Онтогенез. Размножение организмов Выращивание томата в теплице и парнике. 6 класс
Выращивание томата в теплице и парнике. 6 класс Подготовка к ЕГЭ. Решение генетических задач по схемам родословных
Подготовка к ЕГЭ. Решение генетических задач по схемам родословных Плоды и семена
Плоды и семена Приобретенные формы поведения. (Лекция 3)
Приобретенные формы поведения. (Лекция 3) Презентация по теме Основные типы экологических взаимодействий
Презентация по теме Основные типы экологических взаимодействий Новосибирский зоопарк
Новосибирский зоопарк Отдел Злаковые
Отдел Злаковые Клеточная оболочка. Оболочка животных и растительных клеток
Клеточная оболочка. Оболочка животных и растительных клеток Тваринний світ України
Тваринний світ України Органические вещества клетки – нуклеиновые кислоты ДНК и РНК
Органические вещества клетки – нуклеиновые кислоты ДНК и РНК Начальные этапы развития жизни на Земле
Начальные этапы развития жизни на Земле Synecology or - Community Ecology лекция
Synecology or - Community Ecology лекция Опасные дикие животные Якутии (Волк). 6 класс
Опасные дикие животные Якутии (Волк). 6 класс Методы исследования генетики человека
Методы исследования генетики человека Нервная система, ее структурно-функциональная характеристика. Центральная нервная система
Нервная система, ее структурно-функциональная характеристика. Центральная нервная система Клеточная теория. Цитология
Клеточная теория. Цитология