- Аминокислоты. Белки. Лекция 2

Содержание
- 2. 1. Классификация и физико-химические свойства протеиногенных аминокислот
- 3. Протеиногенные аминокислоты – аминокислоты, из которых синтезируются белки, т.е. это те аминокислоты, которые кодируются геномом (20
- 4. Структурная формула аминокислоты H │ H2N — Cα — COOH │ R
- 5. Классификации аминокислот: 1. Структурная (по строению бокового радикала) – ациклические (алифатические), циклические (ароматические, гетероциклические), иминокислота –
- 6. Классификация аминокислот по их химическому строению
- 7. Классификация аминокислот по их химическому строению
- 8. Классификация аминокислот по их химическому строению
- 9. Свойства аминокислот Все протеиногенные аминокислоты, за исключением глицина, имеют как минимум один асимметрический атом углерода и
- 10. Свойства стереоизомеров аминокислот
- 11. Изменение суммарного заряда аминокислот в зависимости от рН среды (амфотерность)
- 12. В зависимости от рН среды аминокислоты имеют суммарный нулевой, положительный или отрицательный заряд. Состояние, в котором
- 13. Образование пептидов в результате реакций конденсации Дипептид Пептидная связь Пептидная группа
- 14. Схема образования пептидной связи
- 15. Плоская пептидная группа (на красном фоне). C―N-связь частично имеет характер двойной связи и поэтому вокруг неё
- 16. 2. Классификация белков: простые и сложные, глобулярные и фибриллярные, мономерные и олигомерные
- 17. Классификация пептидов и белков по количеству аминокислотных остатков Олигопептиды – пептиды, содержащие до 10 аминокислот (трипептид,
- 18. Принцип строения пептидов
- 19. Ограничения, налагаемые на вращение вокруг одинарных связей полипептидной цепи. C―N-связи, входящие в состав плоских пептидных групп
- 20. Плоскости расположения пептидных групп и α-углеродных атомов в пространстве
- 21. Классификация пептидов по физиологическому действию Пептиды, обладающие гормональной активностью (окситоцин, вазопрессин, рилизинг-гормоны, меланоцитстимулирующий гормон, глюкагон и
- 22. Виды классификаций белков: 1. По составу (простые и сложные). 2. По форме (глобулярные и фибриллярные). 3.
- 23. Классификации белков По молекулярной массе - низкомолекулярные, высокомолекулярные.
- 24. Классификации белков По химическому строению (отсутствие или наличие небелковой части – простые или сложные): простые (при
- 25. Классификации белков По локализации в клетке (ядерные, цитоплазматические, мембраносвязанные, лизосомальные и др.). По локализации в организме
- 26. Количество протомеров в структуре олигомерных белков В состав олигомерных белков может входить от 2 до нескольких
- 27. Субъединичная структура глутаминсинтетазы
- 28. Классификации белков По форме молекул (глобулярные, фибриллярные). Глобулярные (соотношение продольной и поперечной осей не превышает 1:10,
- 29. Схема структурной организации коллагена а – первичная структура коллагена; б – первичная структура коллагена в виде
- 30. 3. Физико-химические свойства белков: растворимость, амфотерность, ионизация, гидратация, осаждение
- 31. Физико-химические свойства белков: растворимость; амфотерность; ионизация; гидратация белков; осаждение; буферные свойства; коллоидные и осмотические свойства; оптические
- 32. 4. Уровни структурной организации белков: первичная, вторичная, надвторичная, третичная и четвертичная структуры, домены, надмолекулярные структуры. Связи,
- 33. Структура белков Белки представляют собой цепи, содержащие десятки, сотни и тысячи аминокислотных остатков, соединённых прочными ковалентными
- 34. Возможные конформации белков
- 35. Первичная структура белков Первичная структура белков – последовательность аминокислот в полипептидной цепи, качественный и количественный состав
- 36. Вторичная структура белков Вторичная структура белков – пространственная структура, образующаяся в результате взаимодействий между функциональными группами,
- 37. α-Спираль Пространственное строение α-спирали-зованного участка полипептидной цепи и образование водородных связей, участвующих в формировании α-спирали
- 38. Вторичная структура белка в виде β-складчатого слоя
- 39. Параллельный и антипараллельный β-складчатые слои β-структуры обозначены широкими стрелками. А – антипараллельная β-структура; Б – параллельные
- 40. Супервторичная (надвторичная) структура белков Специфический порядок формирования вторичной структуры, содержащий сочетания элементов вторичной структуры (α-спирали и
- 41. Классификация супервторичных структур («структурные мотивы») «α-спираль-поворот-α-спираль» (ДНК-связывающий белок); «структура β-бочонка» (домен пируваткиназы, триозофосфатизомераза); «лейциновая застёжка-молния» (молекулы
- 42. Фрагмент ДНК-связывающего белка в форме «цинкового пальца»
- 43. Если полипептидная цепь белка содержит более 200 аминокислот, то, как правило, её пространственная структура сформирована в
- 44. Конформации белков Доменное строение глобулярных белков: а – β-субъединица гемоглобина; б – константный домен иммуноглобулина; в
- 45. Конформации белков Третичная структура белков – трехмерная пространственная структура, образующаяся за счет взаимодействий между радикалами аминокислот,
- 46. Связи, принимающие участие в формировании третичной структуры белков
- 47. Типы связей, возникающих между радикалами аминокислот при формировании третичной структуры 1. Ионные связи 2.Водородные связи 3.
- 48. Связи, принимающие участие в формировании третичной структуры белков Гидрофобные радикалы аминокислот стремятся к объединению внутри глобулярной
- 49. Связи, принимающие участие в формировании третичной структуры белков Ионные связи могут возникать между отрицательно заряженными (анионными)
- 50. Структура волоса и входящего в его состав α-кератина. Три α-спиральные цепи образуют скрученный (суперспирализованный) трёхжильный «канат».
- 51. Аминокислота цистеин, имеющая в R-группе атом серы, способна с такой же молекулой образовывать прочную, ковалентную дисульфидную
- 52. Последовательные стадии перманентной завивки волос. А – прямой волос. Б – Разрушение поперечных связей. В –
- 53. Дисульфидные связи в структуре гормона инсулина
- 54. Все белки с одинаковой первичной структурой, находящиеся в одинаковых условиях, приобретают одинаковую, характерную для данного индивидуального
- 55. Конформации белков Количество и взаиморасположение полипептидных цепей в пространстве называют «четвертичная структура белков». Отдельные полипептидные цепи
- 56. Строение гемоглобина
- 57. Модель гемоглобина человека Центральная полость Гем
- 58. 5. Взаимосвязь структуры и функции. Денатурация и ренатурация
- 59. Денатурация – изменение нативной структуры белка под действием различных факторов (химические – концентрированные кислоты, щёлочи, спирты
- 60. Структура нативной молекулы белка (в центре) и трех денатурированных молекул этого же белка
- 61. Денатурация и ренатурация рибонуклеазы А – нативная молекула рибонуклеазы, в третичной структуре которой имеются 4 дисульфидных
- 62. Формирование трёхмерной структуры белка в клетке
- 63. Форма белка определяется с помощью шаперонов
- 64. Болезни, связанные с нарушением фолдинга белков Некоторые растворимые в воде белки при изменении условий могут приобретать
- 65. 6. Функции белков: структурная, каталитическая, транспортная, рецепторная, регуляторная, защитная, сократительная и др.
- 66. Классификации белков По выполняемым функциям: ферменты (катализаторы – каталаза, амилаза и др.), регуляторные (гормоны и др.),
- 67. 7. Свойства простых белков. Гистоны, альбумины. Структурные белки: тубулины, кератины, коллаген, эластин
- 68. Простые белки при гидролизе распадаются только на свободные аминокислоты. Простые белки делятся на основании некоторых условно
- 69. Гистоны – являются белками основного характера, растворимы в солевых растворах. Гистоны выполняют две основные функции: они
- 70. Формирование нуклеосомы
- 71. Альбумины (от лат. albumen, род. падеж albuminis – белок) – растворимы в воде и солевых растворах,
- 72. Кератины – семейство фибриллярных белков, обладающих механической прочностью, которая среди материалов биологического происхождения уступает лишь хитину.
- 73. Тубулин – основной белок микротрубочек, обладающий сократительными функциями; димер, состоящий из α- и β-цепей с молекулярными
- 74. Коллаген – составляет 25-33% от общего количества белка организма взрослого человека, или 6% от массы тела.
- 75. Эластин – основной белковый компонент, из которого состоят эластические волокна. При ферментативном гидролизе эластина в гидролизате
- 77. Скачать презентацию

1. Классификация и физико-химические свойства протеиногенных аминокислот
1. Классификация и физико-химические свойства протеиногенных аминокислот

Протеиногенные аминокислоты – аминокислоты, из которых синтезируются белки, т.е. это те
Протеиногенные аминокислоты – аминокислоты, из которых синтезируются белки, т.е. это те
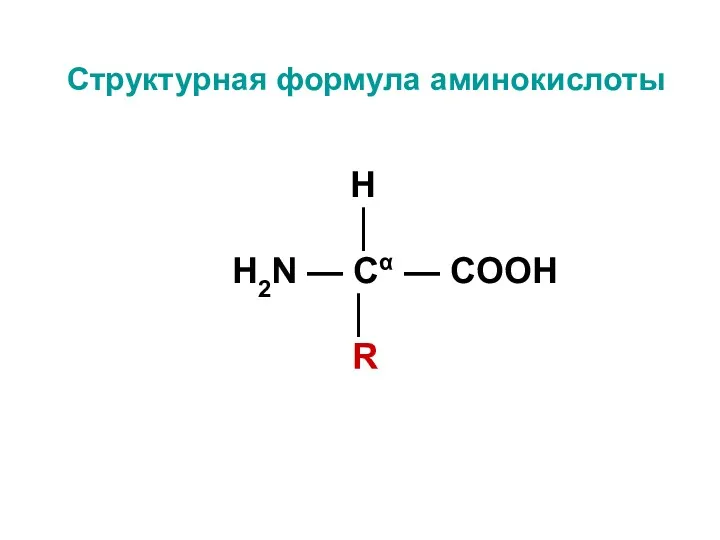
Структурная формула аминокислоты
H
│
H2N — Cα — COOH
Структурная формула аминокислоты
H
│
H2N — Cα — COOH

Классификации аминокислот:
1. Структурная (по строению бокового радикала) – ациклические (алифатические), циклические
Классификации аминокислот:
1. Структурная (по строению бокового радикала) – ациклические (алифатические), циклические

Классификация аминокислот по их химическому строению
Классификация аминокислот по их химическому строению

Классификация аминокислот по их химическому строению
Классификация аминокислот по их химическому строению
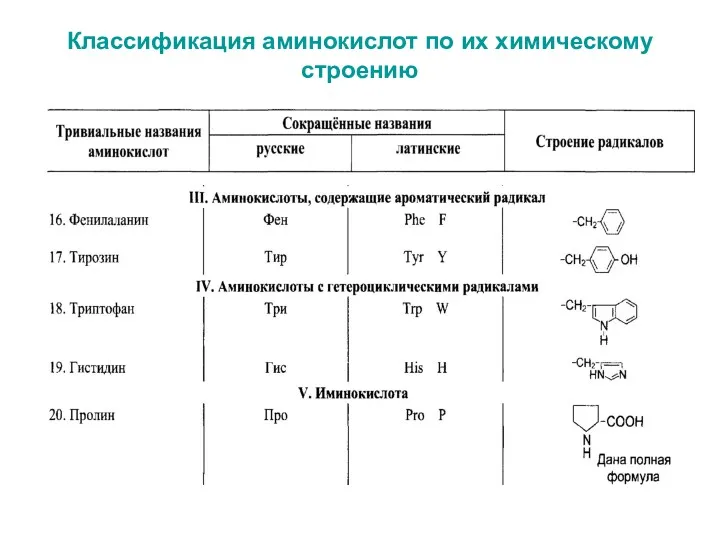
Классификация аминокислот по их химическому строению
Классификация аминокислот по их химическому строению
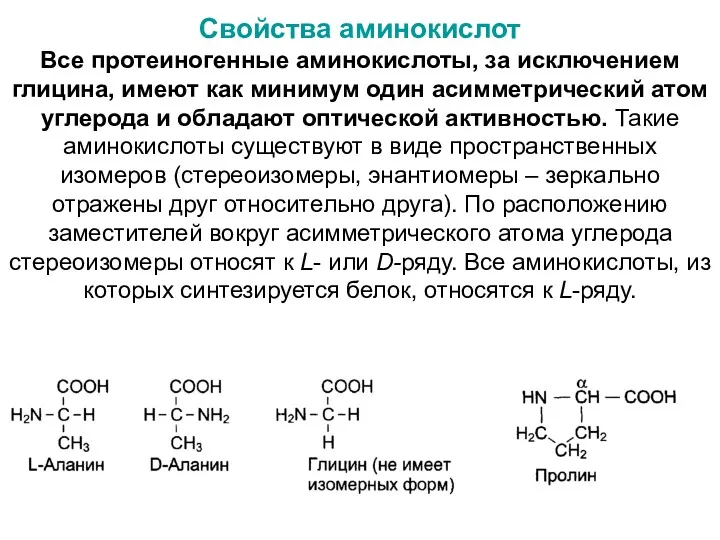
Свойства аминокислот
Все протеиногенные аминокислоты, за исключением глицина, имеют как минимум один
Свойства аминокислот Все протеиногенные аминокислоты, за исключением глицина, имеют как минимум один

Свойства стереоизомеров аминокислот
Свойства стереоизомеров аминокислот
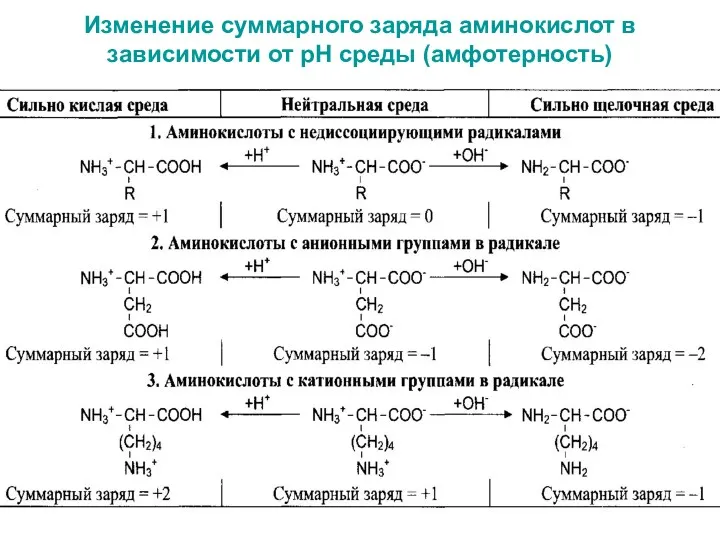
Изменение суммарного заряда аминокислот в зависимости от рН среды (амфотерность)
Изменение суммарного заряда аминокислот в зависимости от рН среды (амфотерность)

В зависимости от рН среды аминокислоты имеют суммарный нулевой, положительный или
В зависимости от рН среды аминокислоты имеют суммарный нулевой, положительный или
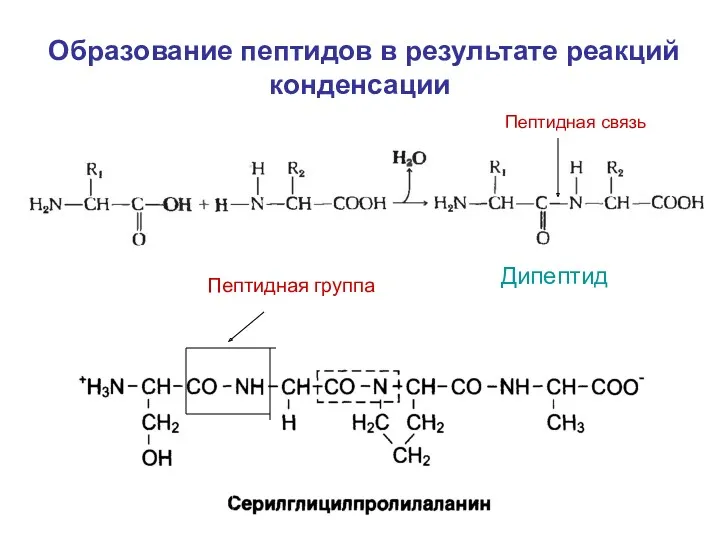
Образование пептидов в результате реакций конденсации
Дипептид
Пептидная связь
Пептидная группа
Образование пептидов в результате реакций конденсации
Дипептид
Пептидная связь
Пептидная группа
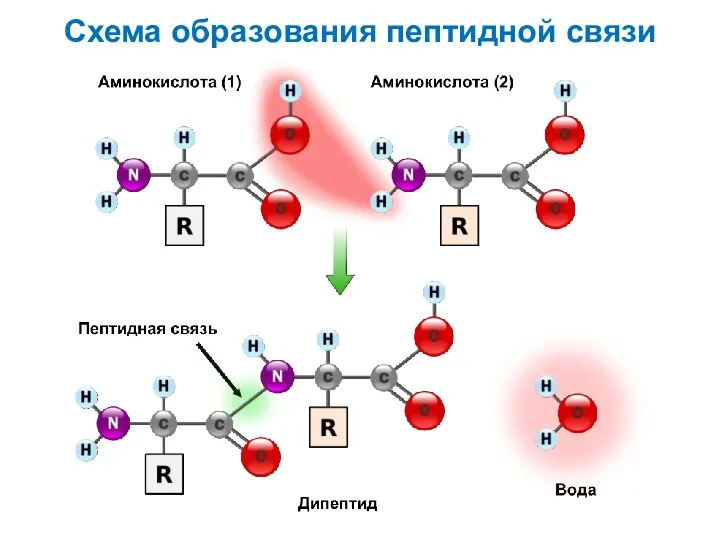
Схема образования пептидной связи
Схема образования пептидной связи
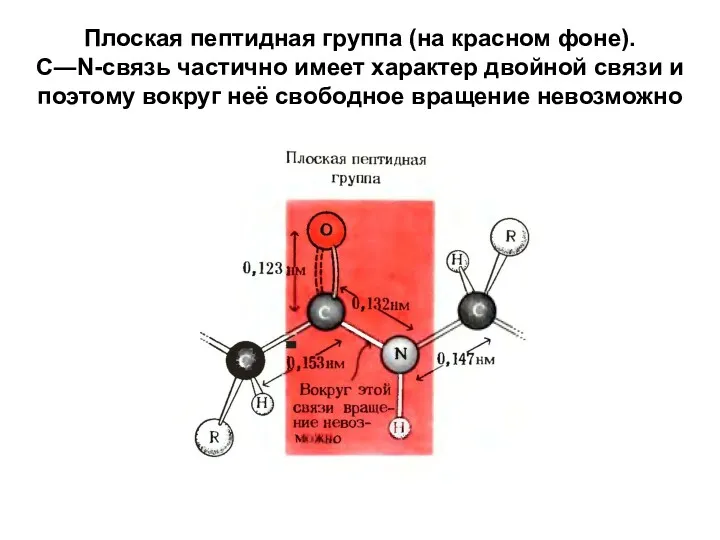
Плоская пептидная группа (на красном фоне).
C―N-связь частично имеет характер двойной связи
Плоская пептидная группа (на красном фоне). C―N-связь частично имеет характер двойной связи

2. Классификация белков: простые и сложные, глобулярные и фибриллярные, мономерные и
2. Классификация белков: простые и сложные, глобулярные и фибриллярные, мономерные и

Классификация пептидов и белков по количеству аминокислотных остатков
Олигопептиды – пептиды, содержащие
Классификация пептидов и белков по количеству аминокислотных остатков
Олигопептиды – пептиды, содержащие
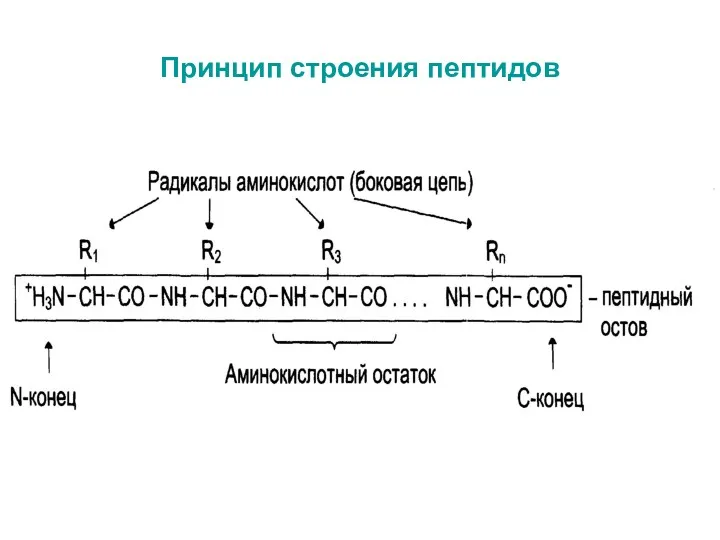
Принцип строения пептидов
Принцип строения пептидов
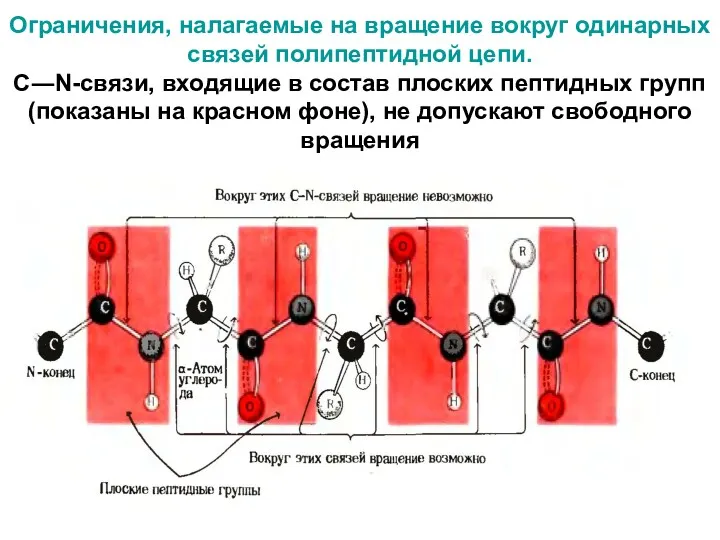
Ограничения, налагаемые на вращение вокруг одинарных связей полипептидной цепи.
C―N-связи, входящие в
Ограничения, налагаемые на вращение вокруг одинарных связей полипептидной цепи. C―N-связи, входящие в
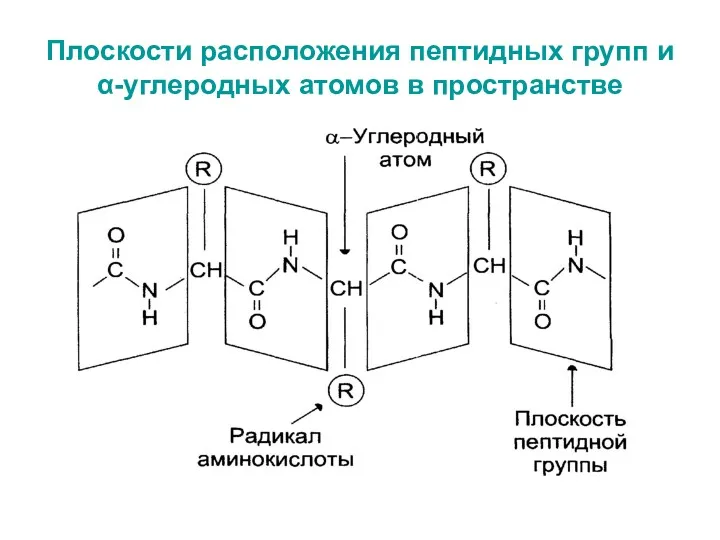
Плоскости расположения пептидных групп и α-углеродных атомов в пространстве
Плоскости расположения пептидных групп и α-углеродных атомов в пространстве

Классификация пептидов по физиологическому действию
Пептиды, обладающие гормональной активностью (окситоцин, вазопрессин, рилизинг-гормоны,
Классификация пептидов по физиологическому действию
Пептиды, обладающие гормональной активностью (окситоцин, вазопрессин, рилизинг-гормоны,

Виды классификаций белков:
1. По составу (простые и сложные).
2. По форме (глобулярные
Виды классификаций белков:
1. По составу (простые и сложные).
2. По форме (глобулярные
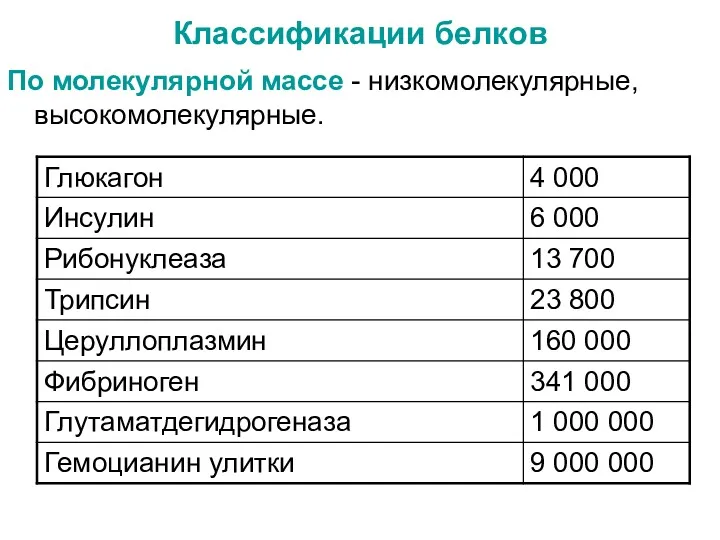
Классификации белков
По молекулярной массе - низкомолекулярные, высокомолекулярные.
Классификации белков
По молекулярной массе - низкомолекулярные, высокомолекулярные.

Классификации белков
По химическому строению (отсутствие или наличие небелковой части – простые
Классификации белков
По химическому строению (отсутствие или наличие небелковой части – простые

Классификации белков
По локализации в клетке (ядерные, цитоплазматические, мембраносвязанные, лизосомальные и др.).
По
Классификации белков
По локализации в клетке (ядерные, цитоплазматические, мембраносвязанные, лизосомальные и др.).
По

Количество протомеров в структуре олигомерных белков
В состав олигомерных белков может входить
Количество протомеров в структуре олигомерных белков
В состав олигомерных белков может входить
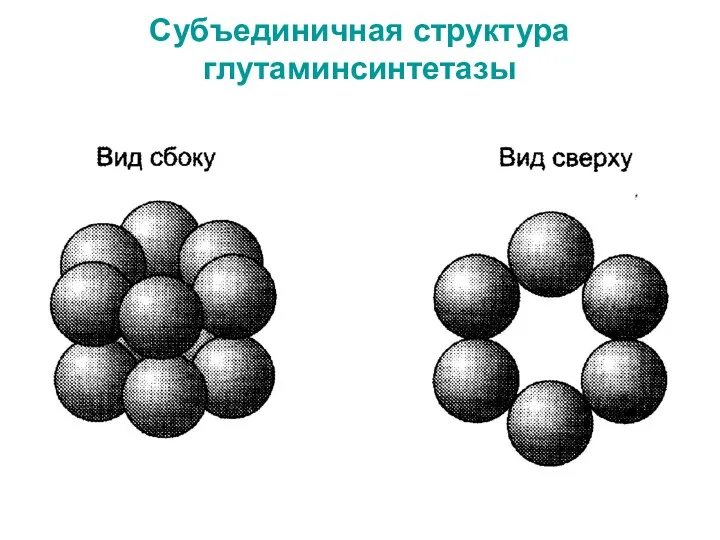
Субъединичная структура глутаминсинтетазы
Субъединичная структура глутаминсинтетазы

Классификации белков
По форме молекул (глобулярные, фибриллярные). Глобулярные (соотношение продольной и поперечной
Классификации белков
По форме молекул (глобулярные, фибриллярные). Глобулярные (соотношение продольной и поперечной
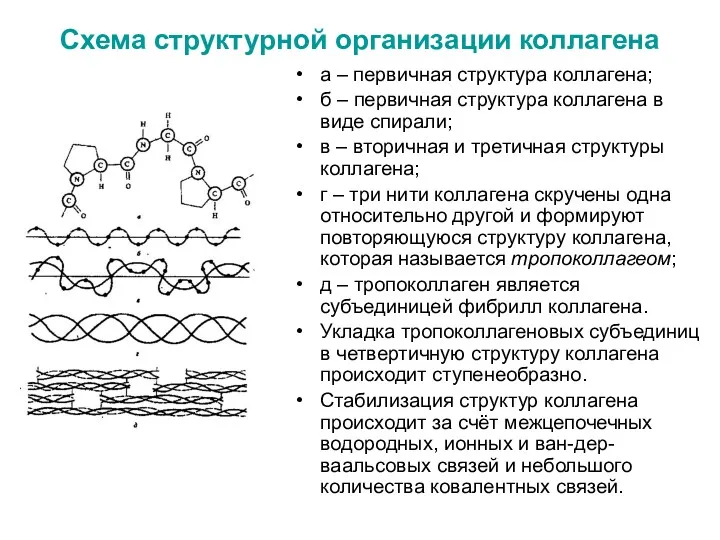
Схема структурной организации коллагена
а – первичная структура коллагена;
б – первичная
Схема структурной организации коллагена
а – первичная структура коллагена;
б – первичная

3. Физико-химические свойства белков: растворимость, амфотерность, ионизация, гидратация, осаждение
3. Физико-химические свойства белков: растворимость, амфотерность, ионизация, гидратация, осаждение

Физико-химические свойства белков:
растворимость;
амфотерность;
ионизация;
гидратация белков;
осаждение;
буферные свойства;
коллоидные и осмотические свойства;
оптические
Физико-химические свойства белков:
растворимость;
амфотерность;
ионизация;
гидратация белков;
осаждение;
буферные свойства;
коллоидные и осмотические свойства;
оптические

4. Уровни структурной организации белков: первичная, вторичная, надвторичная, третичная и четвертичная
4. Уровни структурной организации белков: первичная, вторичная, надвторичная, третичная и четвертичная

Структура белков
Белки представляют собой цепи, содержащие десятки, сотни и тысячи аминокислотных
Структура белков
Белки представляют собой цепи, содержащие десятки, сотни и тысячи аминокислотных
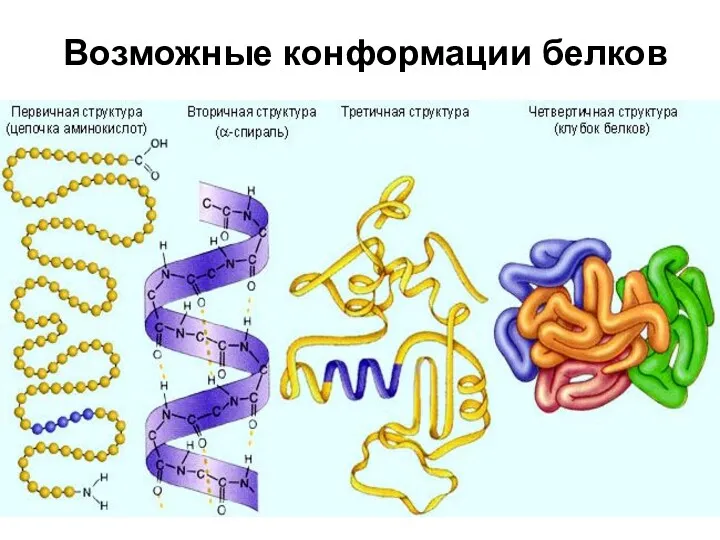
Возможные конформации белков
Возможные конформации белков

Первичная структура белков
Первичная структура белков – последовательность аминокислот в полипептидной цепи,
Первичная структура белков
Первичная структура белков – последовательность аминокислот в полипептидной цепи,

Вторичная структура белков
Вторичная структура белков – пространственная структура, образующаяся в результате
Вторичная структура белков
Вторичная структура белков – пространственная структура, образующаяся в результате
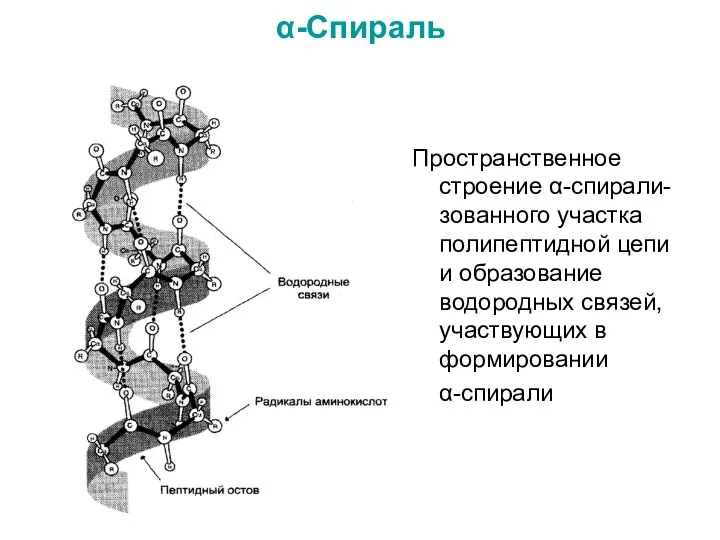
α-Спираль
Пространственное строение α-спирали-зованного участка полипептидной цепи и образование водородных связей, участвующих
α-Спираль
Пространственное строение α-спирали-зованного участка полипептидной цепи и образование водородных связей, участвующих
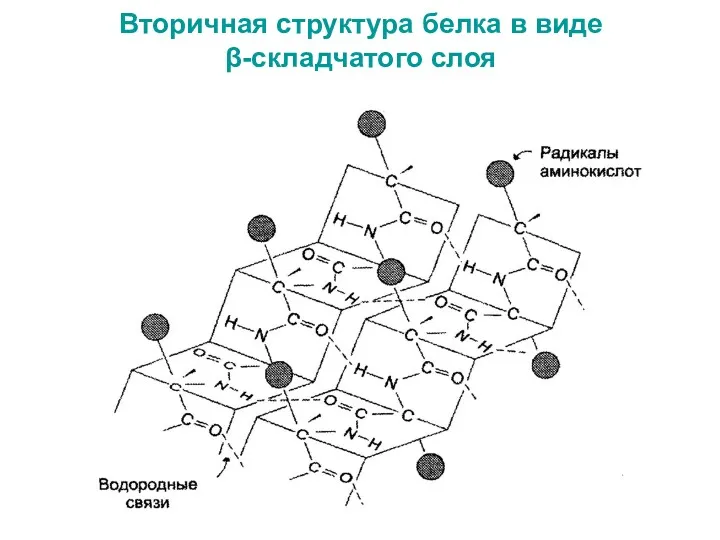
Вторичная структура белка в виде
β-складчатого слоя
Вторичная структура белка в виде
β-складчатого слоя
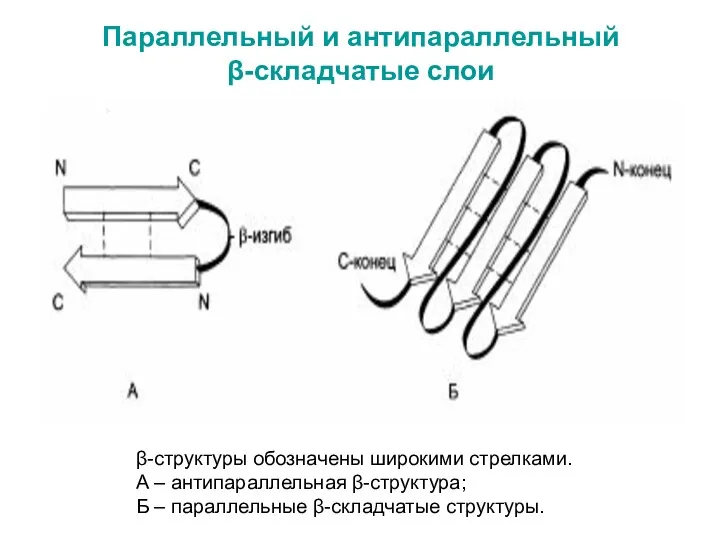
Параллельный и антипараллельный
β-складчатые слои
β-структуры обозначены широкими стрелками.
А – антипараллельная β-структура;
Б
Параллельный и антипараллельный
β-складчатые слои
β-структуры обозначены широкими стрелками.
А – антипараллельная β-структура;
Б

Супервторичная (надвторичная) структура белков
Специфический порядок формирования вторичной структуры, содержащий сочетания элементов
Супервторичная (надвторичная) структура белков
Специфический порядок формирования вторичной структуры, содержащий сочетания элементов

Классификация супервторичных структур («структурные мотивы»)
«α-спираль-поворот-α-спираль» (ДНК-связывающий белок);
«структура β-бочонка» (домен пируваткиназы, триозофосфатизомераза);
«лейциновая
Классификация супервторичных структур («структурные мотивы»)
«α-спираль-поворот-α-спираль» (ДНК-связывающий белок);
«структура β-бочонка» (домен пируваткиназы, триозофосфатизомераза);
«лейциновая
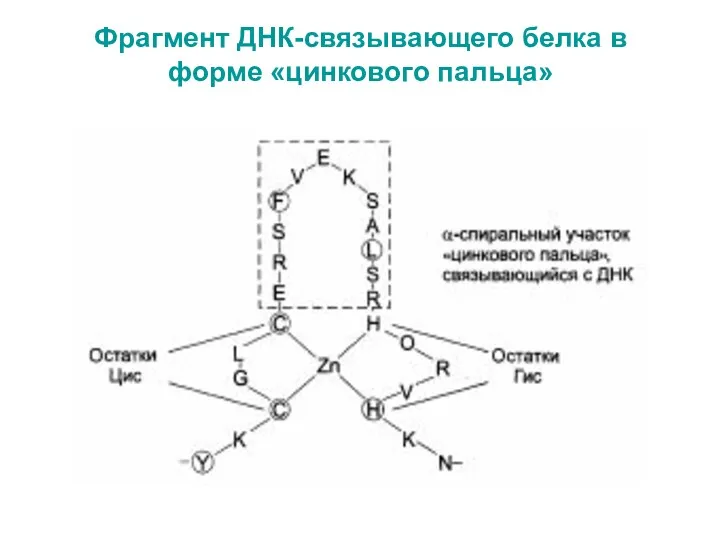
Фрагмент ДНК-связывающего белка в форме «цинкового пальца»
Фрагмент ДНК-связывающего белка в форме «цинкового пальца»

Если полипептидная цепь белка содержит более 200 аминокислот, то, как правило,
Если полипептидная цепь белка содержит более 200 аминокислот, то, как правило,
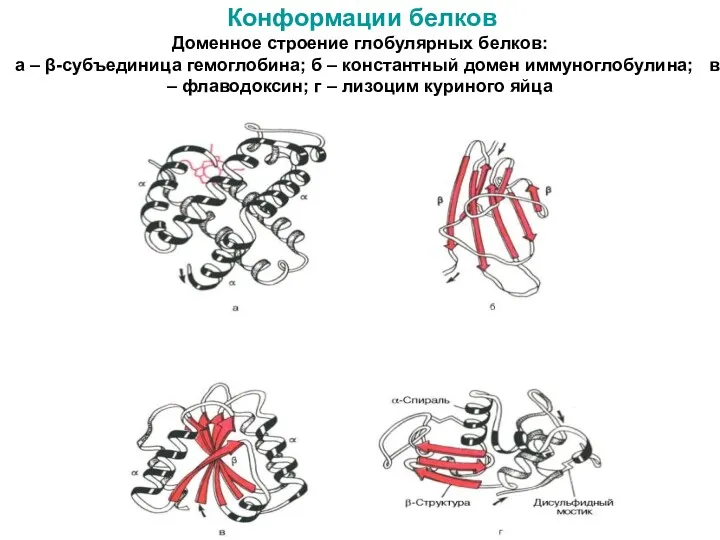
Конформации белков
Доменное строение глобулярных белков:
а – β-субъединица гемоглобина; б
Конформации белков Доменное строение глобулярных белков: а – β-субъединица гемоглобина; б

Конформации белков
Третичная структура белков – трехмерная пространственная структура, образующаяся за счет
Конформации белков
Третичная структура белков – трехмерная пространственная структура, образующаяся за счет

Связи, принимающие участие в формировании третичной структуры белков
Связи, принимающие участие в формировании третичной структуры белков
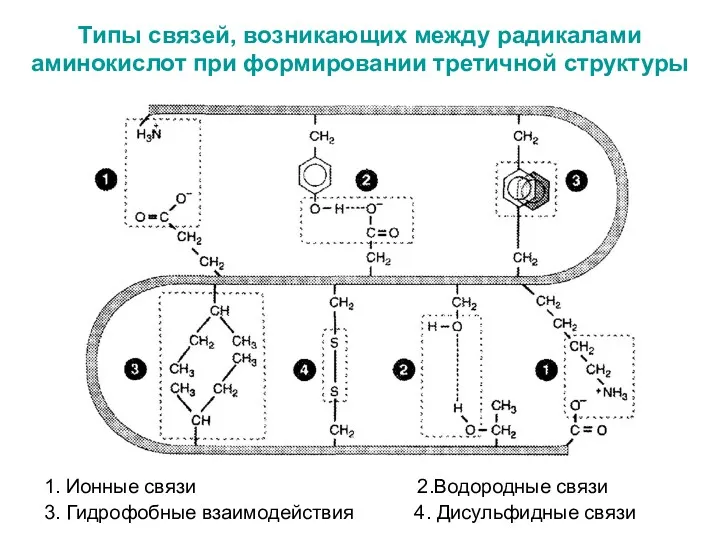
Типы связей, возникающих между радикалами аминокислот при формировании третичной структуры
1. Ионные
Типы связей, возникающих между радикалами аминокислот при формировании третичной структуры
1. Ионные

Связи, принимающие участие в формировании третичной структуры белков
Гидрофобные радикалы аминокислот стремятся
Связи, принимающие участие в формировании третичной структуры белков
Гидрофобные радикалы аминокислот стремятся

Связи, принимающие участие в формировании третичной структуры белков
Ионные связи могут возникать
Связи, принимающие участие в формировании третичной структуры белков
Ионные связи могут возникать
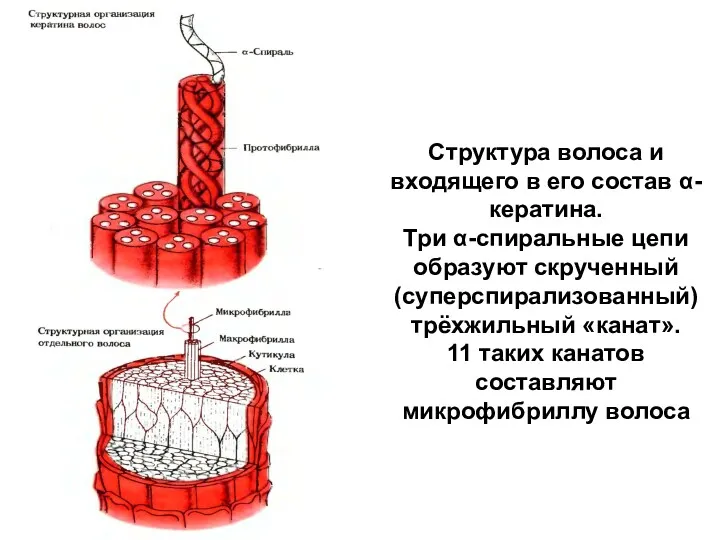
Структура волоса и входящего в его состав α-кератина.
Три α-спиральные цепи образуют
Структура волоса и входящего в его состав α-кератина. Три α-спиральные цепи образуют
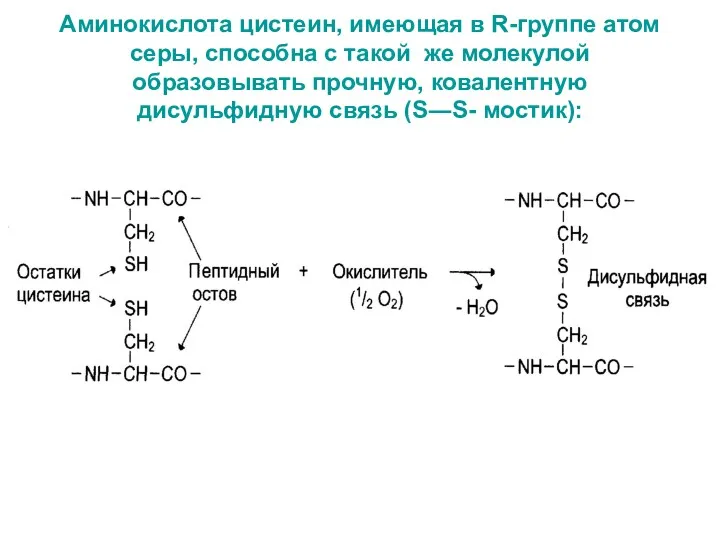
Аминокислота цистеин, имеющая в R-группе атом серы, способна с такой же
Аминокислота цистеин, имеющая в R-группе атом серы, способна с такой же
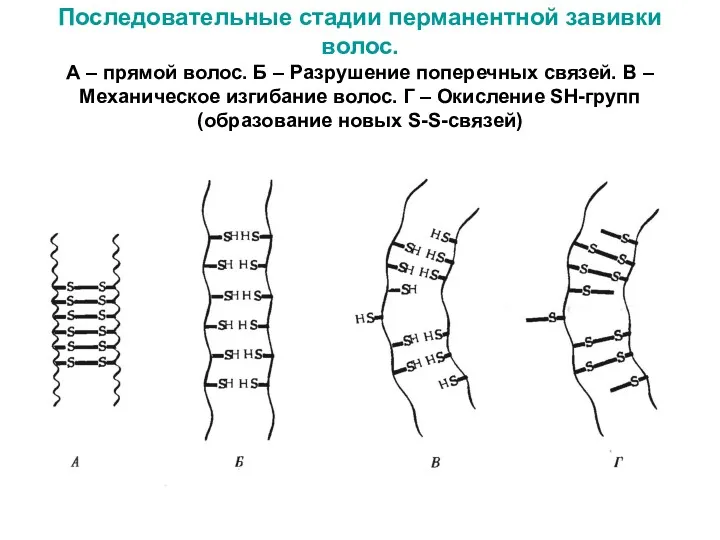
Последовательные стадии перманентной завивки волос.
А – прямой волос. Б – Разрушение
Последовательные стадии перманентной завивки волос. А – прямой волос. Б – Разрушение
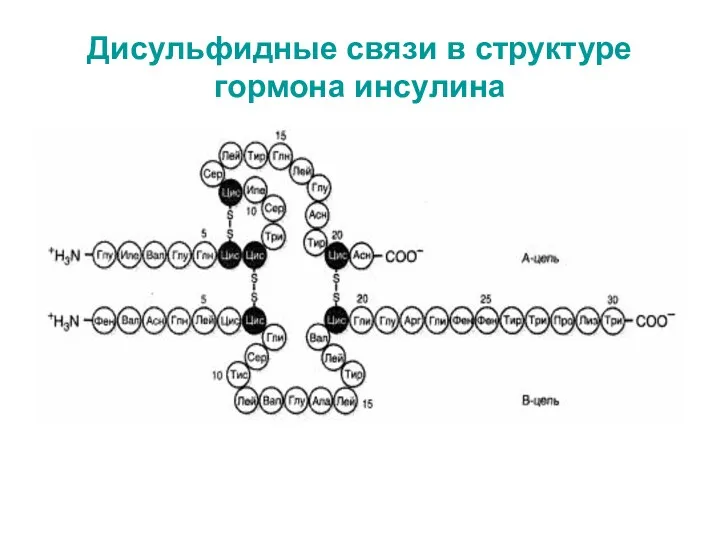
Дисульфидные связи в структуре гормона инсулина
Дисульфидные связи в структуре гормона инсулина

Все белки с одинаковой первичной структурой, находящиеся в одинаковых условиях, приобретают
Все белки с одинаковой первичной структурой, находящиеся в одинаковых условиях, приобретают

Конформации белков
Количество и взаиморасположение полипептидных цепей в пространстве называют «четвертичная структура
Конформации белков
Количество и взаиморасположение полипептидных цепей в пространстве называют «четвертичная структура
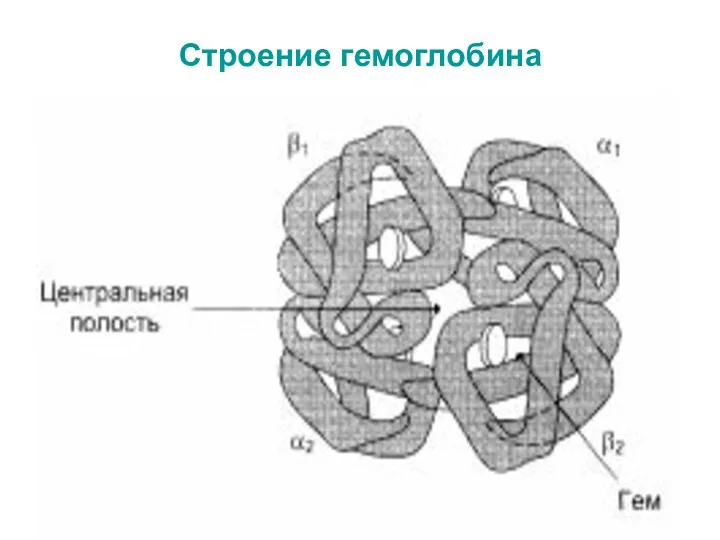
Строение гемоглобина
Строение гемоглобина
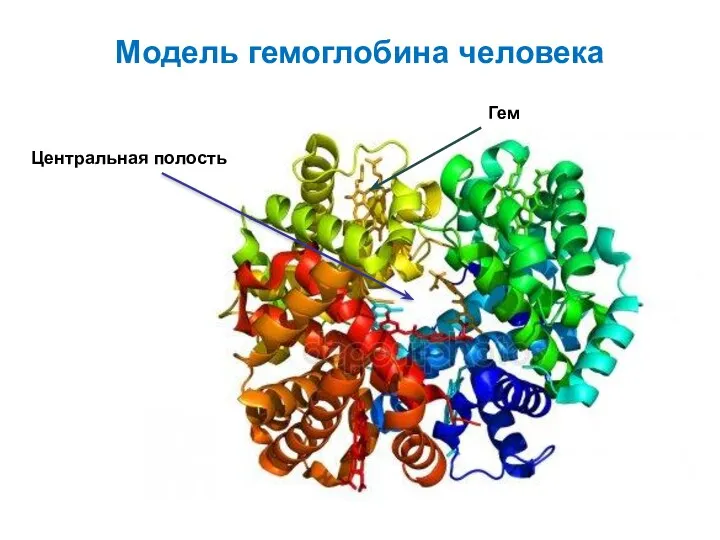
Модель гемоглобина человека
Центральная полость
Гем
Модель гемоглобина человека
Центральная полость
Гем

5. Взаимосвязь структуры и функции. Денатурация и ренатурация
5. Взаимосвязь структуры и функции. Денатурация и ренатурация

Денатурация – изменение нативной структуры белка под действием различных факторов (химические
Денатурация – изменение нативной структуры белка под действием различных факторов (химические
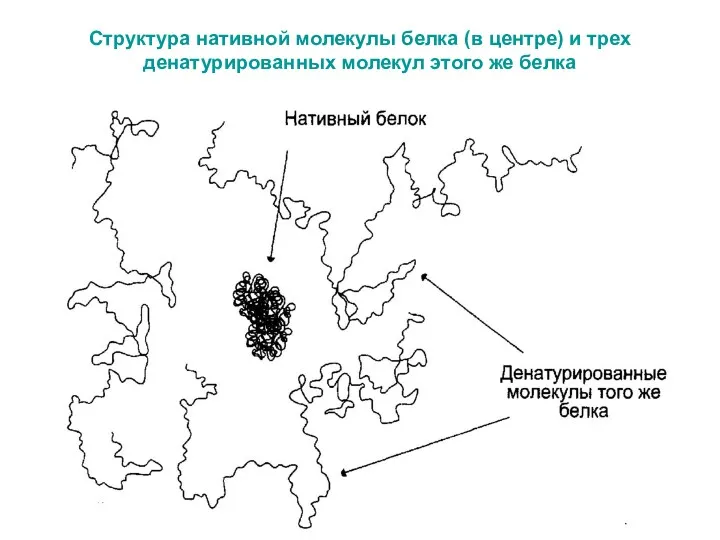
Структура нативной молекулы белка (в центре) и трех денатурированных молекул этого
Структура нативной молекулы белка (в центре) и трех денатурированных молекул этого
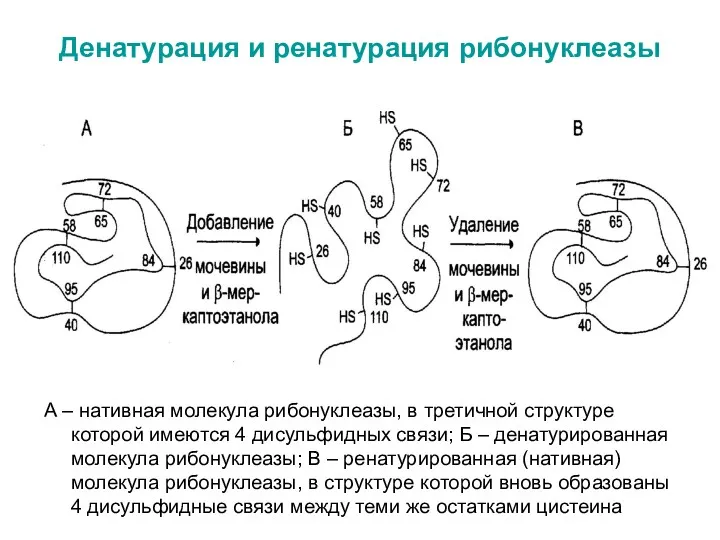
Денатурация и ренатурация рибонуклеазы
А – нативная молекула рибонуклеазы, в третичной структуре
Денатурация и ренатурация рибонуклеазы
А – нативная молекула рибонуклеазы, в третичной структуре

Формирование трёхмерной структуры белка в клетке
Формирование трёхмерной структуры белка в клетке
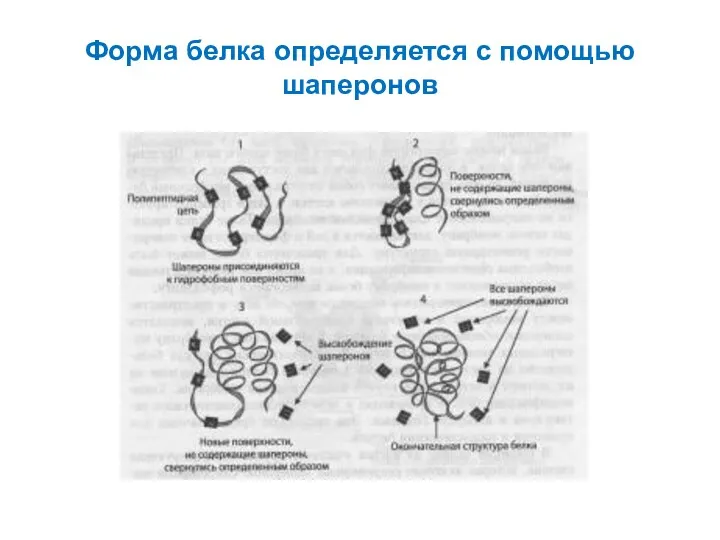
Форма белка определяется с помощью шаперонов
Форма белка определяется с помощью шаперонов

Болезни, связанные с нарушением фолдинга белков
Некоторые растворимые в воде белки при
Болезни, связанные с нарушением фолдинга белков
Некоторые растворимые в воде белки при

6. Функции белков: структурная, каталитическая, транспортная, рецепторная, регуляторная, защитная, сократительная и
6. Функции белков: структурная, каталитическая, транспортная, рецепторная, регуляторная, защитная, сократительная и

Классификации белков
По выполняемым функциям:
ферменты (катализаторы – каталаза, амилаза и др.),
регуляторные
Классификации белков
По выполняемым функциям:
ферменты (катализаторы – каталаза, амилаза и др.),
регуляторные

7. Свойства простых белков. Гистоны, альбумины. Структурные белки: тубулины, кератины, коллаген,
7. Свойства простых белков. Гистоны, альбумины. Структурные белки: тубулины, кератины, коллаген,

Простые белки при гидролизе распадаются только на свободные аминокислоты.
Простые белки делятся
Простые белки при гидролизе распадаются только на свободные аминокислоты. Простые белки делятся

Гистоны – являются белками основного характера, растворимы в солевых растворах.
Гистоны выполняют
Гистоны – являются белками основного характера, растворимы в солевых растворах.
Гистоны выполняют
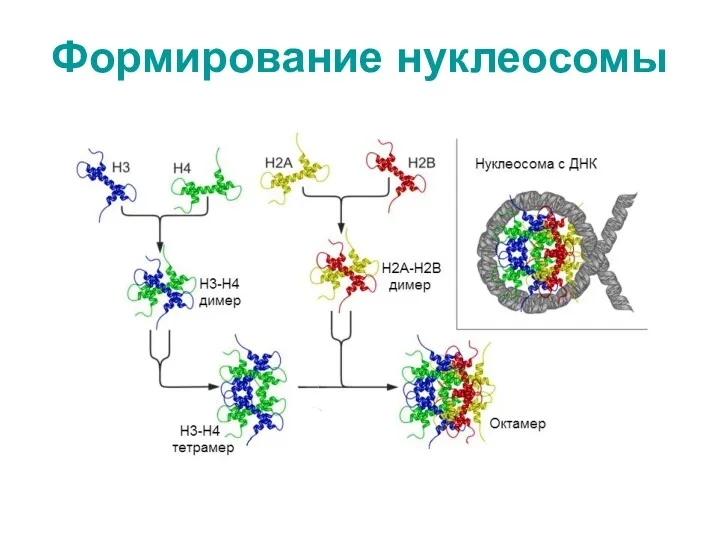
Формирование нуклеосомы
Формирование нуклеосомы

Альбумины (от лат. albumen, род. падеж albuminis – белок) – растворимы
Альбумины (от лат. albumen, род. падеж albuminis – белок) – растворимы

Кератины – семейство фибриллярных белков, обладающих механической прочностью, которая среди материалов
Кератины – семейство фибриллярных белков, обладающих механической прочностью, которая среди материалов

Тубулин – основной белок микротрубочек, обладающий сократительными функциями; димер, состоящий из
Тубулин – основной белок микротрубочек, обладающий сократительными функциями; димер, состоящий из

Коллаген – составляет 25-33% от общего количества белка организма взрослого человека,
Коллаген – составляет 25-33% от общего количества белка организма взрослого человека,

Эластин – основной белковый компонент, из которого состоят эластические волокна.
При
Эластин – основной белковый компонент, из которого состоят эластические волокна. При
 Тип Кишечнополостные. Тип Гребневики. Жизненные циклы кишечнополостных
Тип Кишечнополостные. Тип Гребневики. Жизненные циклы кишечнополостных The endangered animals
The endangered animals Пищеварение в ротовой полости и желудке
Пищеварение в ротовой полости и желудке Постельные клопы
Постельные клопы Мочеполовой аппарат. Органы мочеотделения. (Лекция 5)
Мочеполовой аппарат. Органы мочеотделения. (Лекция 5) Педсовет Современные подходы к преподаванию в условиях ФГОС
Педсовет Современные подходы к преподаванию в условиях ФГОС Размножение организмов. (Лекция 5)
Размножение организмов. (Лекция 5) Оснащение микробиологической лаборатории
Оснащение микробиологической лаборатории Кора большого мозга
Кора большого мозга Венозная система. Лекция № 19
Венозная система. Лекция № 19 Лабораторная диагностика нарушений обмена белков плазмы крови (Преальбумин, С-РБ, Фибронектин, Фибриноген)
Лабораторная диагностика нарушений обмена белков плазмы крови (Преальбумин, С-РБ, Фибронектин, Фибриноген) Теории происхождения жизни. (Лекция 15)
Теории происхождения жизни. (Лекция 15) Становление наук о человеке
Становление наук о человеке Рябина обыкновенная
Рябина обыкновенная Строение сердечно-сосудистой системы
Строение сердечно-сосудистой системы Қызыл балдырлардың түрлерінің жалпы
Қызыл балдырлардың түрлерінің жалпы Различия в строении клеток эукариот и прокариот
Различия в строении клеток эукариот и прокариот Объекты дендрологии. Понятие о жизненной форме у древесных растений
Объекты дендрологии. Понятие о жизненной форме у древесных растений Мышцы туловища, головы, шеи
Мышцы туловища, головы, шеи презентация Орган слуха
презентация Орган слуха Різноманітність ссавців
Різноманітність ссавців Высшие жирные кислоты. Липиды
Высшие жирные кислоты. Липиды Вирусы, вирусология.
Вирусы, вирусология. Обмін білків
Обмін білків Симпатический отдел автономной нервной системы
Симпатический отдел автономной нервной системы Биотические связи
Биотические связи Углеводы. 9 класс
Углеводы. 9 класс Строение клетки растения и животного
Строение клетки растения и животного