- Анализ взаимодействий in vitro. Стратегия, от белка к гену. (Лекция 6)

Содержание
- 2. Стратегия: от белка к гену Изолировать белок на основе его функциональной активности (например, энзиматической или гормональной)
- 3. Определение последовательности аминокислот с помощью метода деградации по Эдману фенилизотиоцианат гидролиз Цикл 1 Цикл 2 Идентификация
- 4. Стратегия: от гена к белку Изолировать геномный клон, соответствующий измененному белку в мутанте (например, ауксотрофия, наследственные
- 5. Взаимодействия двух белков in vivo in vitro центрифугирование хроматография ко-иммунопреципитация Blot-overlay Pull-down Surface Plasmon Resonance бесклеточные
- 6. Электрофорез белков в полиакриламидном геле Определение содержания белка во фракциях Выделение белка для последующего анализа Оценка
- 7. Двумерный электрофорез – максимальное разрешение белков в геле - - + + без SDS, но в
- 8. 10 мкг белка 14С-метка 825 часов экспозиции 1100 белков E. сoli, разделенные с помощью двумерного электрофореза
- 9. Окрашивание белков в геле и на мембране SDS-гель перенос на мембрану Ag (0,1-1 ng) Coomassie (1-2
- 10. Масс-спектрометрия MALDI-TOF Matrix-Assisted Laser Desorption/Ionization - Time Of Flight UV лазер (330-360 nm ) катод анод
- 11. белковое пятно вырезают из геля белок переваривают трипсином; смесь пептидов вносят в масс-спектрометр и получают спектр
- 12. отдельный пик белка, очищенного хроматографией пептиды, полученные обработкой очищенного белка трипсином первый масс-спектр дает массы пептидов
- 13. Центрифугирование: угловой ротор сила гравитации в ультрацентрифуге Fs=mω r супернатант осадок Type 45 Ti 6 x
- 14. градиент плотности отверстие частицы возрастающей плотности сила гравитации TLS-55 4 x 2.2 mL 6 x 39
- 15. Аналитическое ультрацентрифугирование S – константа седиментации (сек) 10 сек = 1 S (Svedberg) -13 постоянная скорость
- 16. Дифференциальное осаждение гомогенат целые клетки ядра элементы цитоскелета митохондрии лизосомы хлоропласты пероксисомы микросомы мелкие везикулы рибосомы
- 17. Наиболее популярные детергенты неионные ионные Triton X-100 Octylglucoside (-)Sodium Deoxycholate (-)Sodium Dodecylsulfate (+)Цетавлон = Cetyltrimethylammonium bromide
- 18. Детергенты растворяют гидрофобные белки детергентно-липидные мицеллы Детергент Детергентно-белковые мицеллы Детергентные мицеллы Фрагментация Липидно-белковая мембрана ККМ -
- 19. Центрифугирование в градиенте плотности Седиментация: непрерывный линейный градиент формируется в процессе центрифугирования Плавучая плотность ДНК в
- 20. Анализ фракций градиента номера фракций OD280 концентрация сахарозы Концентрация сахарозы Концентрация белка Энзиматическая активность Белковый состав
- 21. Пример: флотация микротрубочек (from Fuchs and Westermann) 1 3 5 смесь митохондрий и микротрубочек Anti-Nkin2 Предварительная
- 22. Гель-фильтрация Аффинная хроматография Ионообменная хроматография Виды хроматографии антитело пористая гранула заряженная гранула Na+ Cl- DEAE-группа CM-группа
- 23. Pull-down анализ: связывание с рекомбинантным белком, синтезированным в бактериях в бактериях аффинный домен аффинный домен N
- 24. Ко-иммунопреципитация: нужны антитела к обоим партнерам bead А А А А центрифугирование и промывание bead А
- 25. Пример: взаимодействие двух секретируемых белков ДНК, кодирующая FLAG-белок А ДНК, кодирующая Myc-белок В Иммунопреципитация антителами к
- 26. белки разделяются электрофорезом и переносятся из геля на мембрану инкубация с белком-«наживкой» инкубация с антителами против
- 27. Surface Plasmon Resonance: анализ взаимодействия очищенных белков или белков и нуклеиновых кислот призма лазер камера поляризатор
- 28. Surface Plasmon Resonance: анализ взаимодействия антиген-антитело
- 29. Быстрая идентификация продукта гена Локализация мутаций посредством синтеза укороченных продуктов Включение модифицированных неприродных аминокислот для структурных
- 30. Обычно используемые системы 1. Экстракт ретикулоцитов кролика. Ретикулоцит – предшественник эритроцита, он уже не имеет ядра.
- 31. Пример: синтез люциферазы светлячка в бесклеточной системе Вопрос: происходит ли при синтезе в бесклеточной системе E.
- 32. Черные столбики – активность люциферазы Серые столбики – активность люциферазы после добавления пуромицина (from Kolb V.
- 33. Лекция 7 Взаимодействия in vivo
- 34. Электрофорез в неденатурирующих условиях Химическая сшивка Иммунопреципитация хроматина Колокализация в клетке Pull-down из клеточного лизата Дигибридная
- 35. Электрофорез белков в неденатурирующих условиях 1. Получение белка в очищенном нативном виде 2. Определение свойств выделенного
- 36. Фото-активируемые аналоги аминокислот L-Photo-Leucine и L-Photo-methionine – аналоги нормальных аминокислот; присутствуя в среде, они включаются в
- 37. Химические сшивки белков С О Н Н Простейший способ – «сшить» формальдегидом: в водной среде НО
- 38. Химическая сшивка белка с ДНК – иммунопреципитация хроматина: идентификация на ДНК сайта связывания белка Х химически
- 39. кДНК эукариотический вектор, кодирующий белок + таг или репортер клонирование экспрессия Экспрессия трансгена в клетках эукариот
- 41. Доставка крупных молекул в клетки Микроинъекция: ДНК, мРНК, белок Липосомная трансфекция: ДНК, мРНК Са-фосфатная трансфекция: ДНК
- 42. таг-1 таг-2 Ко-экспрессия Иммунопреципитация антителами против тага-1 Иммуноблоттинг преципитата с антителами против тага-2 Ко-экспрессия возможных партнеров:
- 43. Ко-экспрессия возможных партнеров: ко-локализация Перераспределение зиксина в ответ на экспрессию фактора Xanf-1 (from Martynova et al.)
- 44. фрагмент белка А, связывающий иммуноглобулины белок, специфически связывающий калмодулин в присутствии ионов Са2+ В клетке: клеточный
- 46. Скачать презентацию

Стратегия: от белка к гену
Изолировать белок на основе его функциональной активности
Стратегия: от белка к гену
Изолировать белок на основе его функциональной активности
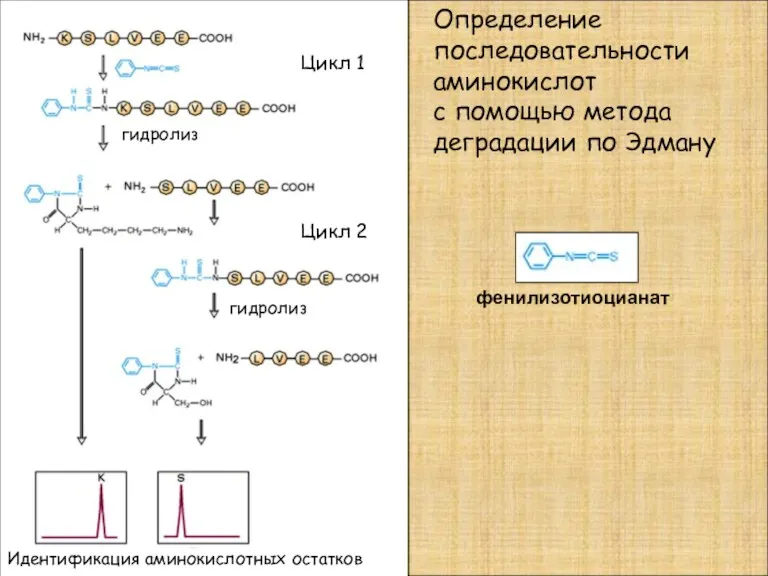
Определение
последовательности
аминокислот
с помощью метода
деградации по Эдману
фенилизотиоцианат
гидролиз
Цикл 1
Цикл 2
Идентификация аминокислотных
Определение
последовательности
аминокислот
с помощью метода
деградации по Эдману
фенилизотиоцианат
гидролиз
Цикл 1
Цикл 2
Идентификация аминокислотных

Стратегия: от гена к белку
Изолировать геномный клон, соответствующий измененному белку в
Стратегия: от гена к белку
Изолировать геномный клон, соответствующий измененному белку в
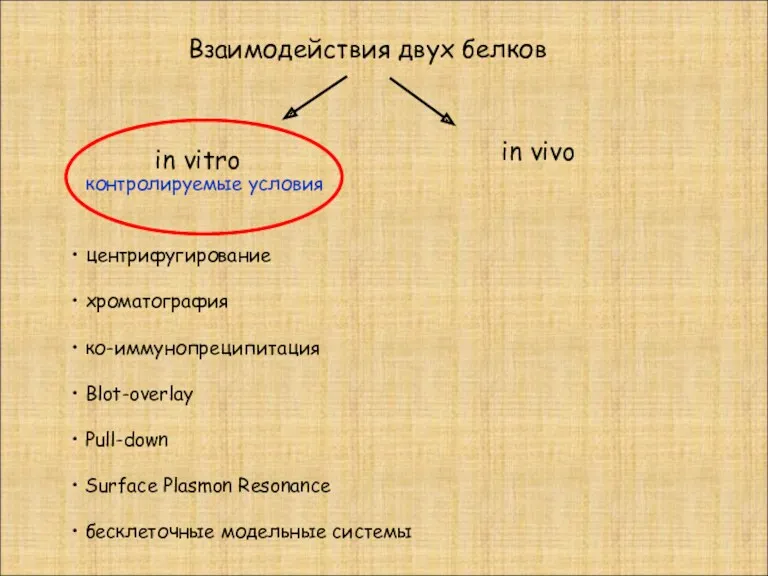
Взаимодействия двух белков
in vivo
in vitro
центрифугирование
хроматография
ко-иммунопреципитация
Blot-overlay
Pull-down
Surface
Взаимодействия двух белков
in vivo
in vitro
центрифугирование
хроматография
ко-иммунопреципитация
Blot-overlay
Pull-down
Surface
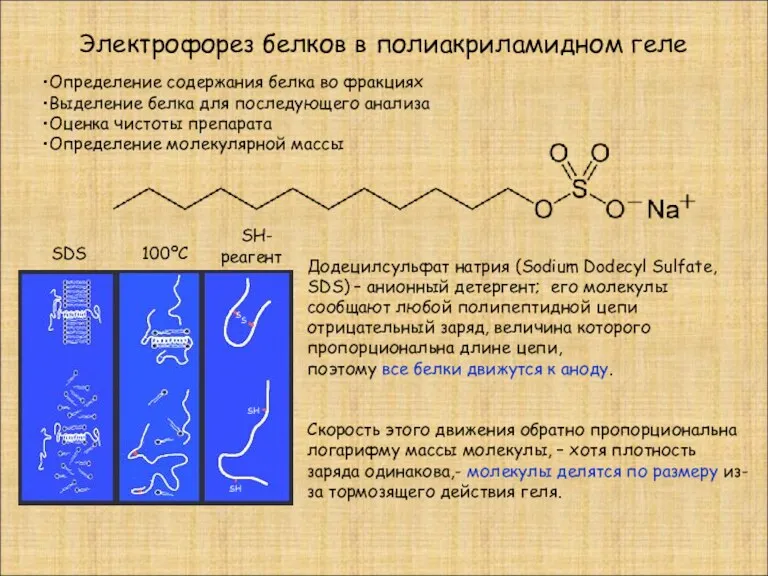
Электрофорез белков в полиакриламидном геле
Определение содержания белка во фракциях
Выделение белка
Электрофорез белков в полиакриламидном геле
Определение содержания белка во фракциях
Выделение белка
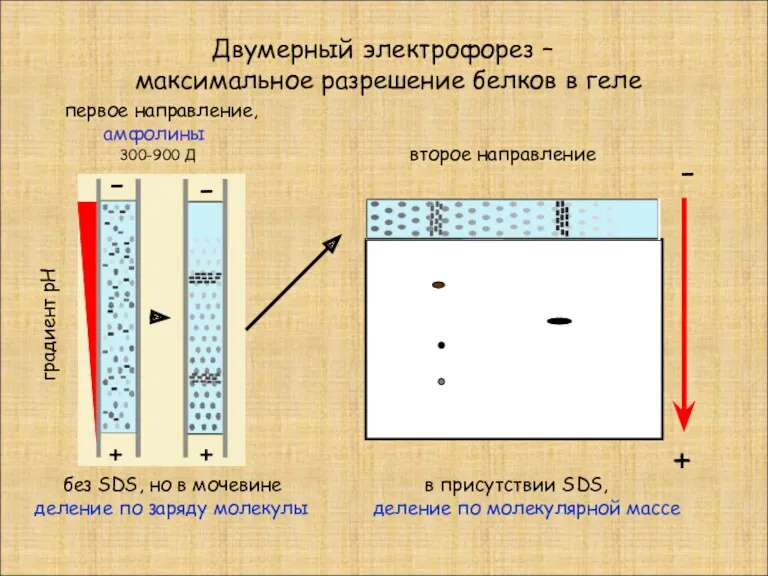
Двумерный электрофорез –
максимальное разрешение белков в геле
-
Двумерный электрофорез –
максимальное разрешение белков в геле
-
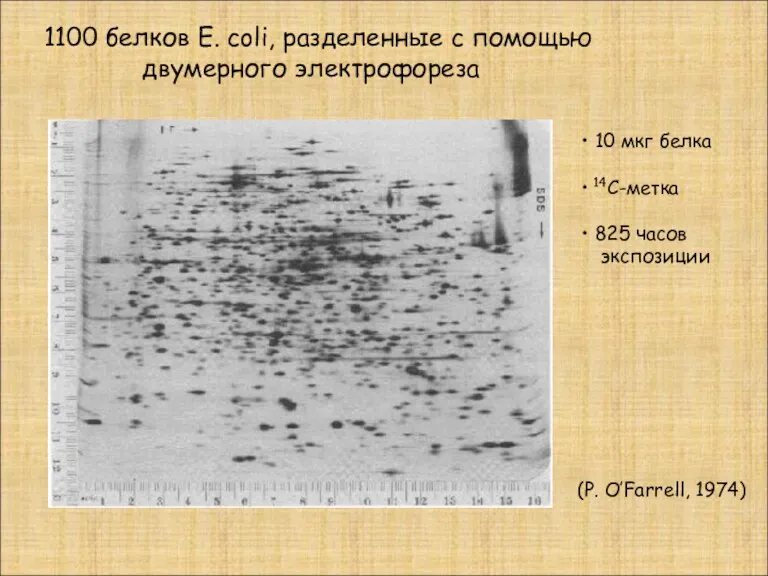
10 мкг белка
14С-метка
825 часов
экспозиции
1100 белков E.
10 мкг белка
14С-метка
825 часов
экспозиции
1100 белков E.
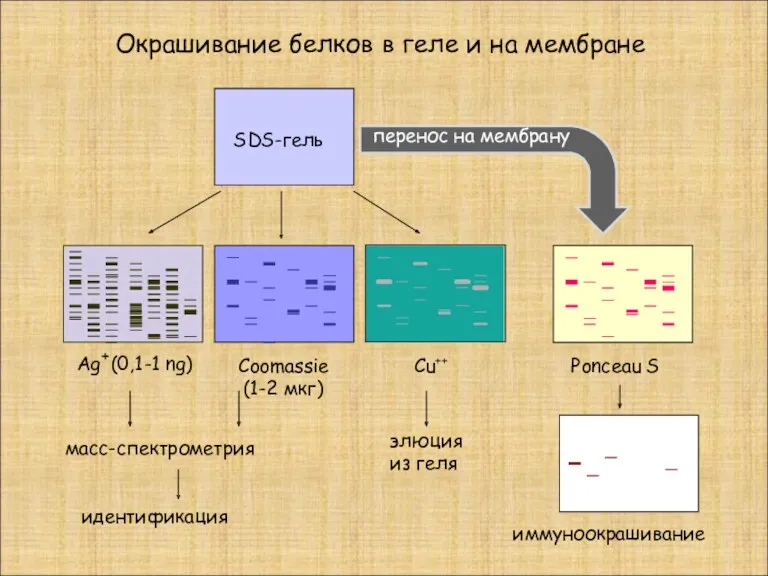
Окрашивание белков в геле и на мембране
SDS-гель
перенос на мембрану
Окрашивание белков в геле и на мембране
SDS-гель
перенос на мембрану
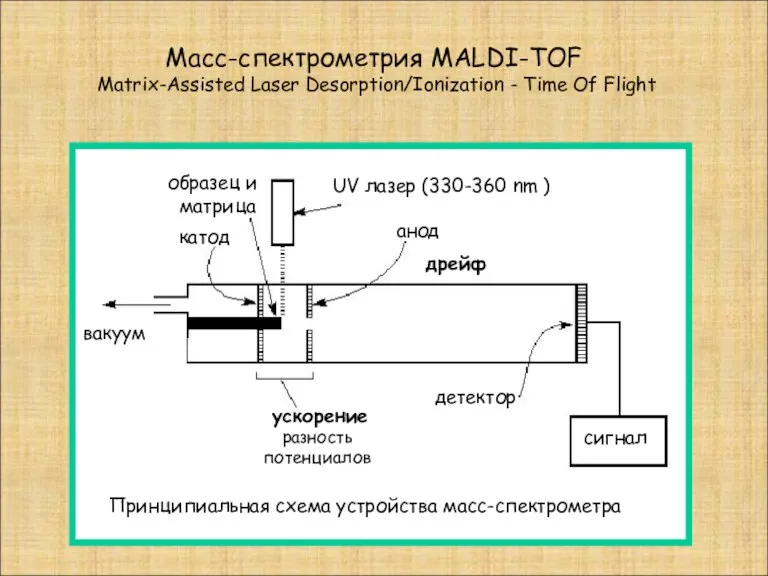
Масс-спектрометрия MALDI-TOF
Matrix-Assisted Laser Desorption/Ionization - Time Of Flight
UV лазер
Масс-спектрометрия MALDI-TOF
Matrix-Assisted Laser Desorption/Ionization - Time Of Flight
UV лазер
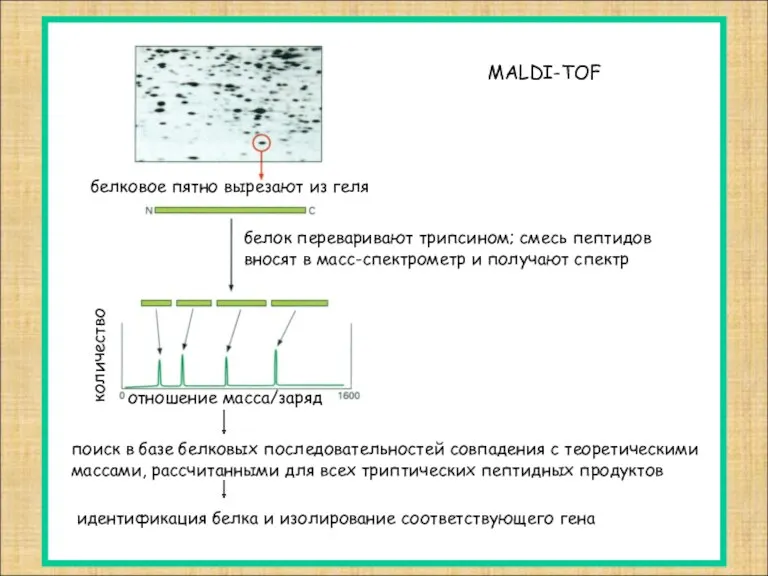
белковое пятно вырезают из геля
белок переваривают трипсином; смесь пептидов
вносят в
белковое пятно вырезают из геля
белок переваривают трипсином; смесь пептидов
вносят в
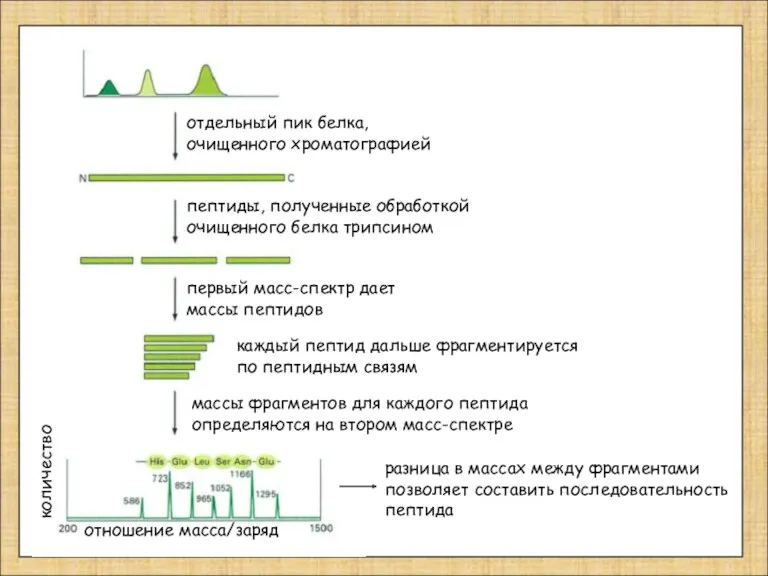
отдельный пик белка,
очищенного хроматографией
пептиды, полученные обработкой
очищенного белка трипсином
первый масс-спектр
отдельный пик белка,
очищенного хроматографией
пептиды, полученные обработкой
очищенного белка трипсином
первый масс-спектр
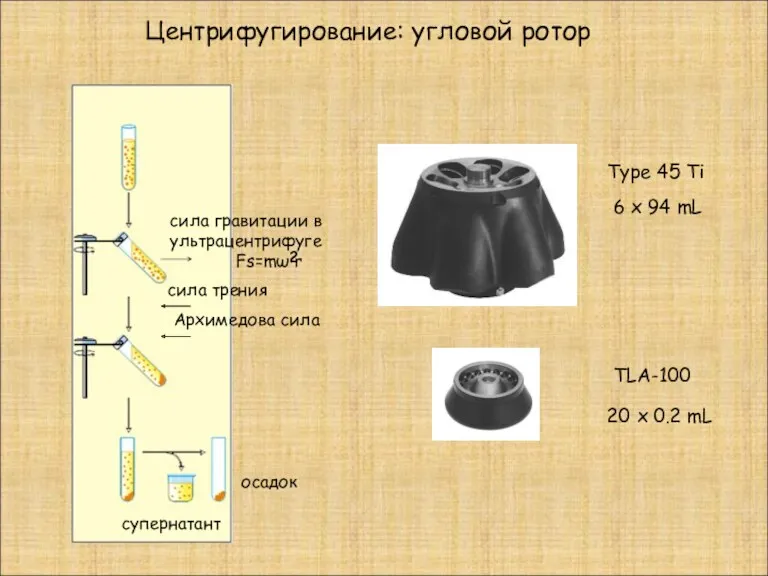
Центрифугирование: угловой ротор
сила гравитации в
ультрацентрифуге
Fs=mω r
супернатант
осадок
Type 45
Центрифугирование: угловой ротор
сила гравитации в
ультрацентрифуге
Fs=mω r
супернатант
осадок
Type 45
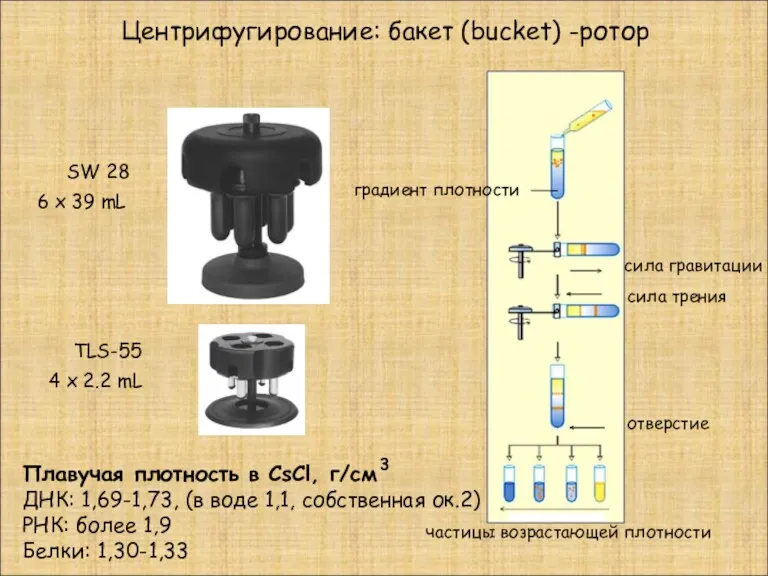
градиент плотности
отверстие
частицы возрастающей плотности
сила гравитации
TLS-55
4 x 2.2 mL
6 x
градиент плотности
отверстие
частицы возрастающей плотности
сила гравитации
TLS-55
4 x 2.2 mL
6 x
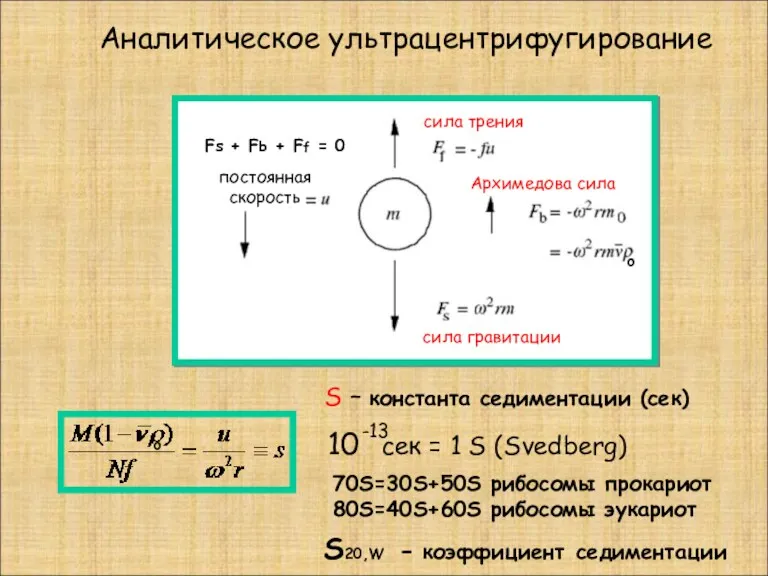
Аналитическое ультрацентрифугирование
S – константа седиментации (сек)
10 сек = 1 S
Аналитическое ультрацентрифугирование
S – константа седиментации (сек)
10 сек = 1 S
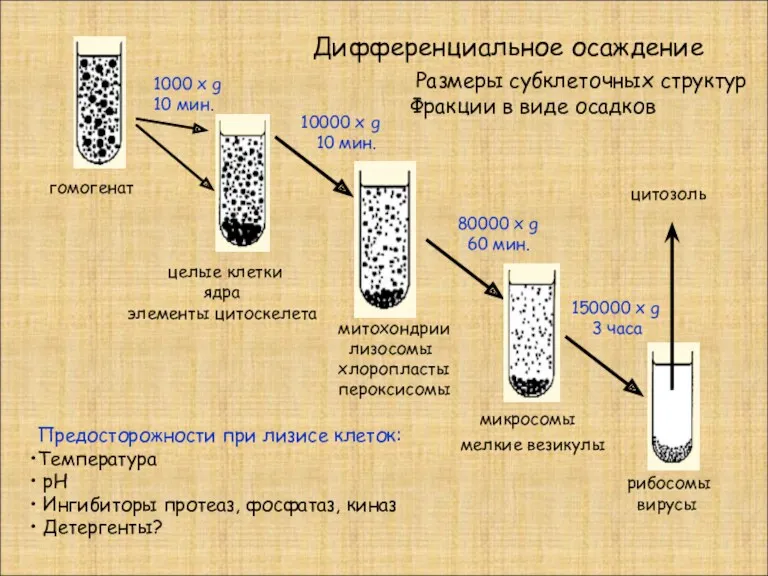
Дифференциальное осаждение
гомогенат
целые клетки
ядра
элементы цитоскелета
митохондрии
лизосомы
хлоропласты
пероксисомы
микросомы
мелкие везикулы
рибосомы
вирусы
Дифференциальное осаждение
гомогенат
целые клетки
ядра
элементы цитоскелета
митохондрии
лизосомы
хлоропласты
пероксисомы
микросомы
мелкие везикулы
рибосомы
вирусы
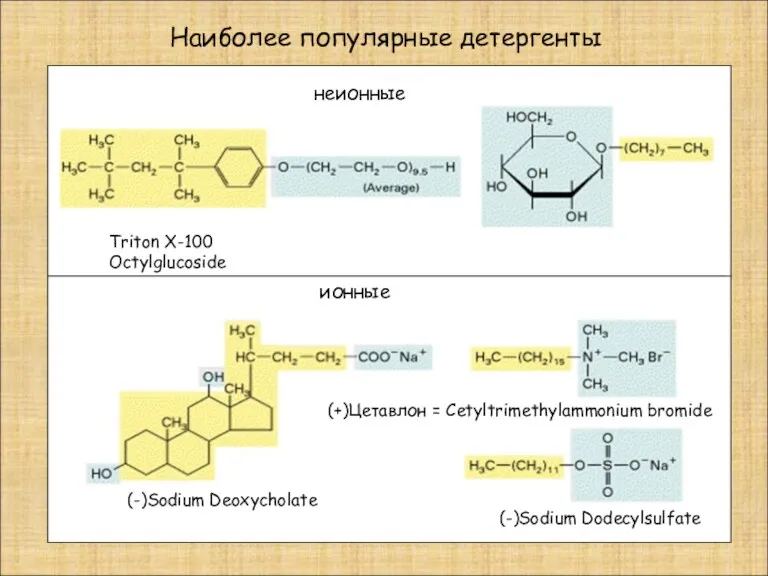
Наиболее популярные детергенты
неионные
ионные
Triton X-100 Octylglucoside
(-)Sodium Deoxycholate
(-)Sodium Dodecylsulfate
(+)Цетавлон = Cetyltrimethylammonium bromide
Наиболее популярные детергенты
неионные
ионные
Triton X-100 Octylglucoside
(-)Sodium Deoxycholate
(-)Sodium Dodecylsulfate
(+)Цетавлон = Cetyltrimethylammonium bromide
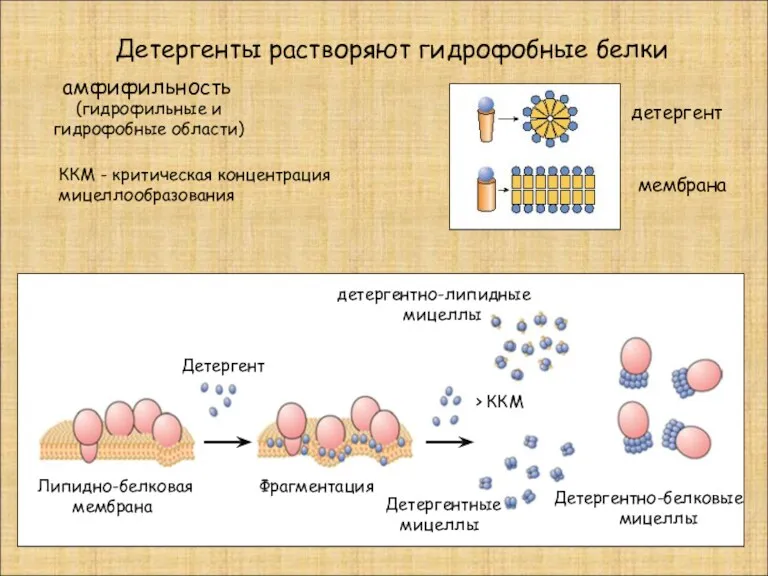
Детергенты растворяют гидрофобные белки
детергентно-липидные
мицеллы
Детергент
Детергентно-белковые
мицеллы
Детергентные
мицеллы
Фрагментация
Липидно-белковая
мембрана
ККМ - критическая
Детергенты растворяют гидрофобные белки
детергентно-липидные
мицеллы
Детергент
Детергентно-белковые
мицеллы
Детергентные
мицеллы
Фрагментация
Липидно-белковая
мембрана
ККМ - критическая
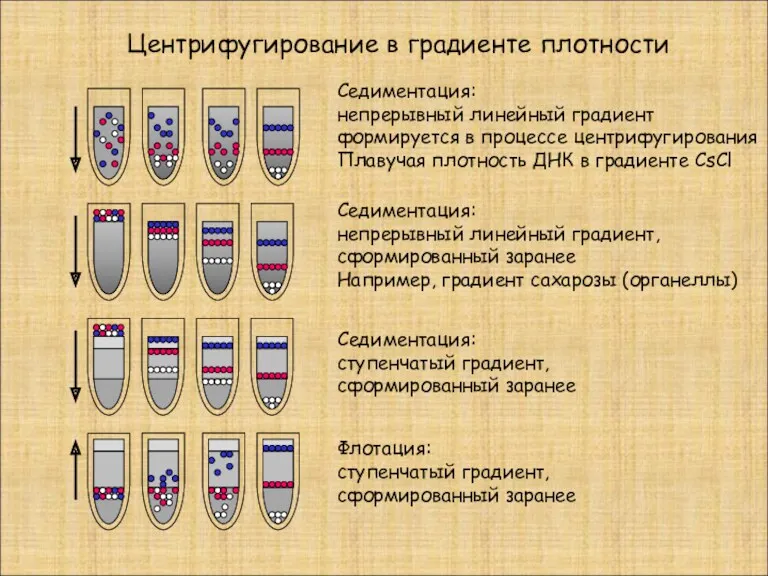
Центрифугирование в градиенте плотности
Седиментация:
непрерывный линейный градиент
формируется в процессе центрифугирования
Плавучая плотность
Центрифугирование в градиенте плотности
Седиментация:
непрерывный линейный градиент
формируется в процессе центрифугирования
Плавучая плотность
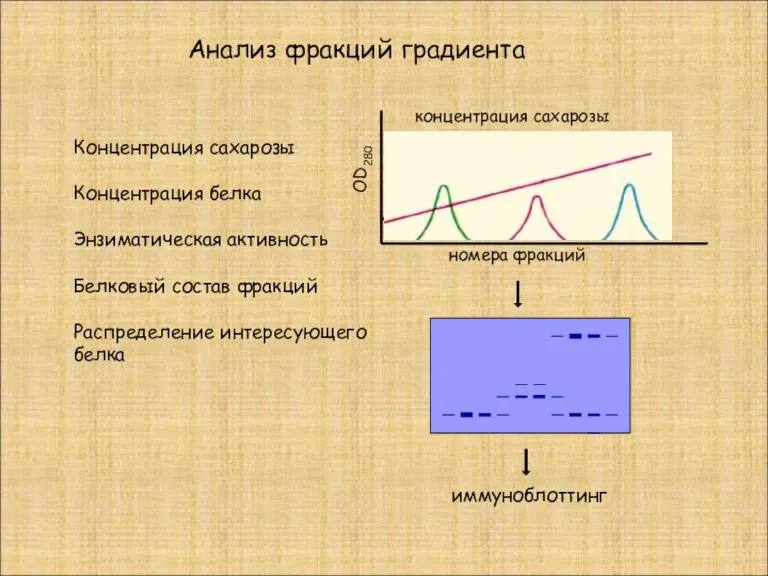
Анализ фракций градиента
номера фракций
OD280
концентрация сахарозы
Концентрация сахарозы
Концентрация белка
Энзиматическая активность
Белковый состав фракций
Распределение интересующего
Анализ фракций градиента
номера фракций
OD280
концентрация сахарозы
Концентрация сахарозы
Концентрация белка
Энзиматическая активность
Белковый состав фракций
Распределение интересующего
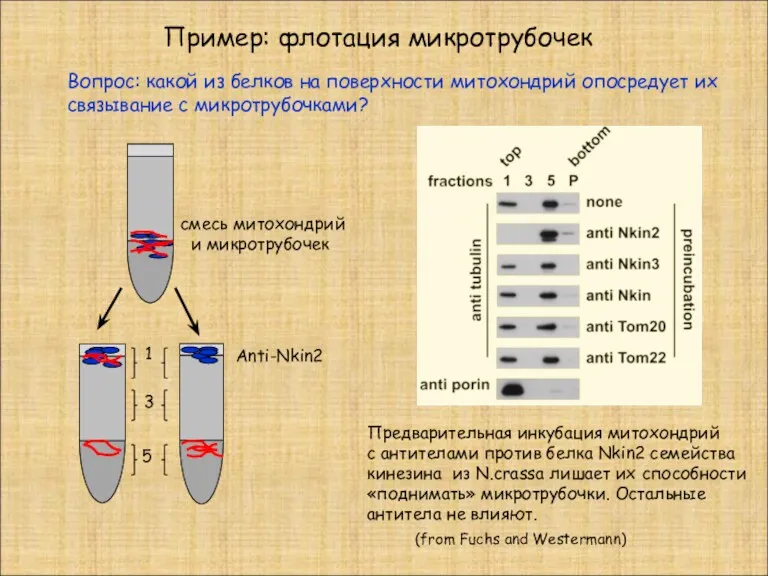
Пример: флотация микротрубочек
(from Fuchs and Westermann)
1
3
5
смесь митохондрий
и
Пример: флотация микротрубочек
(from Fuchs and Westermann)
1
3
5
смесь митохондрий
и
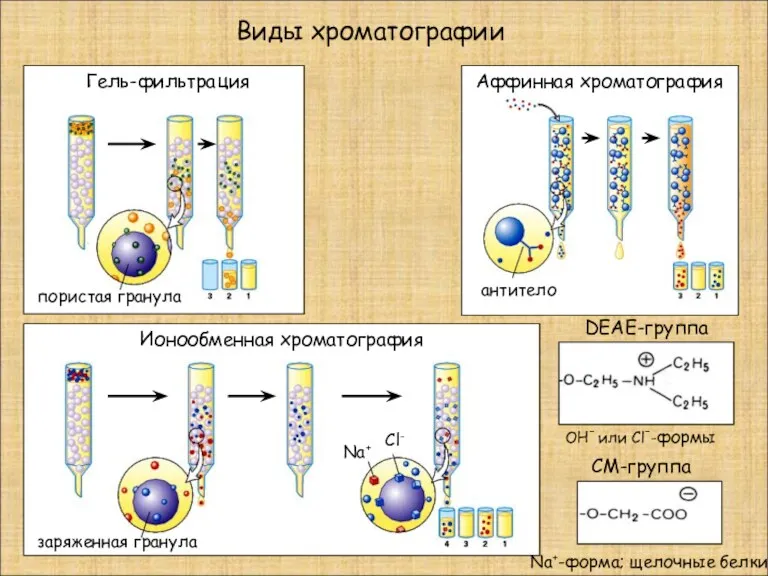
Гель-фильтрация
Аффинная хроматография
Ионообменная хроматография
Виды хроматографии
антитело
пористая гранула
заряженная гранула
Na+
Cl-
DEAE-группа
CM-группа
Na+-форма; щелочные белки
OH- или Cl--формы
Гель-фильтрация
Аффинная хроматография
Ионообменная хроматография
Виды хроматографии
антитело
пористая гранула
заряженная гранула
Na+
Cl-
DEAE-группа
CM-группа
Na+-форма; щелочные белки
OH- или Cl--формы
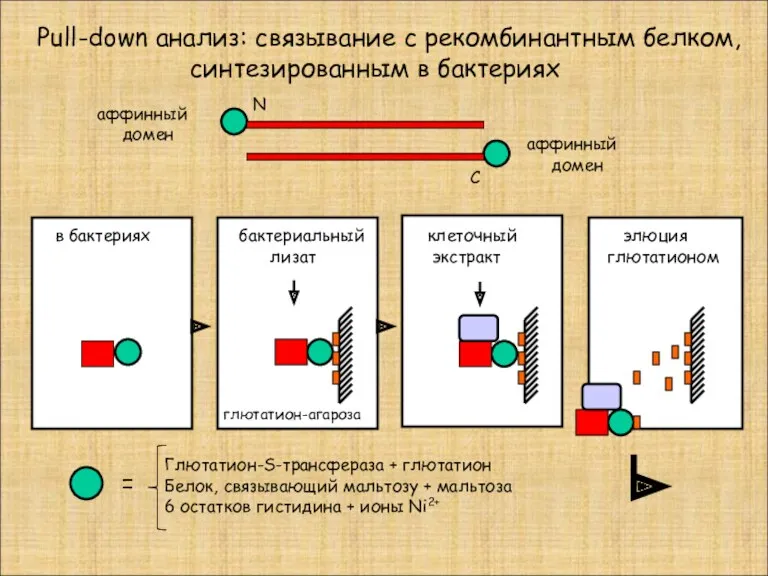
Pull-down анализ: связывание с рекомбинантным белком,
синтезированным в бактериях
в бактериях
аффинный
Pull-down анализ: связывание с рекомбинантным белком,
синтезированным в бактериях
в бактериях
аффинный
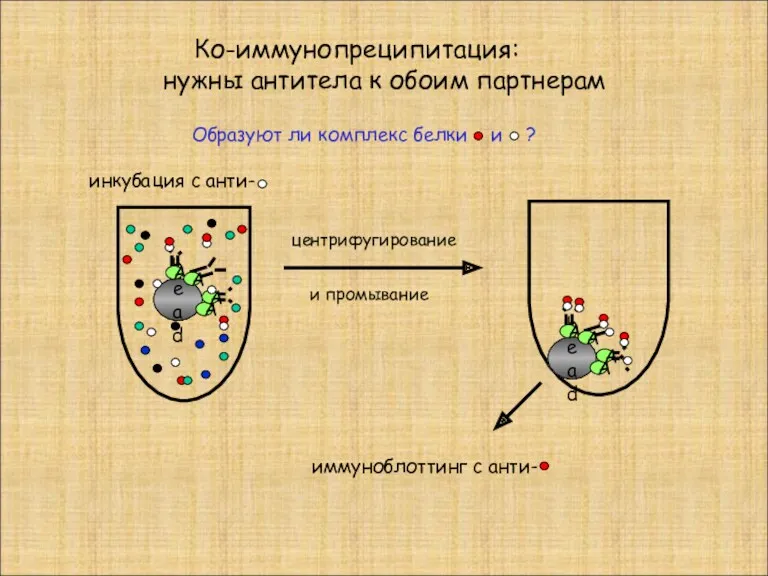
Ко-иммунопреципитация:
нужны антитела к обоим партнерам
bead
А
А
А
А
центрифугирование
и промывание
bead
А
А
А
А
инкубация с анти-
иммуноблоттинг с
Ко-иммунопреципитация:
нужны антитела к обоим партнерам
bead
А
А
А
А
центрифугирование
и промывание
bead
А
А
А
А
инкубация с анти-
иммуноблоттинг с
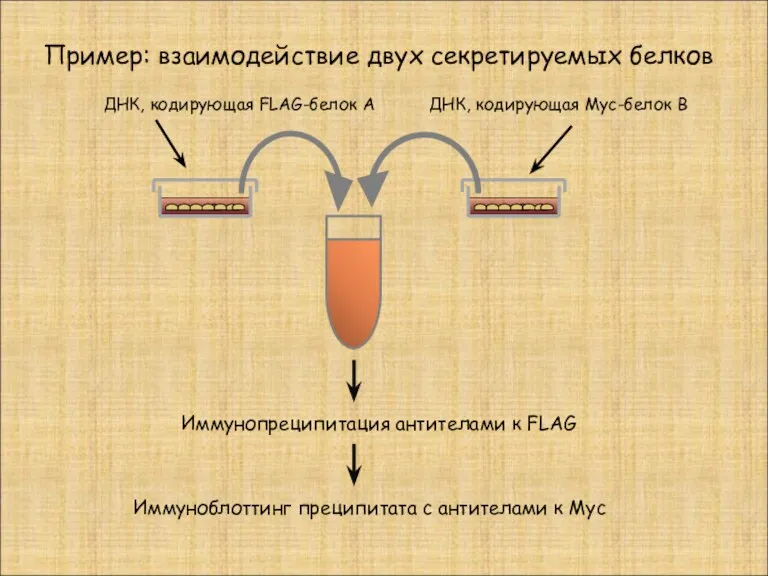
Пример: взаимодействие двух секретируемых белков
ДНК, кодирующая FLAG-белок А
ДНК, кодирующая Myc-белок В
Иммунопреципитация
Пример: взаимодействие двух секретируемых белков
ДНК, кодирующая FLAG-белок А
ДНК, кодирующая Myc-белок В
Иммунопреципитация
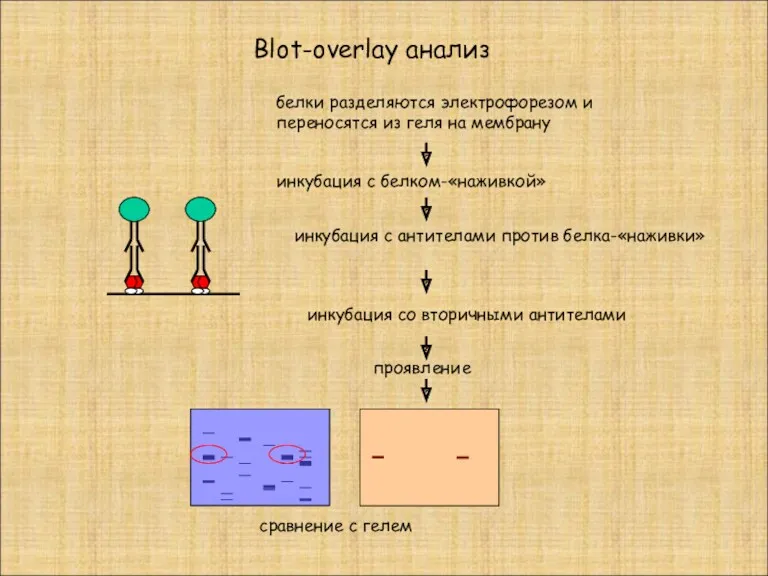
белки разделяются электрофорезом и переносятся из геля на мембрану
инкубация с белком-«наживкой»
инкубация
белки разделяются электрофорезом и переносятся из геля на мембрану
инкубация с белком-«наживкой»
инкубация
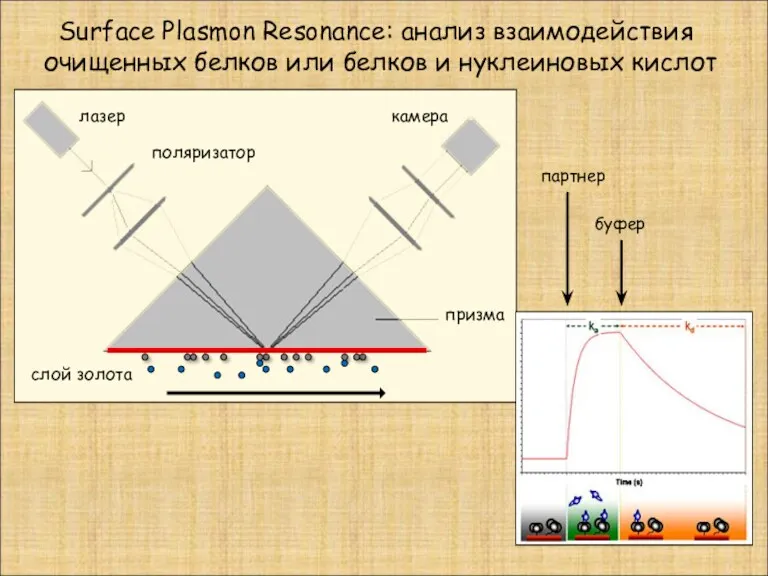
Surface Plasmon Resonance: анализ взаимодействия очищенных белков или белков и
Surface Plasmon Resonance: анализ взаимодействия очищенных белков или белков и
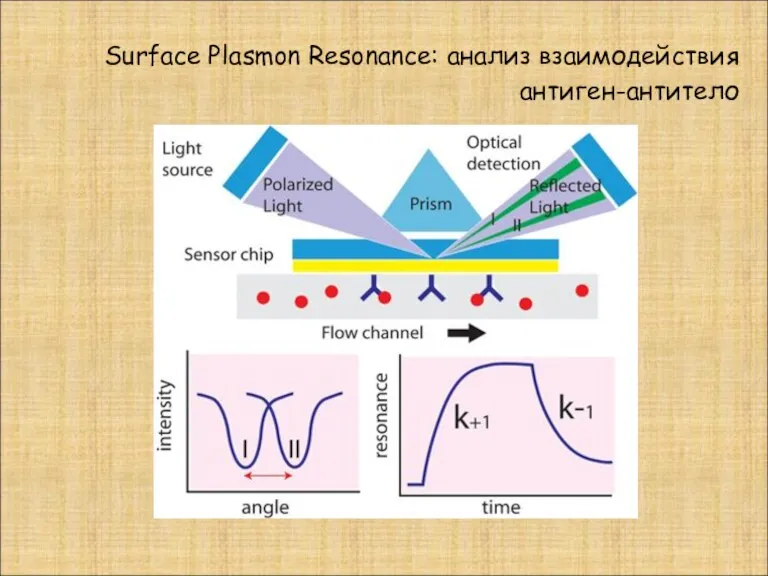
Surface Plasmon Resonance: анализ взаимодействия антиген-антитело
Surface Plasmon Resonance: анализ взаимодействия антиген-антитело
Быстрая идентификация продукта гена
Локализация мутаций посредством
синтеза укороченных продуктов
Включение модифицированных
Локализация мутаций посредством
синтеза укороченных продуктов
Включение модифицированных

Обычно используемые системы
1. Экстракт ретикулоцитов кролика.
Ретикулоцит – предшественник эритроцита, он
Обычно используемые системы
1. Экстракт ретикулоцитов кролика.
Ретикулоцит – предшественник эритроцита, он
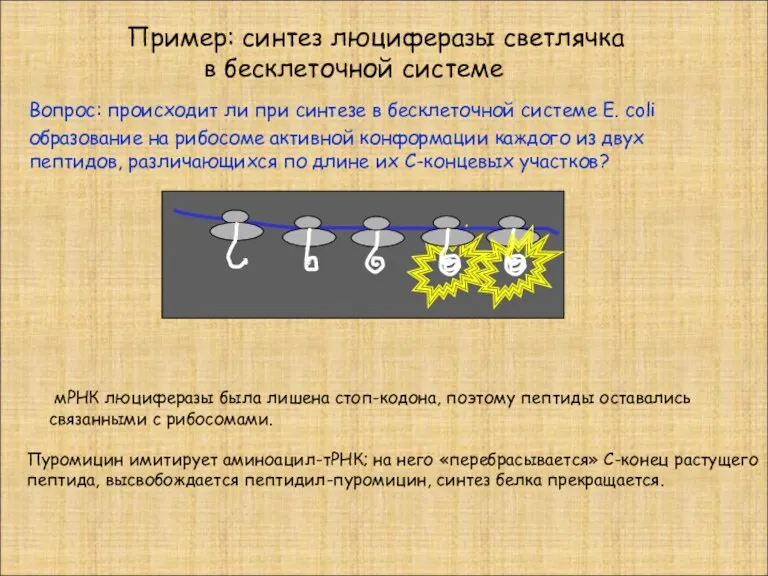
Пример: синтез люциферазы светлячка
в бесклеточной системе
Вопрос: происходит ли при
Пример: синтез люциферазы светлячка
в бесклеточной системе
Вопрос: происходит ли при
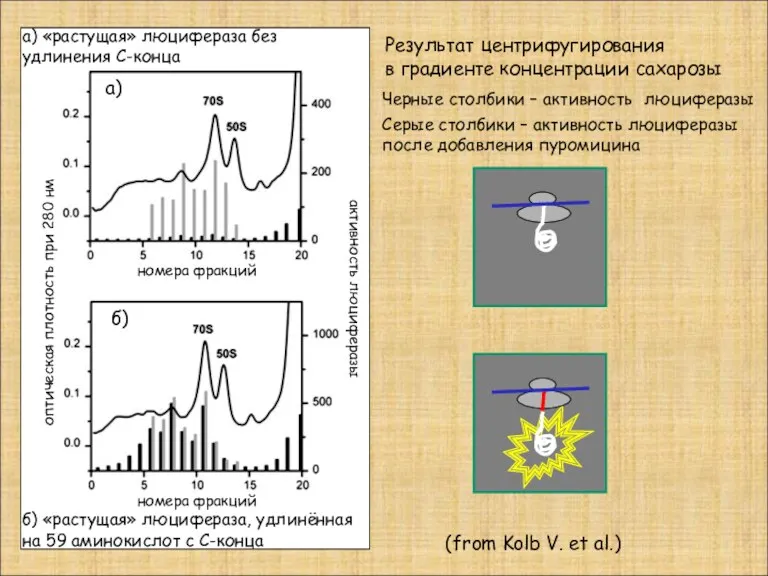
Черные столбики – активность люциферазы
Серые столбики – активность люциферазы
после
Черные столбики – активность люциферазы
Серые столбики – активность люциферазы
после

Лекция 7
Взаимодействия in vivo
Лекция 7
Взаимодействия in vivo

Электрофорез в неденатурирующих условиях
Химическая сшивка
Иммунопреципитация хроматина
Колокализация в клетке
Электрофорез в неденатурирующих условиях
Химическая сшивка
Иммунопреципитация хроматина
Колокализация в клетке

Электрофорез белков в неденатурирующих условиях
1. Получение белка в очищенном нативном
Электрофорез белков в неденатурирующих условиях
1. Получение белка в очищенном нативном
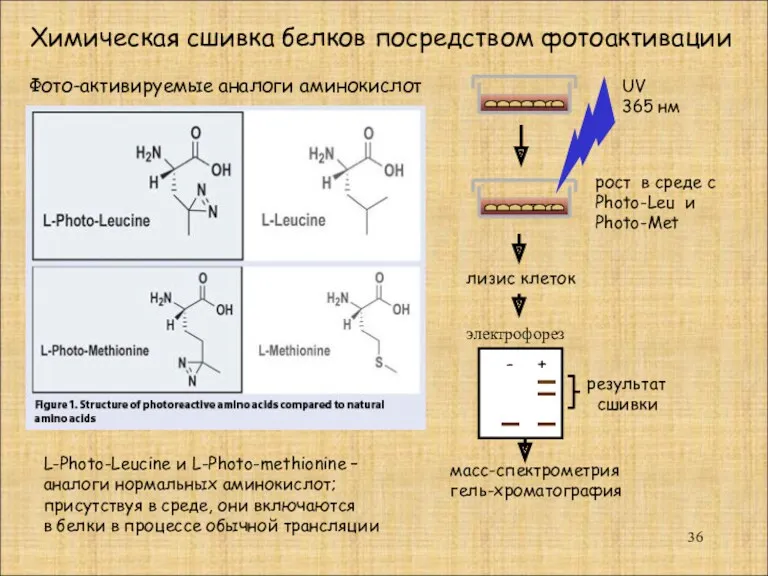
Фото-активируемые аналоги аминокислот
L-Photo-Leucine и L-Photo-methionine –
аналоги нормальных аминокислот;
присутствуя в среде,
Фото-активируемые аналоги аминокислот
L-Photo-Leucine и L-Photo-methionine –
аналоги нормальных аминокислот;
присутствуя в среде,
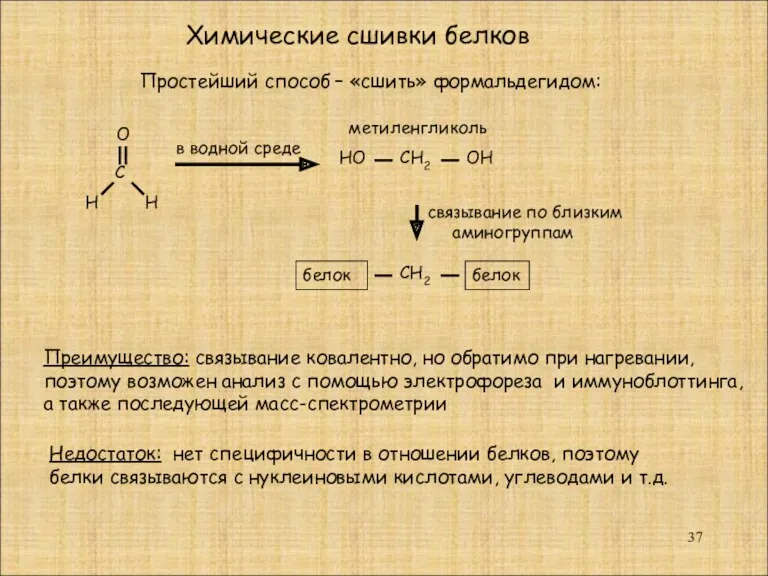
Химические сшивки белков
С
О
Н
Н
Простейший способ – «сшить» формальдегидом:
в водной среде
НО
СН2
ОН
белок
белок
СН2
метиленгликоль
Преимущество:
Химические сшивки белков
С
О
Н
Н
Простейший способ – «сшить» формальдегидом:
в водной среде
НО
СН2
ОН
белок
белок
СН2
метиленгликоль
Преимущество:
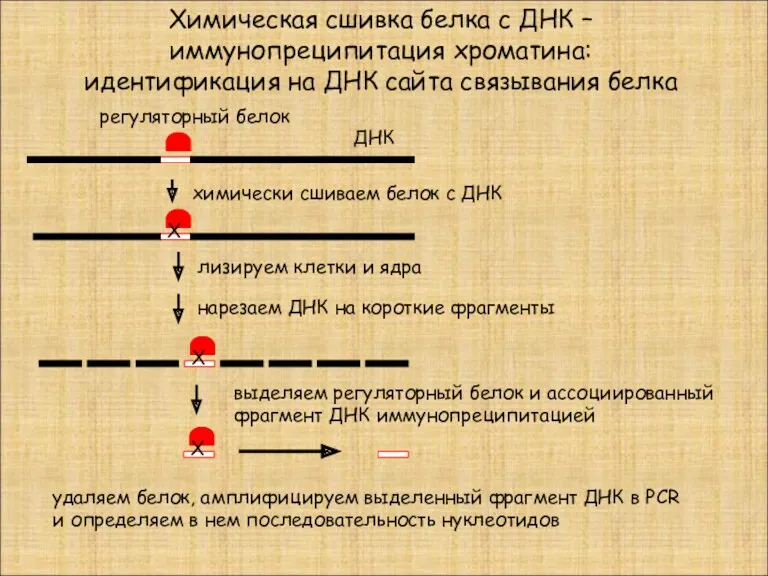
Химическая сшивка белка с ДНК –
иммунопреципитация хроматина:
идентификация на ДНК сайта
Химическая сшивка белка с ДНК –
иммунопреципитация хроматина:
идентификация на ДНК сайта
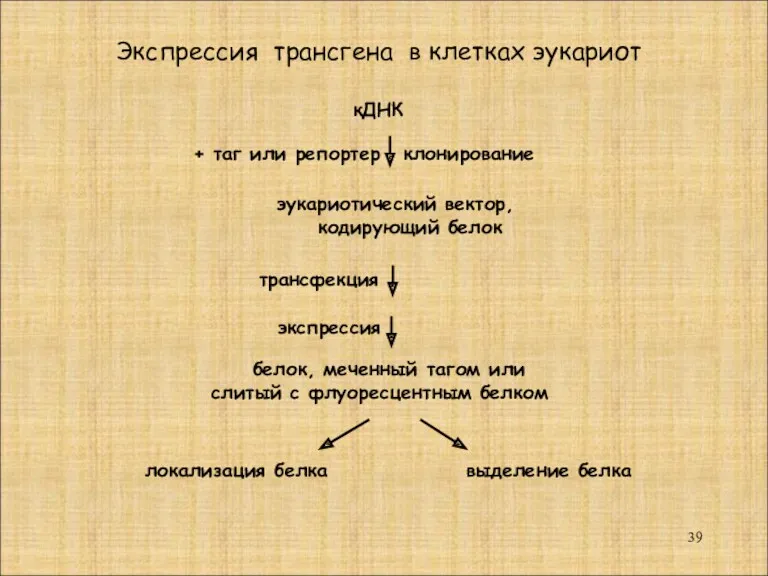
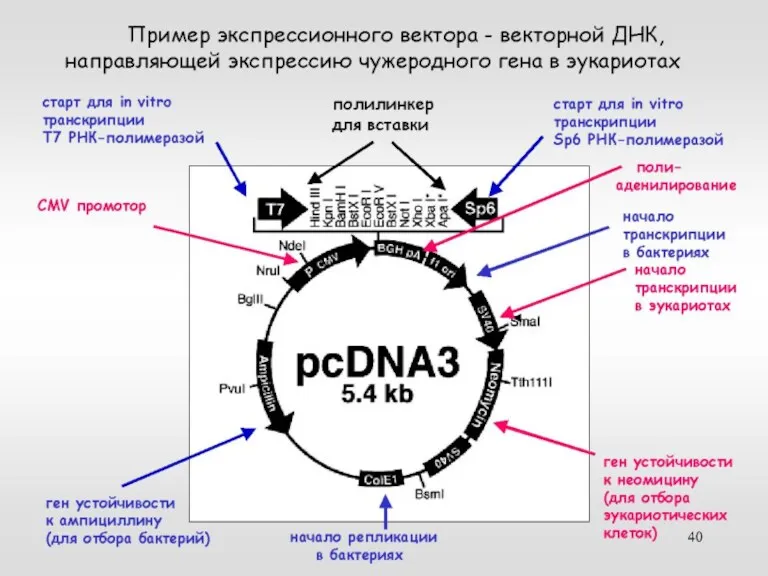
кДНК
эукариотический вектор,
кодирующий белок
+ таг или репортер
клонирование
экспрессия
Экспрессия трансгена в
кДНК
эукариотический вектор,
кодирующий белок
+ таг или репортер
клонирование
экспрессия
Экспрессия трансгена в

Доставка крупных молекул в клетки
Микроинъекция: ДНК, мРНК, белок
Липосомная трансфекция: ДНК,
Доставка крупных молекул в клетки
Микроинъекция: ДНК, мРНК, белок
Липосомная трансфекция: ДНК,
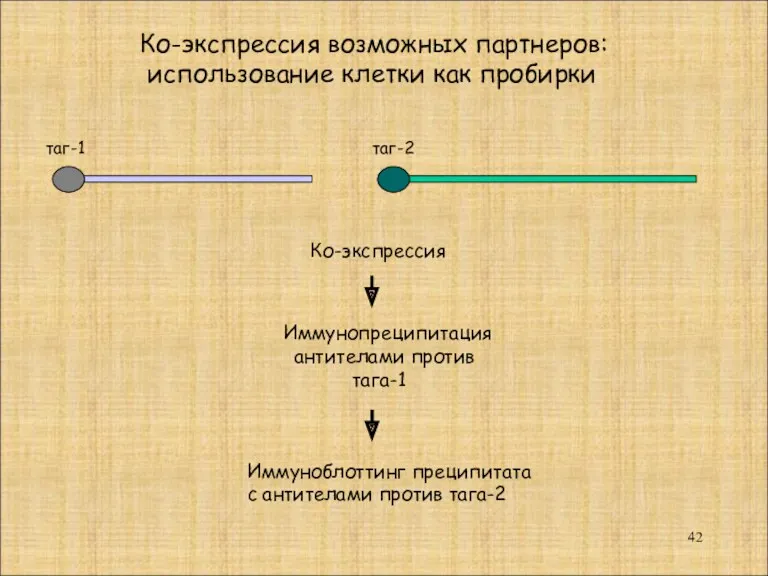
таг-1
таг-2
Ко-экспрессия
Иммунопреципитация
антителами против
тага-1
Иммуноблоттинг преципитата
с антителами против тага-2
Ко-экспрессия возможных партнеров:
таг-1
таг-2
Ко-экспрессия
Иммунопреципитация
антителами против
тага-1
Иммуноблоттинг преципитата
с антителами против тага-2
Ко-экспрессия возможных партнеров:
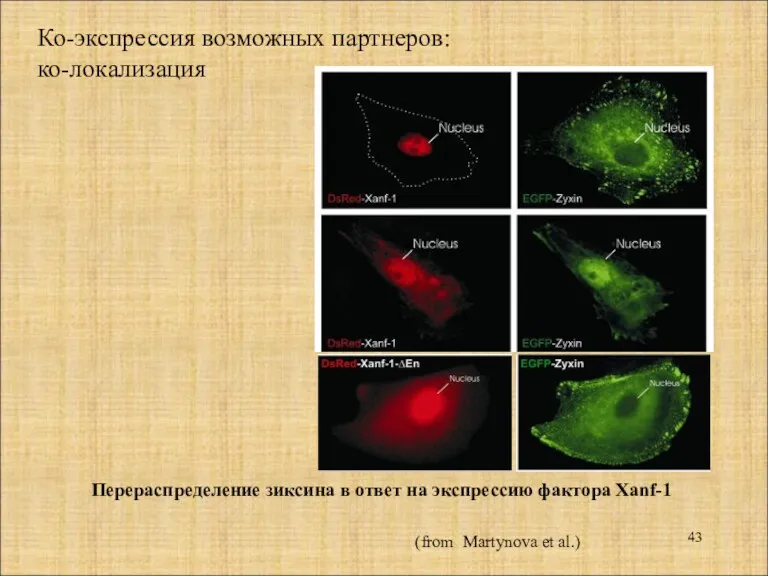
Ко-экспрессия возможных партнеров:
ко-локализация
Перераспределение зиксина в ответ на экспрессию фактора Xanf-1
(from
Ко-экспрессия возможных партнеров:
ко-локализация
Перераспределение зиксина в ответ на экспрессию фактора Xanf-1
(from
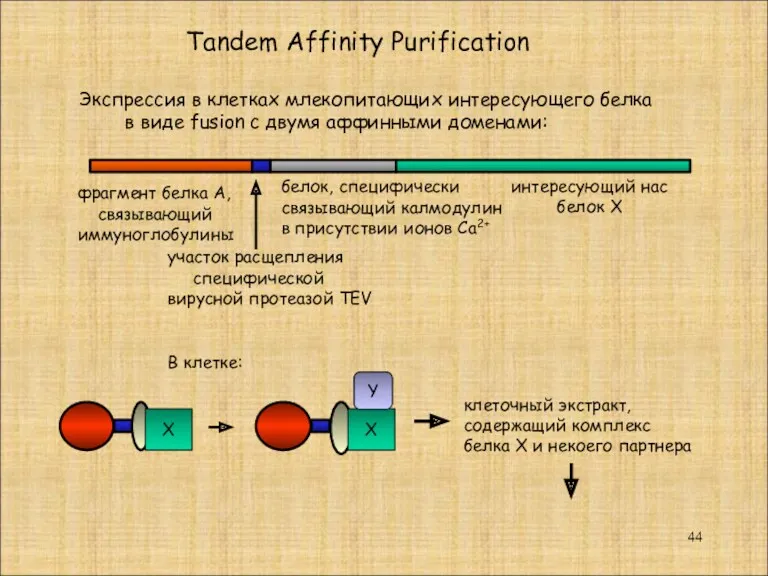
фрагмент белка А,
связывающий
иммуноглобулины
белок, специфически
связывающий калмодулин
в
фрагмент белка А,
связывающий
иммуноглобулины
белок, специфически
связывающий калмодулин
в
 Репликация. Эукариоты. Часть 2
Репликация. Эукариоты. Часть 2 Тип Хордовые. Бесчерепные. Ланцетник
Тип Хордовые. Бесчерепные. Ланцетник Механика и энергетика мышц
Механика и энергетика мышц Система опоры и движения. Опорно-двигательная система ОДС
Система опоры и движения. Опорно-двигательная система ОДС Обмен веществ и превращение энергии
Обмен веществ и превращение энергии Витамин B1
Витамин B1 Биотехнология: достижения и перспективы развития
Биотехнология: достижения и перспективы развития Обонятельный и вкусовой анализаторы
Обонятельный и вкусовой анализаторы Питание клетки.Фотосинтез.
Питание клетки.Фотосинтез. Соединительная ткань
Соединительная ткань Эволюция. Развитие представлений о возникновении жизни на Земле (часть 1)
Эволюция. Развитие представлений о возникновении жизни на Земле (часть 1) Естественный отбор – главная движущая сила эволюции
Естественный отбор – главная движущая сила эволюции Рисовая косметика
Рисовая косметика Краснокнижные виды растений и животных РТ
Краснокнижные виды растений и животных РТ Флокуляційне очищення питної води за допомогою катіонних та аніонних флокулянтів
Флокуляційне очищення питної води за допомогою катіонних та аніонних флокулянтів Транскрипция. Биосинтез белка
Транскрипция. Биосинтез белка Клетки и их разнообразие в многоклеточном организме
Клетки и их разнообразие в многоклеточном организме Охрана растений. Красная книга Архангельской области
Охрана растений. Красная книга Архангельской области Группы крови. Резус-фактор
Группы крови. Резус-фактор Липиды - сложные эфиры высших карбоновых кислот и ряда спиртов
Липиды - сложные эфиры высших карбоновых кислот и ряда спиртов Презентация Моллюски
Презентация Моллюски Пищевые цепи
Пищевые цепи Анатомия, физиология и методы исследования органа зрения
Анатомия, физиология и методы исследования органа зрения Проектно-исследовательская деятельность в условиях внедрения ФГОС ООО
Проектно-исследовательская деятельность в условиях внедрения ФГОС ООО Презентации к уроку по теме: Иммунитет
Презентации к уроку по теме: Иммунитет Отряды млекопитающих
Отряды млекопитающих Физиология прокариот
Физиология прокариот Вегетативное размножение растения
Вегетативное размножение растения