- Археи – уникальная группа организмов

Содержание
- 2. Археи – уникальная группа организмов Клетки 0,1 до 15 мкм диаметром Кокки Бациллы Необычные формы клеток:
- 4. Мембраны архей Разветвленные изопреноиды Простая эфирная связь Трехслойная структура из одного слоя молекул Ассиметричный атом углерода
- 5. Клеточная стенка архей Муреин бактерий заменяется на псевдомуреин
- 6. Клеточная стенка архей S-слои Монослои белков и гликопротеинов, не очень отличных от таковых у бактерий Сложноорганизованная
- 7. Клеточные стенки архей Грам+ археи: псевдомуреин Псевдомуреин + S-слои (белковые субъединицы с гексагональной упаковкой) Грам- археи:
- 8. Морфология архей Включения: Запасные вещества – полифосфаты, элементарная сера, полиглюкозиды Газовые вакуоли (CO2 CH4) Рибосомы архей
- 9. Жгутики архей Присутствуют практически у всех Совершенно иная структура Приспособлены к экстремальным условиям Диаметр жгутиков архей
- 10. Жгутики архей
- 11. Жгутики архей Схожи по строению с пилями IV типа бактерий Отсутствует гомология между структурами жгутика бактерий
- 12. Генетика архей Археи чаще всего содержат одну хромосому, могут содержать плазмиды Упаковка хромосомы может схожа с
- 13. Репликация у архей Может быть больше одной точки ori Репликация требует у архей большое число факторов
- 14. Транскрипция ДНК-зависимая РНК-полимераза у архей одна, но эукариотического типа Промоторные области также схожи с эукариотическими Интроны
- 15. Трансляция Трансляционный аппарат архей также схож с эукариотическим Механизм инициации трансляции различен у разных групп архей
- 16. Метаболизм Фототрофы – галофильные археи Литоавтотрофы и литогетеротрофы Органоавтотрофы и органогетеротрофы По большей части анаэробы
- 17. Катаболизм архей Используют поли- олиго- и моносахара либо полипептиды Катализируют их с помощью гликолиза либо КДФГ-пути*
- 18. Переносчики электронов архей Существуют мобильные, прочно не связанные с мембраной переносчики Кроме кислорода могут использоваться элементарная
- 19. Анаболизм архей Автотрофы фиксируют СО2 в модифицированном цикле Кальвина / цикле Арнора / восстановительный Ацетил-CоА цикл
- 20. Систематика архей Archaea: Тип Euryarchaeota – метаногены, экстремальные термофилы, галоархеи Тип Crenarchaeota – космополиты, мезофилы, гипертермофилы,
- 21. Euryarchaeota Археи-Метаногены Уникальная группа, восстанавливающая CO2 до CH4 окисляя при этом водород Обитают в анаэробных нишах,
- 22. Метаногены Источник углерода – CO2, H2, CO, формиат, метанол, метиламины Автотрофы восстанавливают CO2 до CO и
- 23. Биохимия метаногенеза Восстановление CO2 до метана происходит в 7 стадий Уникальные коферменты Один микроорганизм осуществляет весь
- 24. CHO–MF CHO–H4MPT CH≡H4MPT CH2=H4MPT CH3–H4MPT CH3–SCoM H4MPT CoM–S–S–CoB Тетрагидрометаноптерин гетеродисульфид
- 25. Анаболизм метаногенов Автотрофы фиксируют углекислый газ в модифицированном цикле Арнора Существуют метанокисляющие метаногены
- 26. Euryarchaeota Экстремальные галофилы Порядок Halobacteriales Палочки, кокки, треугольники, квадраты, плоские клетки Грам+ или Грам- Для существования
- 27. Фотосинтез галоархей Фотосинтез – не основной источник энергии Его и фотосинтезом то не все считают Пигменты:
- 29. Галоархейный фотосинтез Нет ЭТЦ Нет хлорофиллов Бактериородопсин формирует пурпурную мембрану Поглощает свет с длиной волны 500-650
- 31. Осмотическая адаптация галоархей Накопление осмопротекторов Структурно и физиологически измененные ферменты, рибосомы и т.д. для работы при
- 32. Crenarchaeota Один класс Thermoprotei и три порядка Кокки, палочки, диски, нити По большей части экстремальные термофилы
- 33. Катаболизм Хемоорганотрофы с аэробным дыханием / анаэробным дыханием (серное) / брожения Хемолитотрофы с анаэробным дыханием (серное,
- 34. Места обитания Черные курильщики Геотермальные источники Вулканические котлы
- 35. Значение кренархеот Источники термостабильных ферментов в молекулярной генетике и органической химии Источник знаний о приспособлениях к
- 37. Скачать презентацию

Археи – уникальная группа организмов
Клетки 0,1 до 15 мкм диаметром
Кокки
Бациллы
Необычные формы
Археи – уникальная группа организмов
Клетки 0,1 до 15 мкм диаметром
Кокки
Бациллы
Необычные формы
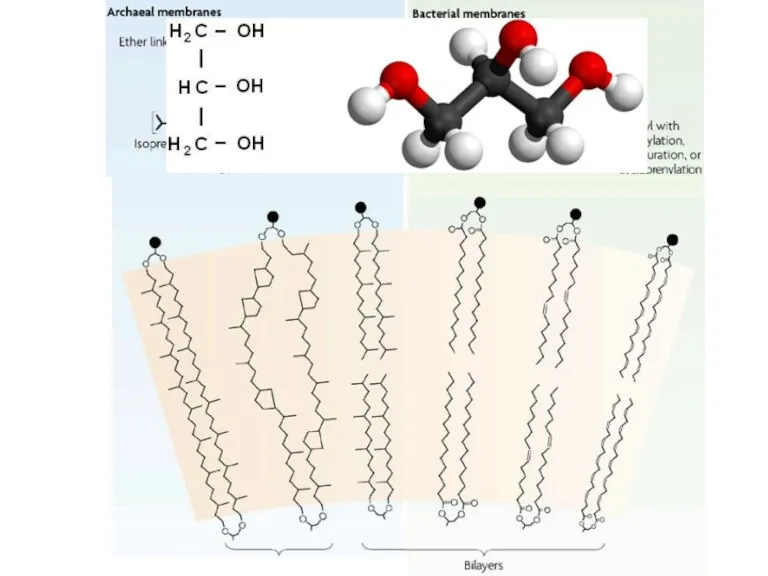

Мембраны архей
Разветвленные изопреноиды
Простая эфирная связь
Трехслойная структура из одного слоя молекул
Ассиметричный атом
Мембраны архей
Разветвленные изопреноиды
Простая эфирная связь
Трехслойная структура из одного слоя молекул
Ассиметричный атом
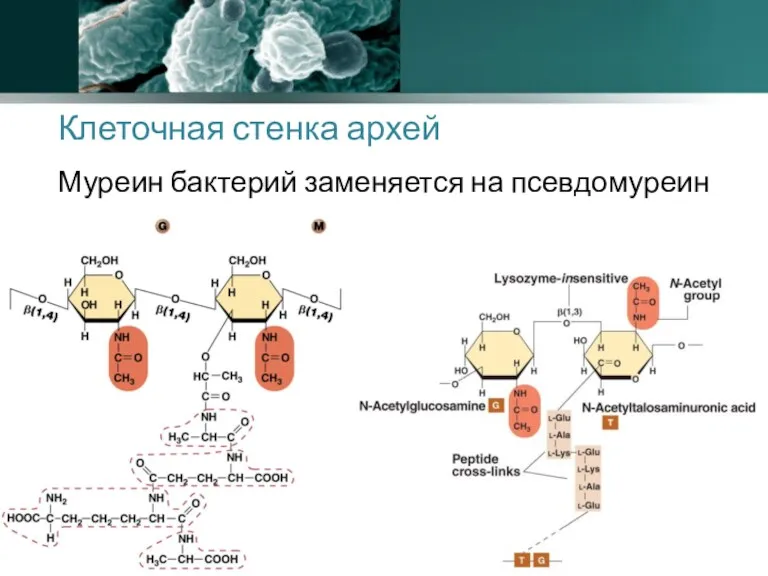
Клеточная стенка архей
Муреин бактерий заменяется на псевдомуреин
Клеточная стенка архей
Муреин бактерий заменяется на псевдомуреин

Клеточная стенка архей
S-слои
Монослои белков и гликопротеинов, не очень отличных от таковых
Клеточная стенка архей
S-слои
Монослои белков и гликопротеинов, не очень отличных от таковых

Клеточные стенки архей
Грам+ археи: псевдомуреин
Псевдомуреин + S-слои (белковые субъединицы с
Клеточные стенки архей
Грам+ археи: псевдомуреин
Псевдомуреин + S-слои (белковые субъединицы с

Морфология архей
Включения:
Запасные вещества – полифосфаты, элементарная сера, полиглюкозиды
Газовые вакуоли (CO2 CH4)
Рибосомы
Морфология архей
Включения:
Запасные вещества – полифосфаты, элементарная сера, полиглюкозиды
Газовые вакуоли (CO2 CH4)
Рибосомы

Жгутики архей
Присутствуют практически у всех
Совершенно иная структура
Приспособлены к экстремальным условиям
Диаметр жгутиков
Жгутики архей
Присутствуют практически у всех
Совершенно иная структура
Приспособлены к экстремальным условиям
Диаметр жгутиков

Жгутики архей
Жгутики архей

Жгутики архей
Схожи по строению с пилями IV типа бактерий
Отсутствует гомология между
Жгутики архей
Схожи по строению с пилями IV типа бактерий
Отсутствует гомология между

Генетика архей
Археи чаще всего содержат одну хромосому, могут содержать плазмиды
Упаковка хромосомы
Генетика архей
Археи чаще всего содержат одну хромосому, могут содержать плазмиды
Упаковка хромосомы

Репликация у архей
Может быть больше одной точки ori
Репликация требует у архей
Репликация у архей
Может быть больше одной точки ori
Репликация требует у архей

Транскрипция
ДНК-зависимая РНК-полимераза у архей одна, но эукариотического типа
Промоторные области также схожи
Транскрипция
ДНК-зависимая РНК-полимераза у архей одна, но эукариотического типа
Промоторные области также схожи

Трансляция
Трансляционный аппарат архей также схож с эукариотическим
Механизм инициации трансляции различен у
Трансляция
Трансляционный аппарат архей также схож с эукариотическим
Механизм инициации трансляции различен у

Метаболизм
Фототрофы – галофильные археи
Литоавтотрофы и литогетеротрофы
Органоавтотрофы и органогетеротрофы
По большей части анаэробы
Метаболизм
Фототрофы – галофильные археи
Литоавтотрофы и литогетеротрофы
Органоавтотрофы и органогетеротрофы
По большей части анаэробы

Катаболизм архей
Используют поли- олиго- и моносахара либо полипептиды
Катализируют их с
Катаболизм архей
Используют поли- олиго- и моносахара либо полипептиды
Катализируют их с

Переносчики электронов архей
Существуют мобильные, прочно не связанные с мембраной переносчики
Кроме кислорода
Переносчики электронов архей
Существуют мобильные, прочно не связанные с мембраной переносчики
Кроме кислорода

Анаболизм архей
Автотрофы фиксируют СО2 в модифицированном цикле Кальвина / цикле Арнора
Анаболизм архей
Автотрофы фиксируют СО2 в модифицированном цикле Кальвина / цикле Арнора

Систематика архей
Archaea:
Тип Euryarchaeota – метаногены, экстремальные термофилы, галоархеи
Тип Crenarchaeota – космополиты,
Систематика архей
Archaea:
Тип Euryarchaeota – метаногены, экстремальные термофилы, галоархеи
Тип Crenarchaeota – космополиты,

Euryarchaeota
Археи-Метаногены
Уникальная группа, восстанавливающая CO2 до CH4 окисляя при этом водород
Обитают
Euryarchaeota
Археи-Метаногены
Уникальная группа, восстанавливающая CO2 до CH4 окисляя при этом водород
Обитают

Метаногены
Источник углерода – CO2, H2, CO, формиат, метанол, метиламины
Автотрофы восстанавливают CO2
Метаногены
Источник углерода – CO2, H2, CO, формиат, метанол, метиламины
Автотрофы восстанавливают CO2

Биохимия метаногенеза
Восстановление CO2 до метана происходит в 7 стадий
Уникальные коферменты
Один
Биохимия метаногенеза
Восстановление CO2 до метана происходит в 7 стадий
Уникальные коферменты
Один

CHO–MF
CHO–H4MPT
CH≡H4MPT
CH2=H4MPT
CH3–H4MPT
CH3–SCoM
H4MPT
CoM–S–S–CoB
Тетрагидрометаноптерин
гетеродисульфид
CHO–MF
CHO–H4MPT
CH≡H4MPT
CH2=H4MPT
CH3–H4MPT
CH3–SCoM
H4MPT
CoM–S–S–CoB
Тетрагидрометаноптерин
гетеродисульфид

Анаболизм метаногенов
Автотрофы фиксируют углекислый газ в модифицированном цикле Арнора
Существуют метанокисляющие метаногены
Анаболизм метаногенов
Автотрофы фиксируют углекислый газ в модифицированном цикле Арнора
Существуют метанокисляющие метаногены

Euryarchaeota
Экстремальные галофилы
Порядок Halobacteriales
Палочки, кокки, треугольники, квадраты, плоские клетки
Грам+ или Грам-
Для
Euryarchaeota
Экстремальные галофилы
Порядок Halobacteriales
Палочки, кокки, треугольники, квадраты, плоские клетки
Грам+ или Грам-
Для

Фотосинтез галоархей
Фотосинтез – не основной источник энергии
Его и фотосинтезом то не
Фотосинтез галоархей
Фотосинтез – не основной источник энергии
Его и фотосинтезом то не
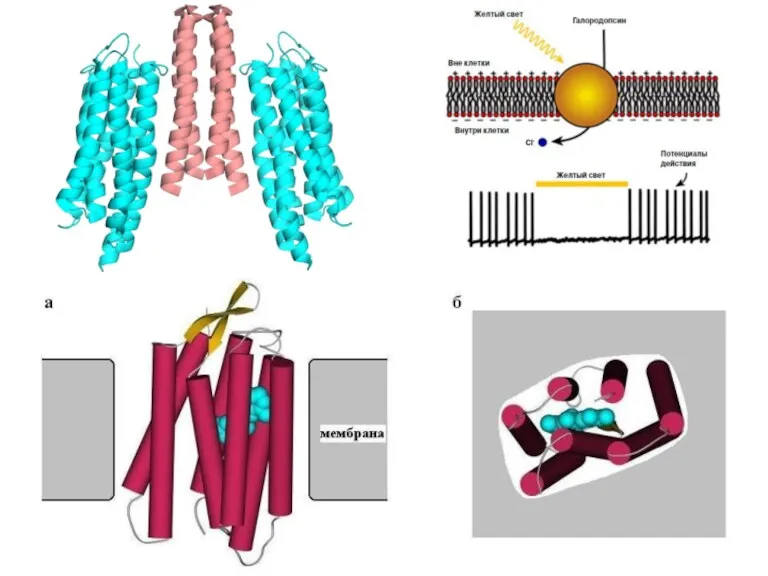
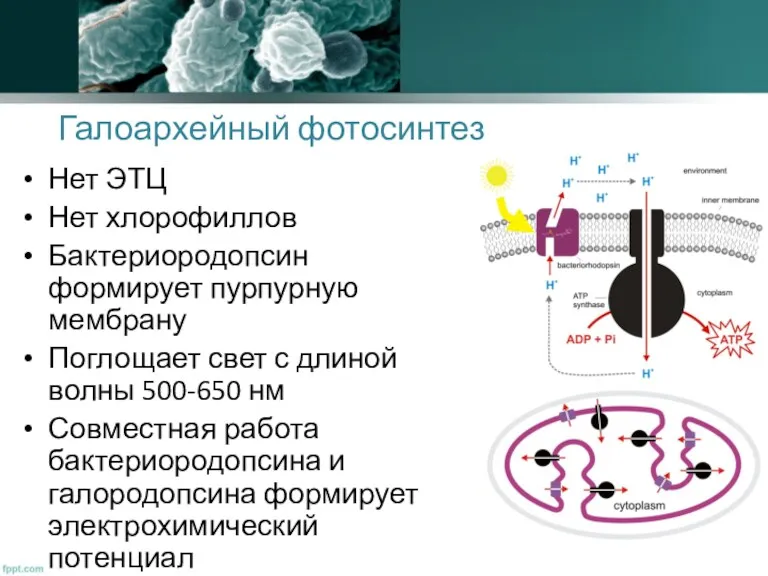
Галоархейный фотосинтез
Нет ЭТЦ
Нет хлорофиллов
Бактериородопсин формирует пурпурную мембрану
Поглощает свет с длиной волны
Галоархейный фотосинтез
Нет ЭТЦ
Нет хлорофиллов
Бактериородопсин формирует пурпурную мембрану
Поглощает свет с длиной волны

Осмотическая адаптация галоархей
Накопление осмопротекторов
Структурно и физиологически измененные ферменты, рибосомы и т.д.
Осмотическая адаптация галоархей
Накопление осмопротекторов
Структурно и физиологически измененные ферменты, рибосомы и т.д.

Crenarchaeota
Один класс Thermoprotei и три порядка
Кокки, палочки, диски, нити
По большей части
Crenarchaeota
Один класс Thermoprotei и три порядка
Кокки, палочки, диски, нити
По большей части

Катаболизм
Хемоорганотрофы с аэробным дыханием / анаэробным дыханием (серное) / брожения
Хемолитотрофы с
Катаболизм
Хемоорганотрофы с аэробным дыханием / анаэробным дыханием (серное) / брожения
Хемолитотрофы с

Места обитания
Черные курильщики
Геотермальные источники
Вулканические котлы
Места обитания
Черные курильщики
Геотермальные источники
Вулканические котлы

Значение кренархеот
Источники термостабильных ферментов в молекулярной генетике и органической химии
Источник знаний
Значение кренархеот
Источники термостабильных ферментов в молекулярной генетике и органической химии
Источник знаний
 Строение ядра. Хромосомы
Строение ядра. Хромосомы Класс Птицы
Класс Птицы Растительные сообщества и взаимосвязи в них
Растительные сообщества и взаимосвязи в них Детям о насекомых
Детям о насекомых Морфологический критерий вида
Морфологический критерий вида Зрение. Строение глаза
Зрение. Строение глаза Оплодотворение у цветковых растений
Оплодотворение у цветковых растений Классификация животных и растений. Эволюция
Классификация животных и растений. Эволюция Тип членистоногие
Тип членистоногие Микроскопические частицы вирусы
Микроскопические частицы вирусы Грибы - часть живой природы
Грибы - часть живой природы Класс Хрящевые рыбы
Класс Хрящевые рыбы Генетика аддиктивного поведения. Преступность и наследственность
Генетика аддиктивного поведения. Преступность и наследственность Мембраналардың иондық каналдары
Мембраналардың иондық каналдары Урок-игра по теме В мире рыб
Урок-игра по теме В мире рыб Легенева вентиляція. Регуляція дихання
Легенева вентиляція. Регуляція дихання Селекцiя
Селекцiя Строение и функции скелета человека
Строение и функции скелета человека Хрящевые рыбы
Хрящевые рыбы Презентация Сообщества воды 5 класс
Презентация Сообщества воды 5 класс Кинетические дисфункции СБС
Кинетические дисфункции СБС Фитотерапия - лечение лекарственными травами. 1 класс
Фитотерапия - лечение лекарственными травами. 1 класс Semenné rostliny Spermatophyta
Semenné rostliny Spermatophyta Растения-хищники. 6 класс
Растения-хищники. 6 класс Царства живой природы
Царства живой природы Поведінка
Поведінка Анатомия и физиология человека
Анатомия и физиология человека Carbohydrate Metabolism I: Aerobic oxidation of glucose. Anaerobic Glycolysis. Gluconeogenesis
Carbohydrate Metabolism I: Aerobic oxidation of glucose. Anaerobic Glycolysis. Gluconeogenesis