- Анатомия и возрастная физиология как учебная дисциплина

Содержание
- 2. Основная литература 1. Козлов В.И., Цехмистренко Т.А. Анатомия нервной системы. М.: Мир, 2006. 2. Безруких М.М.,
- 3. АНАТОМИЯ И ВОЗРАСТНАЯ ФИЗИОЛОГИЯ КАК УЧЕБНАЯ ДИСЦИПЛИНА
- 4. Анатомия и возрастная физиология изучает особенности строения и функционирования органов, систем органов и организма человека в
- 5. Анато́мия (от греч. ἀνα- — вновь, сверху и τέμνω — «режу», «рублю», «рассекаю») — раздел биологии
- 6. Анатомия и возрастная физиология является интегральной наукой о развитии организма человека, включающая в качестве составных частей
- 7. Основные задачи анатомии и возрастной физиологии • 1. Выяснение основных закономерностей роста и развития человека. •
- 8. Общая характеристика процессов роста и развития Общебиологическими свойствами живой материи являются процессы роста и развития, которые
- 9. Факторы, влияющие на рост и развитие ребенка Ребенок от взрослого человека отличается специфическими особенностями строения, биохимических
- 10. Факторы, влияющие на рост и развитие (резюме) Различают три группы основных факторов, определяющих развитие ребенка: Эндогенные
- 11. Акселерация и ретардация развития Акселерация развития (от лат. acceleratio — ускорение) — ускорение физического развития и
- 12. Установление параметров возрастной нормы Установление параметров возрастной нормы — одна из важнейших прикладных задач физиологии развития.
- 13. Основные положения теории онтогенеза В клетке с которой начинается онтогенез заложена генетическая программа дальнейшего развития всего
- 14. Вклад П. К. Анохина в развитие теории онтогенеза Академик П. К. Анохин создал теорию функциональных систем
- 15. Развитие идей П. К. Анохина успешно продолжается его учениками и последователями (С. Н. Хаютиным, К. В
- 16. Вклад Л. С. Выгодского в развитие теории онтогенеза Л. С. Выготский в своих лекциях по педологии
- 17. Возрастная периодизация — периодизация развития человека от рождения и до смерти, определение возрастных границ этапов в
- 18. Обоснование критериев периодизации индивидуального развития Индивидуальное развитие (онтогенез) - это целостный непрерывный процесс развития организма от
- 19. Возрастные периоды у детей Период новорождённости (неонатальный период) — первые 4 недели Грудной период: от 4-х
- 20. Возрастные периоды взрослого человека Юношеский период юноши: 17 — 21 год девушки: 16 — 20 лет
- 21. Критические периоды развития С проблемой возрастной периодизации непосредственно связан вопрос о критических и сенситивных периодах развития.
- 22. Выделяют несколько критических периодов развития Наиболее опасными периодами считаются: 1) время развития половых клеток - овогенез
- 23. Критические периоды пренатального развития ребенка
- 24. Талидомидная трагедия В 1954 году немецкая фармацевтическая компания Chemie Grünenthal разработала препарат талидомид. Тестирование показало, что
- 25. Сенситивные периоды развития Сенситивный период развития (встречается также сензитивный) — период в жизни человека, создающий наиболее
- 26. Основные методологические подходы в биологии развития - это 1) “утрата функции” (“loss of function”) 2) “приобретение
- 27. Карты презумптивных зачатков зародышей позвоночных (создаются на основе топографии ранней гаструлы)
- 28. Раннее развитие зародышей животных может быть детерминированным или регулятивным
- 29. Карта презумптивных зачатков (судьбы) зародыша оболочника, может быть построена уже на одноклеточном зародыше (детерминированное развитие)
- 30. Автономная спецификация (детерминированное развитие) раннего зародыша асцидии: из различных бластомеров при культивировании образуются различные структуры
- 31. Карта судьбы зародыша оболочника (продолжение)
- 32. Автономная спецификация (мозаическое развитие) клеток зародыша моллюска Patella
- 33. Регулятивное развитие Может приводить к появлению однояйцевых близнецов У броненосца Daspus novemcintus первоначальный зародыш разделяется на
- 34. Регулятивное развитие морского ежа
- 35. Регулятивное развитие Изучение регулятивного развития со всей очевидностью позволило установить существование двух важных для развития явлений:
- 36. Зависимая спецификация (регулятивное развитие) у земноводных. Недостающие клетки восполняются без потери функций поврежденного участка. Судьба пересаженных
- 37. Региональная специфичность индукции структур эпидермального эпителия под воздействием различных участков дермиса у куриного зародыша
- 38. Пересадка клеток имеющих морфологические различия, но сходных по судьбе, у зародышей близкородственных видов позволяет проследить судьбу
- 39. Участие транскрипционного фактора Pax6 в индукции зрительных и назальных структур у зародыша крысы (Слева -норма, справа
- 40. Паттернинг - процесс, в котором из эмбриональных клеток устанавливается пространственно упорядоченная организация каких-либо тканей ( паттерн).
- 41. Экспрессия гена sonic hedgehog в различных зачатках 3-х дневного куриного зародыша (иРНК)
- 42. Экспрессия гена Vg1 в вегетативной области ооцита Xenopus laevis (иРНК)
- 43. Паттернирование Создание (установление) паттерна (динамических пространственных структур) может объясняться достаточно простыми причинами (например градиентами концентраций каких-либо
- 44. Реакционно-диффузионная модель Тьюринга для объяснения формирования пространственной морфологической структуры (паттерна). Создание волн (периодических значений) концентраций веществ,
- 45. Компьютерное моделирование паттерна пигментации чешуи рыбки Danio rerio (с учётом реакционно-диффузионной модели) и его соответствие паттернам
- 46. Одни клетки (развивающегося) многоклеточного организма могут влиять на судьбу других (соседних) клеток, секретируя во внеклеточное пространство
- 47. Возможности регенерации плоского червя (регенерация может быть обязана морфогенам, действующим вдоль продольной оси )
- 48. Варианты дифференцировки in vitro клеток анимальной шапочки бластулы шпорцевой лягушки Xenopus laevis под влиянием различных концентраций
- 49. Градиент активина при разных концентрациях индуцирует in vitro экспрессию различных генов в клетках анимальной шапочки Xenopus
- 50. Интерпретация градиента концентрации активина в клетках анимальной шапочки Xenopus laevis (в модельных экспериментах)
- 51. Морфогенетическое поле зачатка конечности саламандры Ambistoma maculatum
- 52. Морфогенетическое поле (МП) может быть определено как группа клеток , чьё расположение и назначение задано в
- 53. Роль позиционной информации в морфогенезе на примере модели “французского (трёхцветного) флага” (L. Wolpert). Интерпретация аналоговой (позиционной)
- 54. Если презумптивные клетки из МП ноги Drosophila перенести в МП антенны (апикальной части предполагаемой антенны) ,
- 55. Развитие многоклеточных организмов происходит в соответствии с двумя парадигмами: Дифференциальная активность генов Избирательные межклеточные взаимодействия
- 56. Дифференциальная экспрессия генов Экспрессия гена - реализация генетической информации, закодированной в гене. Экспрессия происходит последовательно на
- 57. ДИФФЕРЕНЦИАЛЬНАЯ ЭКСПРЕССИЯ ГЕНОВ В основе дифференциальной активности генов на уровне транскрипции лежат обратимые (эпигеномные) изменения отдельных
- 58. Онтогенетическая стабильность генома (эквивалентность клеточных геномов в онтогенезе) - фундаментальная характеристика половых и соматических клеток большинства
- 59. Цитологические, генетические, эмбриологические и молекулярно-биологические свидетельства стабильности (эквивалентности) генома в клетках различных типов тканей и органов.
- 60. Явления онтогенетической нестабильности генома в соматических клетках ряда животных как результат геномных перестроек соматических клеток. Диминуция
- 61. Политенные хромосомы слюнных желёз личинки Drosophila melanogaster (дрозофилы)
- 62. Участие транскрипционного фактора Pax6 в индукции оптических и назальных структур у зародыша крысы (Слева -норма, справа
- 63. Множественные 5’регуляторные участки (энхансеры) гена Pax6 мыши (позволяют гену экспрессироваться в разных тканях и органах)
- 64. Сайленсер NRSE ограничивает районы экспрессии гена L1 в зародыше мыши, позволяя ему экспрессироваться только в клетках
- 65. LCR и кластерные энхансеры. Организация генов семейств альфа и бета глобинов в соответствующие кластеры: смена эмбриональных
- 66. Изменение типов полипетидных цепей глобинов в молекуле гемоглобина в зависимости от локализации эритропоэза в пре- и
- 67. Образование семейства альфа-тропомиозинов у крысы за счёт альтернативного сплайсинга транскрипта одного гена (слева в столбце указаны
- 68. Детерминация пола у дрозофилы. Альтернативный сплайсинг пре-иРНК гена sex-lethal приводит к альтернативному сплайсингу пре-иРНК последовательно генов
- 69. Регуляция экспрессии гена “материнского эффекта” на уровне трансляции в ооците мыши. Инактивация иРНК путём редукции 3'-poly(A)
- 70. Запасение различных иРНК в ооците и их трансляция после оплодотворения (гены “материнского эффекта”)
- 71. Поддержание концентрации внутриклеточного Fe координированной регуляцией трансляции иРНК ферритина и рецептора трансферрина с использованием специфического сайта
- 72. Регуляция трансляции РНК lin 14 нематоды C. elegans c помощью микроРНК lin 4
- 73. Образование и функционирование микроРНК у нематоды
- 74. Регуляция прохождения линек C. elegans микроРНК Figure 1 Model for the activity of heterochronic genes in
- 75. Регуляция экспрессии генов метилированием участков промотора
- 76. Инсуляторы, предполагаемый механизм действия
- 77. Геномный импринтинг у млекопитающих - различия в степени метилирования отцовских и материнских аллелей гена (5-метил-цитозин) может
- 78. Больные с синдромами: Angelman (А-слева ) и Prader-Willi (P-W - справа). A- серьёзная умственная отсталость, припадки,
- 79. Дозовая компенсация генов половых хромосом млекопитающих Инактивация одной из Х-хромосом млекопитающих. Конденсирование хроматина в период раннего
- 80. Гипотеза М. Лайон об инактивации Х-хромосомы (M. Lyon, 1961) 1. В очень раннем развитии самок млекопитающих
- 81. Вероятностный характер инактивации X-хромосом в различных клетках млекопитающих. Пример- животные с рыже-белой (калико) и черепаховой окраской.
- 82. Пояснения к предыдущему рисунку Хотя термины «калико» и «черепаховый» рассматривают иногда как синонимичные, на шкуре кошек
- 83. Механизм инактивация Х-хромосомы у млекопитающих: роль РНК - продукта гена Xist в инициации инактивации. Выбор в
- 84. Процессы, обусловливающие инактивацию X-хромосомы (млекопитающие)
- 86. Число генов в геномах многоклеточных организмов хотя и увеличивается в эволюционом ряду от низших к высшим,
- 87. Экспериментально индуцированная экспрессия гена Pax6 в области имагинального диска антенны дрозофилы
- 88. Генные сети участвуют в реализации генетических программ дифференцировки клеток
- 90. Скачать презентацию

Основная литература
1. Козлов В.И., Цехмистренко Т.А. Анатомия нервной системы. М.: Мир,
Основная литература
1. Козлов В.И., Цехмистренко Т.А. Анатомия нервной системы. М.: Мир,

АНАТОМИЯ И ВОЗРАСТНАЯ ФИЗИОЛОГИЯ КАК УЧЕБНАЯ ДИСЦИПЛИНА
АНАТОМИЯ И ВОЗРАСТНАЯ ФИЗИОЛОГИЯ КАК УЧЕБНАЯ ДИСЦИПЛИНА
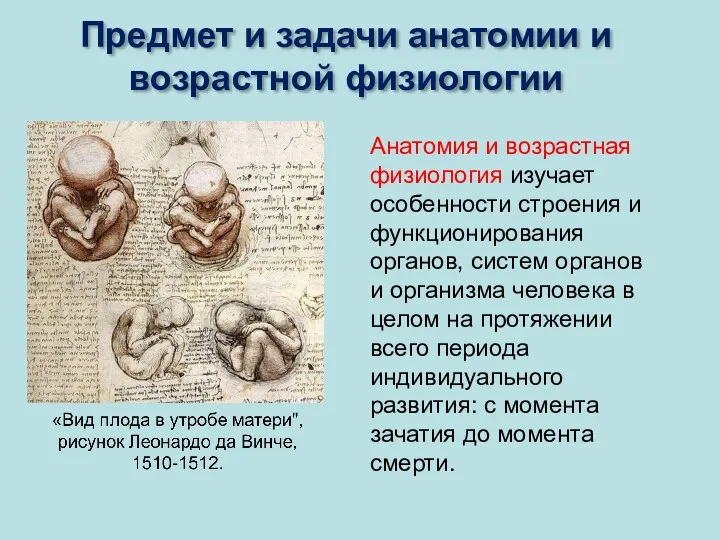
Анатомия и возрастная физиология изучает особенности строения и функционирования органов, систем
Анатомия и возрастная физиология изучает особенности строения и функционирования органов, систем

Анато́мия (от греч. ἀνα- — вновь, сверху и τέμνω — «режу»,
Анато́мия (от греч. ἀνα- — вновь, сверху и τέμνω — «режу»,

Анатомия и возрастная физиология является интегральной наукой о развитии организма человека,
Анатомия и возрастная физиология является интегральной наукой о развитии организма человека,

Основные задачи анатомии и возрастной физиологии
• 1. Выяснение основных закономерностей роста
Основные задачи анатомии и возрастной физиологии
• 1. Выяснение основных закономерностей роста

Общая характеристика процессов роста и развития
Общебиологическими свойствами живой материи являются процессы
Общая характеристика процессов роста и развития
Общебиологическими свойствами живой материи являются процессы

Факторы, влияющие на рост и развитие ребенка
Ребенок от взрослого человека отличается
Факторы, влияющие на рост и развитие ребенка
Ребенок от взрослого человека отличается

Факторы, влияющие на рост и развитие (резюме)
Различают три группы основных
Факторы, влияющие на рост и развитие (резюме)
Различают три группы основных

Акселерация и ретардация развития
Акселерация развития (от лат. acceleratio — ускорение) —
Акселерация и ретардация развития
Акселерация развития (от лат. acceleratio — ускорение) —

Установление параметров возрастной нормы
Установление параметров возрастной нормы — одна из важнейших
Установление параметров возрастной нормы
Установление параметров возрастной нормы — одна из важнейших

Основные положения теории онтогенеза
В клетке с которой начинается онтогенез заложена генетическая
Основные положения теории онтогенеза
В клетке с которой начинается онтогенез заложена генетическая

Вклад П. К. Анохина в развитие теории онтогенеза
Академик П. К. Анохин
Вклад П. К. Анохина в развитие теории онтогенеза
Академик П. К. Анохин

Развитие идей П. К. Анохина успешно продолжается его учениками и последователями
Развитие идей П. К. Анохина успешно продолжается его учениками и последователями

Вклад Л. С. Выгодского в развитие теории онтогенеза
Л. С. Выготский в
Вклад Л. С. Выгодского в развитие теории онтогенеза
Л. С. Выготский в

Возрастная периодизация — периодизация развития человека от рождения и до смерти,
Возрастная периодизация — периодизация развития человека от рождения и до смерти,

Обоснование критериев периодизации индивидуального развития
Индивидуальное развитие (онтогенез) - это целостный непрерывный
Обоснование критериев периодизации индивидуального развития
Индивидуальное развитие (онтогенез) - это целостный непрерывный

Возрастные периоды у детей
Период новорождённости (неонатальный период) — первые 4 недели
Грудной
Возрастные периоды у детей
Период новорождённости (неонатальный период) — первые 4 недели
Грудной

Возрастные периоды взрослого человека
Юношеский период
юноши: 17 — 21 год
девушки: 16
Возрастные периоды взрослого человека
Юношеский период
юноши: 17 — 21 год
девушки: 16

Критические периоды развития
С проблемой возрастной периодизации непосредственно связан вопрос о критических
Критические периоды развития
С проблемой возрастной периодизации непосредственно связан вопрос о критических

Выделяют несколько критических периодов развития
Наиболее опасными периодами считаются:
1) время развития половых
Выделяют несколько критических периодов развития
Наиболее опасными периодами считаются:
1) время развития половых

Критические периоды пренатального
развития ребенка
Критические периоды пренатального
развития ребенка

Талидомидная трагедия
В 1954 году немецкая фармацевтическая компания Chemie Grünenthal разработала препарат
Талидомидная трагедия
В 1954 году немецкая фармацевтическая компания Chemie Grünenthal разработала препарат

Сенситивные периоды развития
Сенситивный период развития (встречается также сензитивный) — период в
Сенситивные периоды развития
Сенситивный период развития (встречается также сензитивный) — период в

Основные методологические подходы в биологии развития - это
1) “утрата функции” (“loss
Основные методологические подходы в биологии развития - это 1) “утрата функции” (“loss

Карты презумптивных зачатков зародышей позвоночных (создаются на основе топографии ранней гаструлы)
Карты презумптивных зачатков зародышей позвоночных (создаются на основе топографии ранней гаструлы)

Раннее развитие зародышей животных может быть детерминированным
или регулятивным
Раннее развитие зародышей животных может быть детерминированным
или регулятивным

Карта презумптивных зачатков (судьбы) зародыша оболочника, может быть построена уже на
Карта презумптивных зачатков (судьбы) зародыша оболочника, может быть построена уже на

Автономная спецификация (детерминированное развитие) раннего зародыша асцидии: из различных бластомеров при
Автономная спецификация (детерминированное развитие) раннего зародыша асцидии: из различных бластомеров при
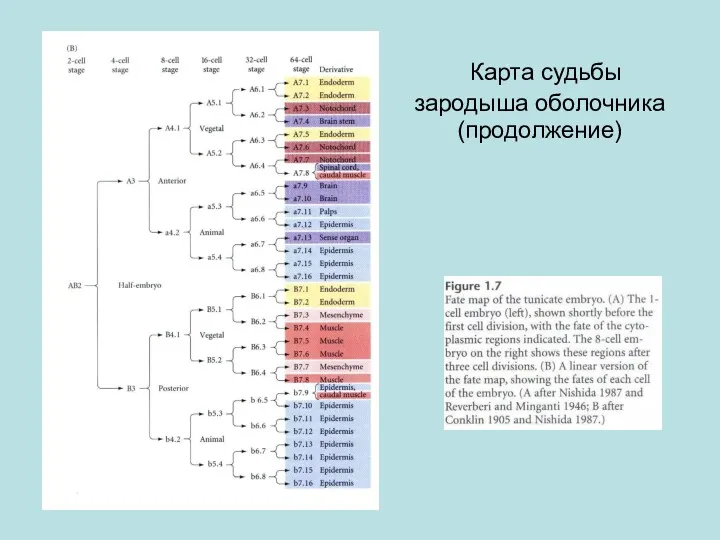
Карта судьбы зародыша оболочника (продолжение)
Карта судьбы зародыша оболочника (продолжение)

Автономная спецификация (мозаическое развитие) клеток зародыша моллюска Patella
Автономная спецификация (мозаическое развитие) клеток зародыша моллюска Patella
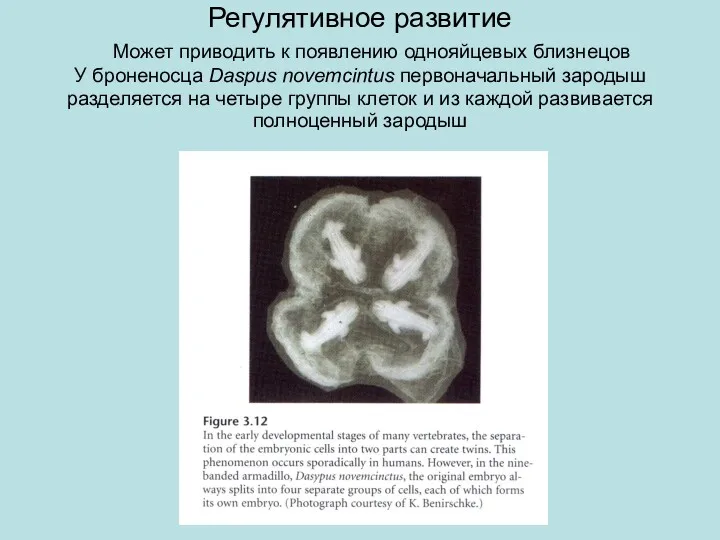
Регулятивное развитие Может приводить к появлению однояйцевых близнецов
У броненосца Daspus novemcintus
Регулятивное развитие Может приводить к появлению однояйцевых близнецов У броненосца Daspus novemcintus

Регулятивное развитие морского ежа
Регулятивное развитие морского ежа

Регулятивное развитие
Изучение регулятивного развития со всей очевидностью позволило установить существование двух
Регулятивное развитие
Изучение регулятивного развития со всей очевидностью позволило установить существование двух
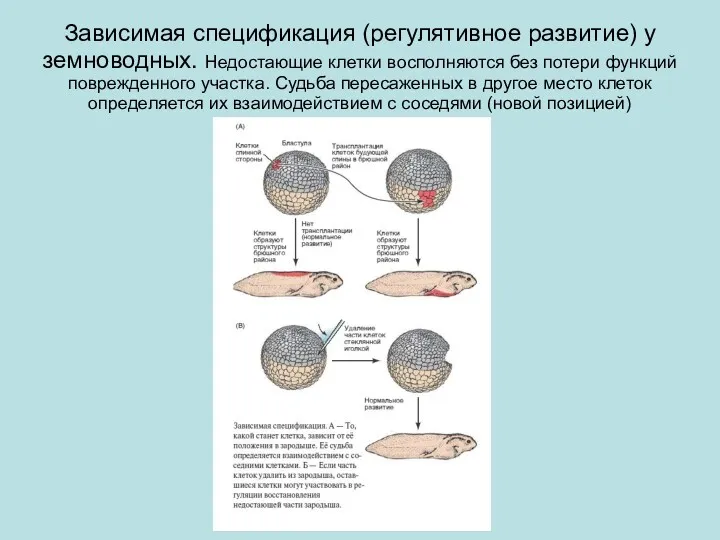
Зависимая спецификация (регулятивное развитие) у земноводных. Недостающие клетки восполняются без потери
Зависимая спецификация (регулятивное развитие) у земноводных. Недостающие клетки восполняются без потери

Региональная специфичность индукции структур эпидермального эпителия под воздействием различных участков дермиса
Региональная специфичность индукции структур эпидермального эпителия под воздействием различных участков дермиса
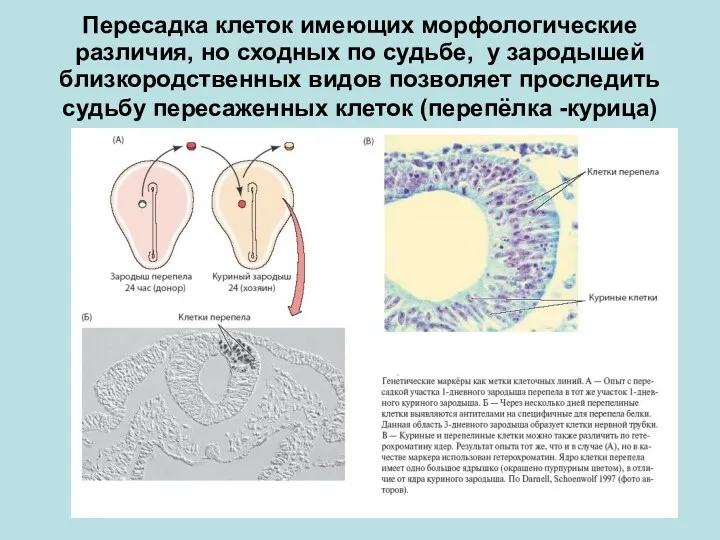
Пересадка клеток имеющих морфологические различия, но сходных по судьбе, у зародышей
Пересадка клеток имеющих морфологические различия, но сходных по судьбе, у зародышей
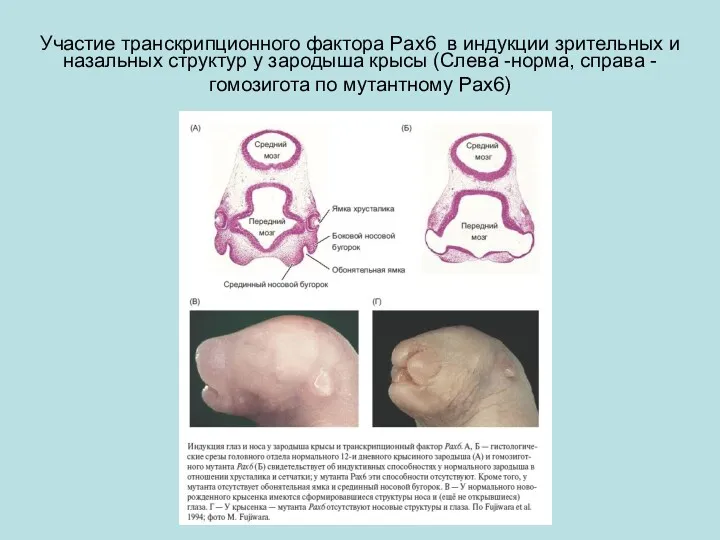
Участие транскрипционного фактора Pax6 в индукции зрительных и назальных структур у
Участие транскрипционного фактора Pax6 в индукции зрительных и назальных структур у

Паттернинг - процесс, в котором из эмбриональных клеток устанавливается пространственно упорядоченная
Паттернинг - процесс, в котором из эмбриональных клеток устанавливается пространственно упорядоченная
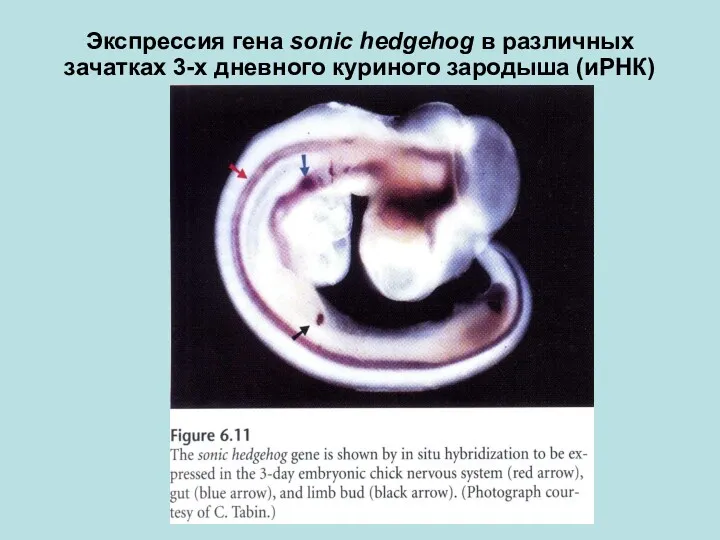
Экспрессия гена sonic hedgehog в различных зачатках 3-х дневного куриного зародыша
Экспрессия гена sonic hedgehog в различных зачатках 3-х дневного куриного зародыша
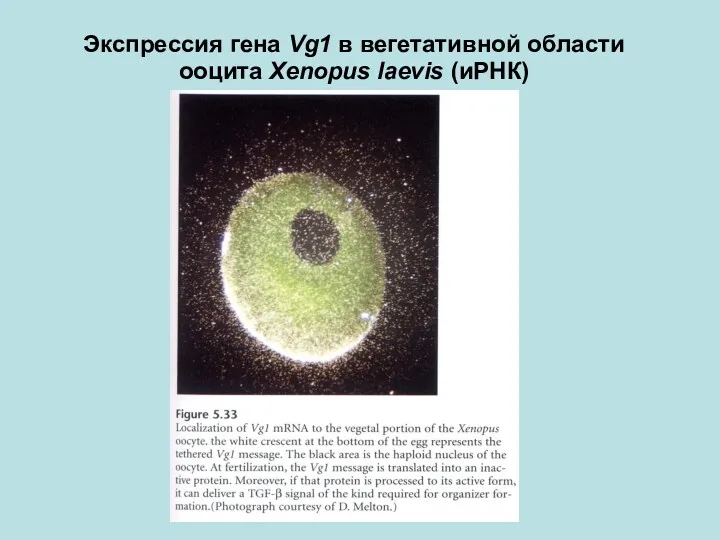
Экспрессия гена Vg1 в вегетативной области ооцита Xenopus laevis (иРНК)
Экспрессия гена Vg1 в вегетативной области ооцита Xenopus laevis (иРНК)

Паттернирование
Создание (установление) паттерна (динамических пространственных структур) может объясняться достаточно простыми причинами
Паттернирование
Создание (установление) паттерна (динамических пространственных структур) может объясняться достаточно простыми причинами

Реакционно-диффузионная модель Тьюринга для объяснения формирования пространственной морфологической структуры (паттерна). Создание
Реакционно-диффузионная модель Тьюринга для объяснения формирования пространственной морфологической структуры (паттерна). Создание

Компьютерное моделирование паттерна пигментации чешуи рыбки Danio rerio (с учётом
Компьютерное моделирование паттерна пигментации чешуи рыбки Danio rerio (с учётом

Одни клетки (развивающегося) многоклеточного организма могут влиять на судьбу других (соседних)
Одни клетки (развивающегося) многоклеточного организма могут влиять на судьбу других (соседних)
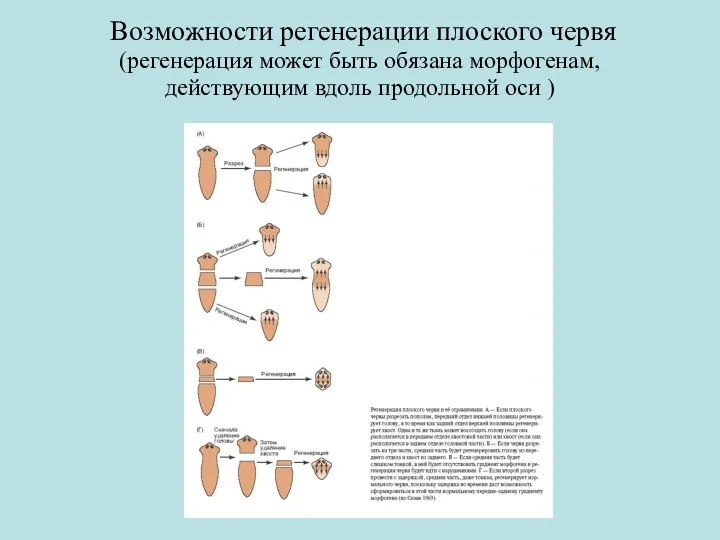
Возможности регенерации плоского червя
(регенерация может быть обязана морфогенам, действующим вдоль
Возможности регенерации плоского червя
(регенерация может быть обязана морфогенам, действующим вдоль
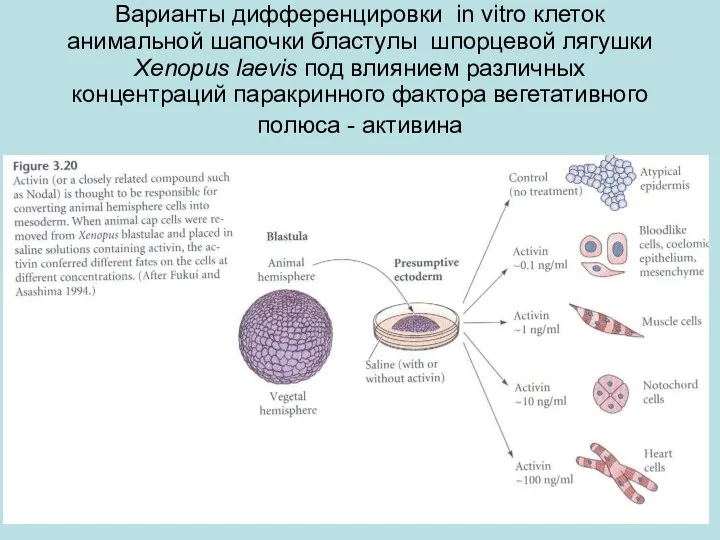
Варианты дифференцировки in vitro клеток анимальной шапочки бластулы шпорцевой лягушки Xenopus
Варианты дифференцировки in vitro клеток анимальной шапочки бластулы шпорцевой лягушки Xenopus
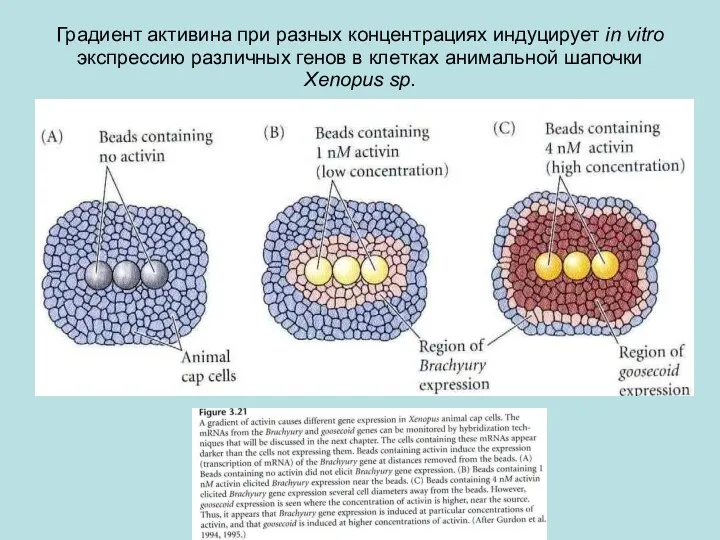
Градиент активина при разных концентрациях индуцирует in vitro экспрессию различных генов
Градиент активина при разных концентрациях индуцирует in vitro экспрессию различных генов

Интерпретация градиента концентрации активина в клетках анимальной шапочки Xenopus laevis (в
Интерпретация градиента концентрации активина в клетках анимальной шапочки Xenopus laevis (в
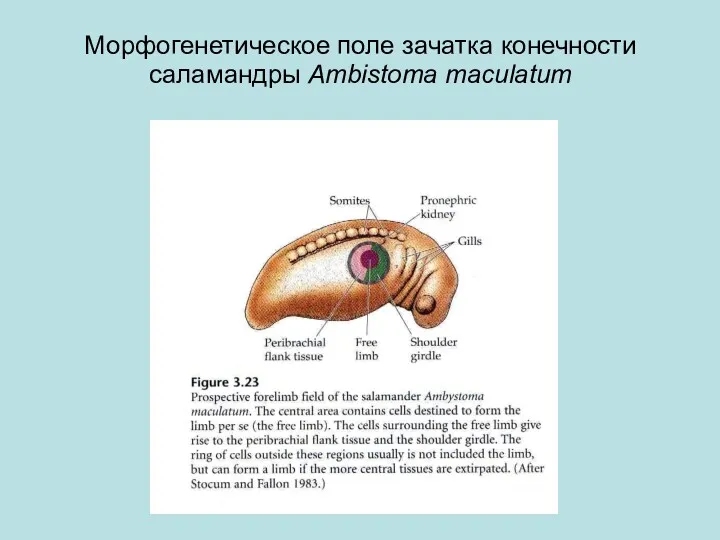
Морфогенетическое поле зачатка конечности саламандры Ambistoma maculatum
Морфогенетическое поле зачатка конечности саламандры Ambistoma maculatum

Морфогенетическое поле (МП) может быть определено как группа клеток , чьё
Морфогенетическое поле (МП) может быть определено как группа клеток , чьё
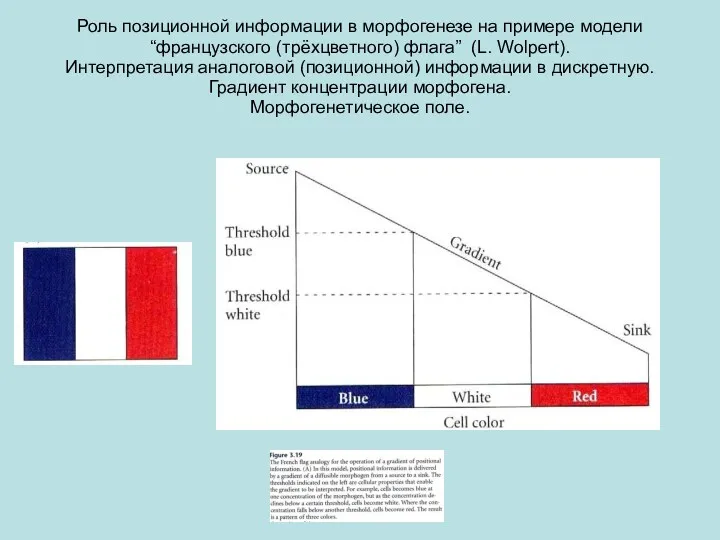
Роль позиционной информации в морфогенезе на примере модели “французского (трёхцветного) флага”
Роль позиционной информации в морфогенезе на примере модели “французского (трёхцветного) флага”
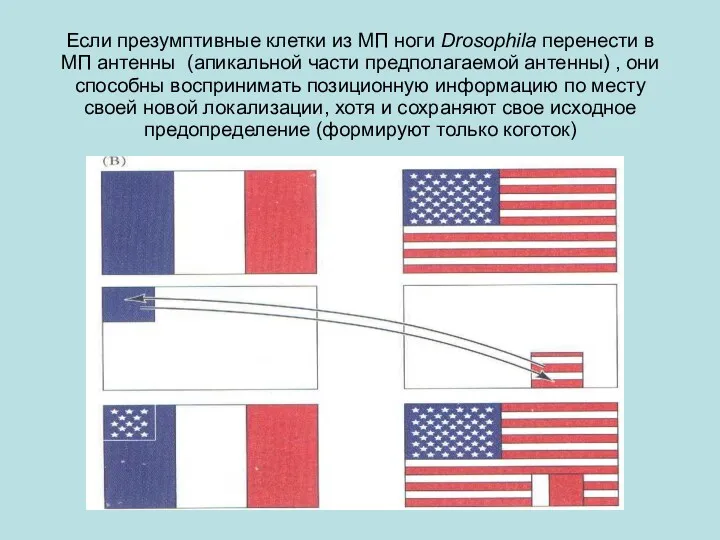
Если презумптивные клетки из МП ноги Drosophila перенести в МП антенны
Если презумптивные клетки из МП ноги Drosophila перенести в МП антенны

Развитие многоклеточных организмов
происходит в соответствии с двумя парадигмами:
Дифференциальная активность генов
Избирательные межклеточные
Развитие многоклеточных организмов
происходит в соответствии с двумя парадигмами:
Дифференциальная активность генов
Избирательные межклеточные

Дифференциальная экспрессия генов
Экспрессия гена - реализация генетической информации, закодированной в гене.
Дифференциальная экспрессия генов Экспрессия гена - реализация генетической информации, закодированной в гене.

ДИФФЕРЕНЦИАЛЬНАЯ ЭКСПРЕССИЯ ГЕНОВ
В основе дифференциальной активности генов на уровне транскрипции
ДИФФЕРЕНЦИАЛЬНАЯ ЭКСПРЕССИЯ ГЕНОВ В основе дифференциальной активности генов на уровне транскрипции

Онтогенетическая стабильность генома (эквивалентность клеточных геномов в онтогенезе) - фундаментальная характеристика
Онтогенетическая стабильность генома (эквивалентность клеточных геномов в онтогенезе) - фундаментальная характеристика

Цитологические, генетические, эмбриологические и молекулярно-биологические свидетельства
стабильности (эквивалентности) генома в клетках
Цитологические, генетические, эмбриологические и молекулярно-биологические свидетельства стабильности (эквивалентности) генома в клетках

Явления онтогенетической нестабильности генома в соматических клетках ряда животных как
Явления онтогенетической нестабильности генома в соматических клетках ряда животных как
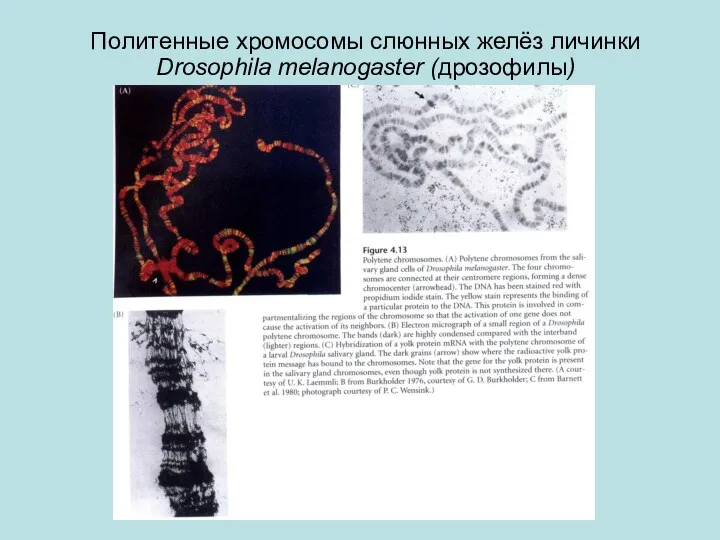
Политенные хромосомы слюнных желёз личинки Drosophila melanogaster (дрозофилы)
Политенные хромосомы слюнных желёз личинки Drosophila melanogaster (дрозофилы)
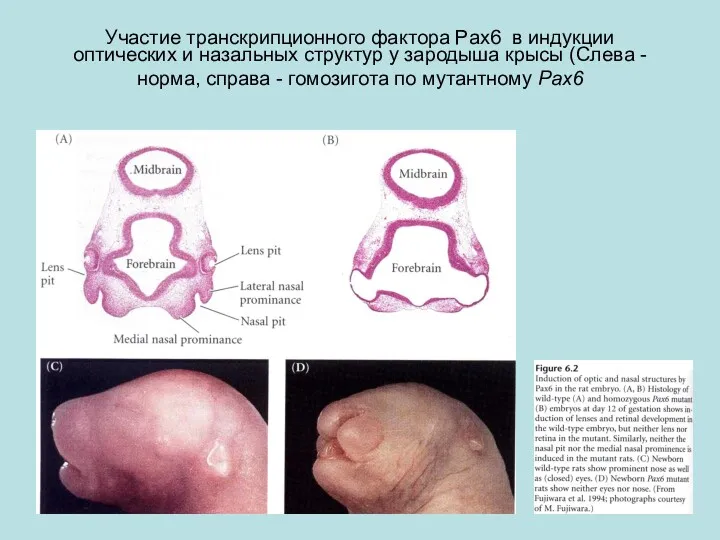
Участие транскрипционного фактора Pax6 в индукции оптических и назальных структур у
Участие транскрипционного фактора Pax6 в индукции оптических и назальных структур у

Множественные 5’регуляторные участки (энхансеры) гена Pax6 мыши (позволяют гену экспрессироваться в
Множественные 5’регуляторные участки (энхансеры) гена Pax6 мыши (позволяют гену экспрессироваться в
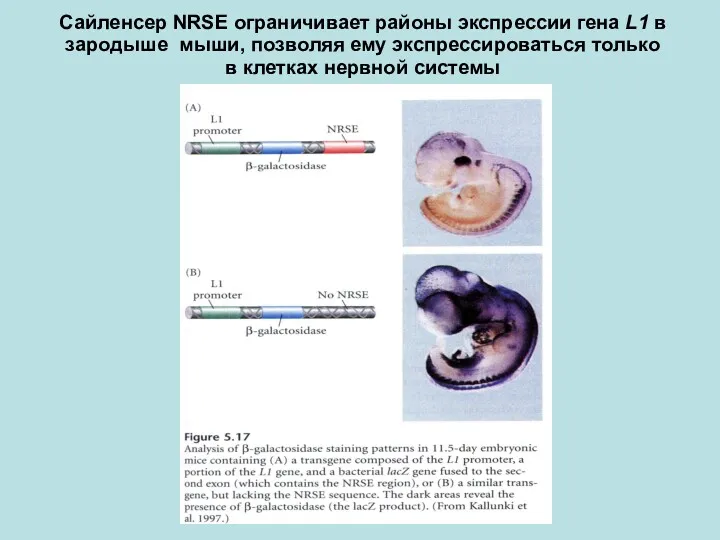
Сайленсер NRSE ограничивает районы экспрессии гена L1 в зародыше мыши, позволяя
Сайленсер NRSE ограничивает районы экспрессии гена L1 в зародыше мыши, позволяя
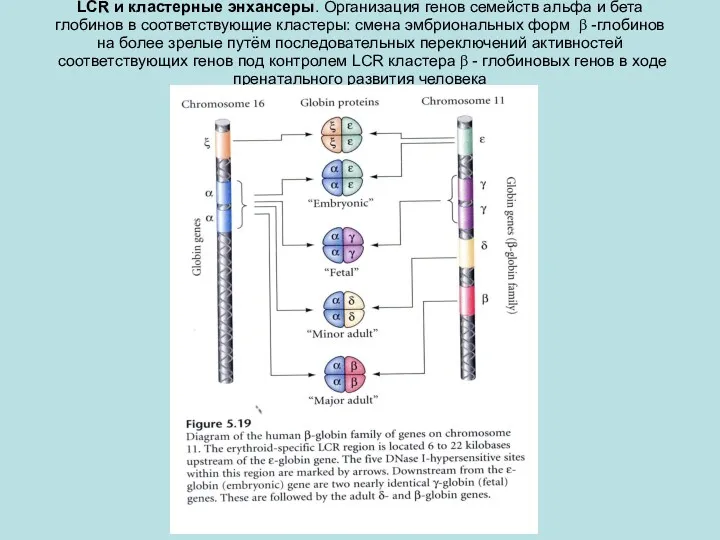
LCR и кластерные энхансеры. Организация генов семейств альфа и бета глобинов
LCR и кластерные энхансеры. Организация генов семейств альфа и бета глобинов
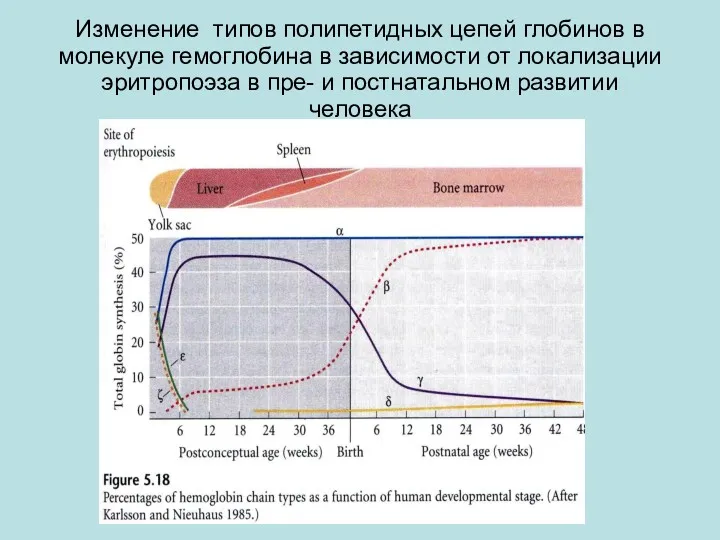
Изменение типов полипетидных цепей глобинов в молекуле гемоглобина в зависимости от
Изменение типов полипетидных цепей глобинов в молекуле гемоглобина в зависимости от

Образование семейства альфа-тропомиозинов у крысы за счёт альтернативного сплайсинга транскрипта одного
Образование семейства альфа-тропомиозинов у крысы за счёт альтернативного сплайсинга транскрипта одного

Детерминация пола у дрозофилы. Альтернативный сплайсинг пре-иРНК гена sex-lethal приводит к
Детерминация пола у дрозофилы. Альтернативный сплайсинг пре-иРНК гена sex-lethal приводит к
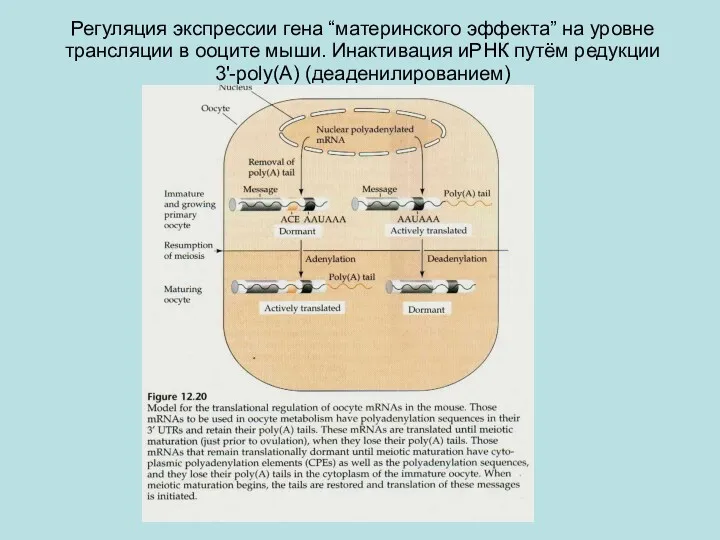
Регуляция экспрессии гена “материнского эффекта” на уровне трансляции в ооците мыши.
Регуляция экспрессии гена “материнского эффекта” на уровне трансляции в ооците мыши.

Запасение различных иРНК в ооците и их трансляция после оплодотворения (гены
Запасение различных иРНК в ооците и их трансляция после оплодотворения (гены
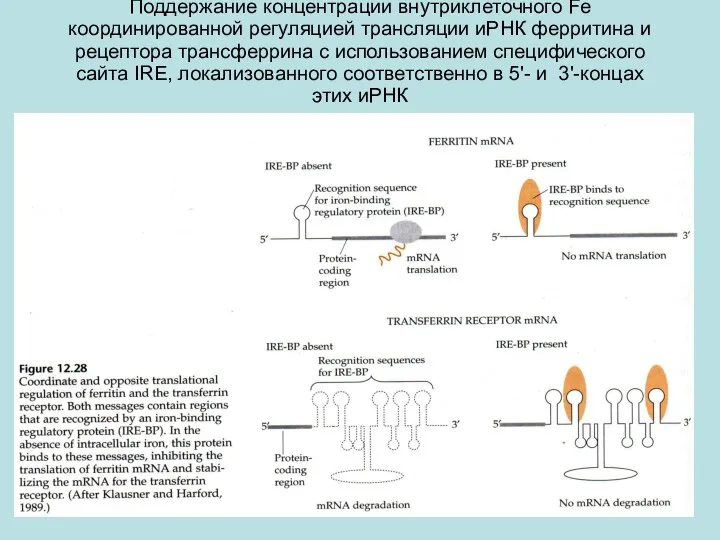
Поддержание концентрации внутриклеточного Fe координированной регуляцией трансляции иРНК ферритина и рецептора
Поддержание концентрации внутриклеточного Fe координированной регуляцией трансляции иРНК ферритина и рецептора

Регуляция трансляции РНК lin 14 нематоды C. elegans c помощью микроРНК
Регуляция трансляции РНК lin 14 нематоды C. elegans c помощью микроРНК

Образование и функционирование микроРНК у нематоды
Образование и функционирование микроРНК у нематоды
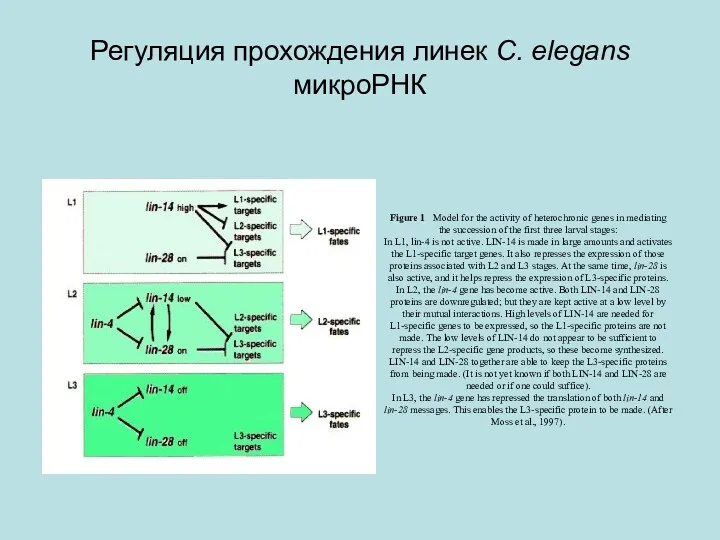
Регуляция прохождения линек C. elegans микроРНК
Figure 1 Model for the activity
Регуляция прохождения линек C. elegans микроРНК
Figure 1 Model for the activity

Регуляция экспрессии генов метилированием участков промотора
Регуляция экспрессии генов метилированием участков промотора

Инсуляторы, предполагаемый механизм действия
Инсуляторы, предполагаемый механизм действия
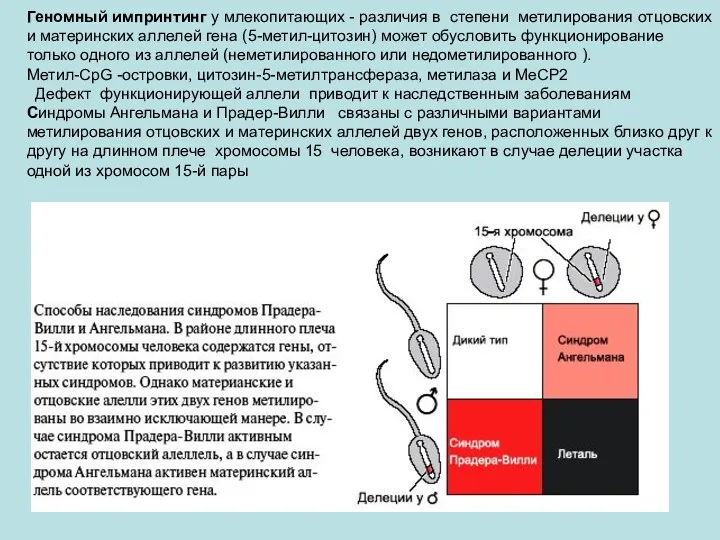
Геномный импринтинг у млекопитающих - различия в степени метилирования отцовских и
Геномный импринтинг у млекопитающих - различия в степени метилирования отцовских и
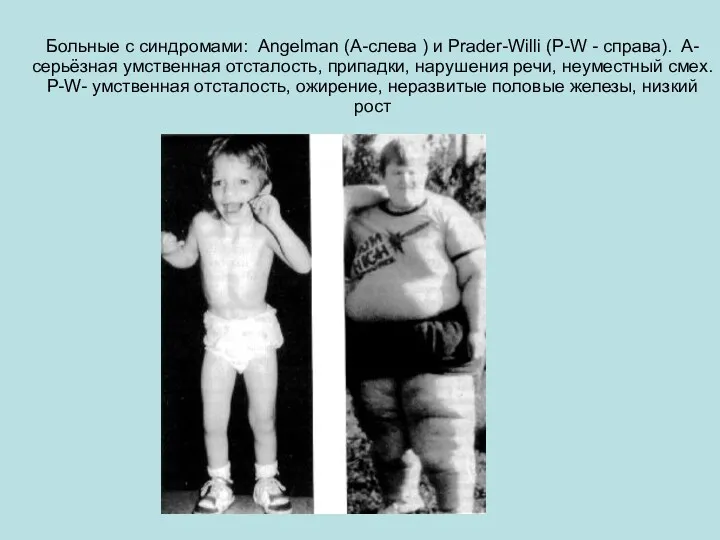
Больные с синдромами: Angelman (А-слева ) и Prader-Willi (P-W - справа).
Больные с синдромами: Angelman (А-слева ) и Prader-Willi (P-W - справа).

Дозовая компенсация генов половых хромосом млекопитающих
Инактивация одной из Х-хромосом млекопитающих. Конденсирование
Дозовая компенсация генов половых хромосом млекопитающих Инактивация одной из Х-хромосом млекопитающих. Конденсирование

Гипотеза М. Лайон об инактивации Х-хромосомы (M. Lyon, 1961)
1. В очень
Гипотеза М. Лайон об инактивации Х-хромосомы (M. Lyon, 1961)
1. В очень

Вероятностный характер инактивации X-хромосом в различных клетках млекопитающих. Пример- животные с
Вероятностный характер инактивации X-хромосом в различных клетках млекопитающих. Пример- животные с

Пояснения к предыдущему рисунку
Хотя термины «калико» и «черепаховый» рассматривают иногда как
Пояснения к предыдущему рисунку
Хотя термины «калико» и «черепаховый» рассматривают иногда как
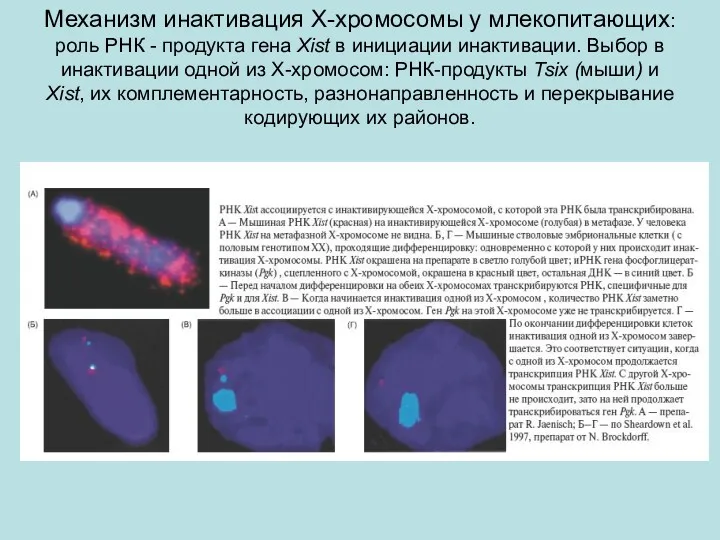
Механизм инактивация Х-хромосомы у млекопитающих: роль РНК - продукта гена Xist
Механизм инактивация Х-хромосомы у млекопитающих: роль РНК - продукта гена Xist
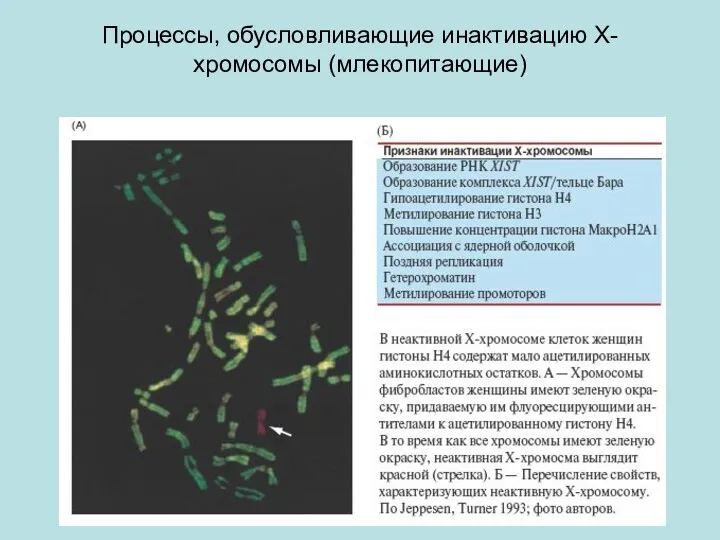
Процессы, обусловливающие инактивацию X-хромосомы (млекопитающие)
Процессы, обусловливающие инактивацию X-хромосомы (млекопитающие)


Число генов в геномах многоклеточных организмов хотя и увеличивается в
Число генов в геномах многоклеточных организмов хотя и увеличивается в

Экспериментально индуцированная экспрессия гена Pax6 в области имагинального диска антенны дрозофилы
Экспериментально индуцированная экспрессия гена Pax6 в области имагинального диска антенны дрозофилы
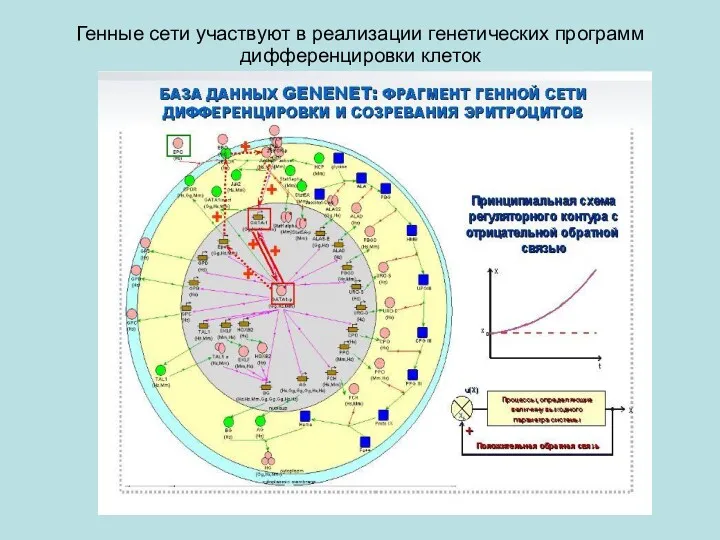
Генные сети участвуют в реализации генетических программ дифференцировки клеток
Генные сети участвуют в реализации генетических программ дифференцировки клеток
 Вегетативті көбею
Вегетативті көбею Девонский период палеозойской эры
Девонский период палеозойской эры Многообразие цветковых растений. Классы покрытосеменных. Семейства. Роль растений в природе и жизни человека
Многообразие цветковых растений. Классы покрытосеменных. Семейства. Роль растений в природе и жизни человека Обмен веществ и энергией
Обмен веществ и энергией Органы цветковых растений
Органы цветковых растений Витамины. Классификация витаминов
Витамины. Классификация витаминов Особенности желудочного пищеварения у жвачных
Особенности желудочного пищеварения у жвачных Тип кишечнополостные (пресноводная гидра)
Тип кишечнополостные (пресноводная гидра) Экспертиза качества молока
Экспертиза качества молока Утворення торфа та кам’яного вугілля
Утворення торфа та кам’яного вугілля Пестрая ситовая (ямчато-волокнистая) гниль корней. Корневая губка
Пестрая ситовая (ямчато-волокнистая) гниль корней. Корневая губка Функциональная структура биосферы
Функциональная структура биосферы Тұқым қуалаушылықтың молекулярлық негіздері
Тұқым қуалаушылықтың молекулярлық негіздері Рефлекс. Рефлекторная дуга. Спинной мозг
Рефлекс. Рефлекторная дуга. Спинной мозг Организация внеклассной работы по биологии. Разновозрастной биологический кружок
Организация внеклассной работы по биологии. Разновозрастной биологический кружок Анатомо-физиологические особенности человека в подростковом возрасте
Анатомо-физиологические особенности человека в подростковом возрасте Здоровое питание
Здоровое питание Круговорот веществ в природе
Круговорот веществ в природе Овчарки со всего мира. Фотографии
Овчарки со всего мира. Фотографии Історія виноробства. Класифікація вин
Історія виноробства. Класифікація вин Unusual animals
Unusual animals Белки
Белки Кровообращение как функциональная система. Анатомия сердца и сосудов. Характерные черты строения и функционирования сердечной мышцы
Кровообращение как функциональная система. Анатомия сердца и сосудов. Характерные черты строения и функционирования сердечной мышцы Публичная презентация 2014 год
Публичная презентация 2014 год Бактериофаги. Вирусы бактерий
Бактериофаги. Вирусы бактерий Фотосинтез. 9 класс
Фотосинтез. 9 класс Тварина - живий організм
Тварина - живий організм Доза і основні одиниці вимірювання іонізуючого випромінювання. Основні типи дозових залежностей в радіобіології
Доза і основні одиниці вимірювання іонізуючого випромінювання. Основні типи дозових залежностей в радіобіології