- Бактериофаги (или просто фаги) – вирусы бактерий

Содержание
- 2. Типичная форма фагов Ножка нужна, чтобы прикрепляться к бактериальной поверхности и «протыкать» ее. Далее из тела
- 3. Фаги бывают очень разные… Но смысл их существования всегда одинаков: Найти подходящую бактерию и впрыснуть в
- 4. Схема жизни фага А вот схема экспрессии фаговых генов внутри бактериальной клетки.
- 5. Фаги с одноцепочечной кольцевой ДНК Икосаэдрические фаги (φX174) Обладают типичной сферической формой, связываются с поверхностью бактерии,
- 6. Выход филаментных фагов из бактериальной клетки Процесс обеспечивается фаговыми белками. Фаговая частица собирается в периплазме, в
- 7. Репликация у оцДНК-фагов Первый этап репликации – синтез так называемой репликативной формы ДНК (RF). В клетку
- 8. Репликация у оцДНК-фагов С первой же синтезированной в клетке RF начинает экспрессироваться ген II, кодирующий эндонуклеазу.
- 9. Фаги с двуцепочечной линейной ДНК Чаще всего если фаг маленький, то его ДНК кольцевая, а если
- 10. Фаги с двуцепочечной линейной ДНК Эукариоты решают эту проблему при помощи теломеразы, но у фагов ничего
- 11. Фаги с двуцепочечной линейной ДНК – T3, T7 Так это же липкие концы! Они и слипаются,
- 12. Фаги с двуцепочечной линейной ДНК – T4 Это большой фаг, его ДНК размером 170 т.п.н. кодирует
- 13. Фаги с двуцепочечной линейной ДНК – T4 У ДНК фага Т4 на концах тоже повторы. Репликация
- 14. Фаговый лизис Как мы уже знаем, некоторые фаги не убивают клетку, которую покидают. Однако же большинство
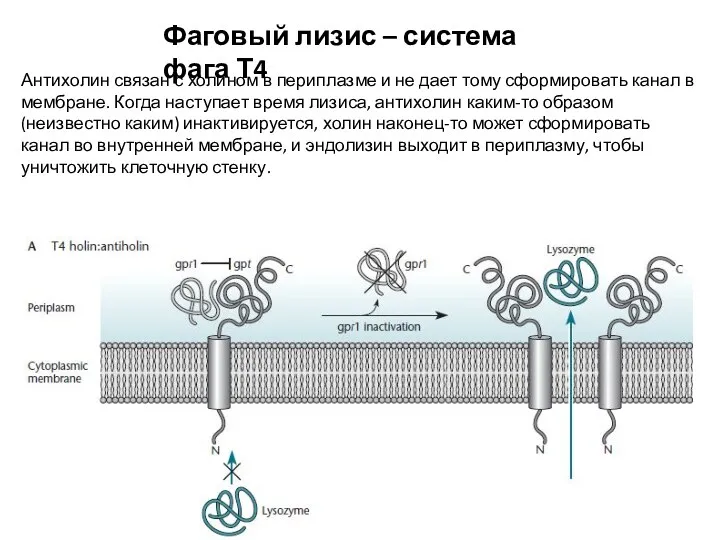
- 15. Фаговый лизис – система фага Т4 Антихолин связан с холином в периплазме и не дает тому
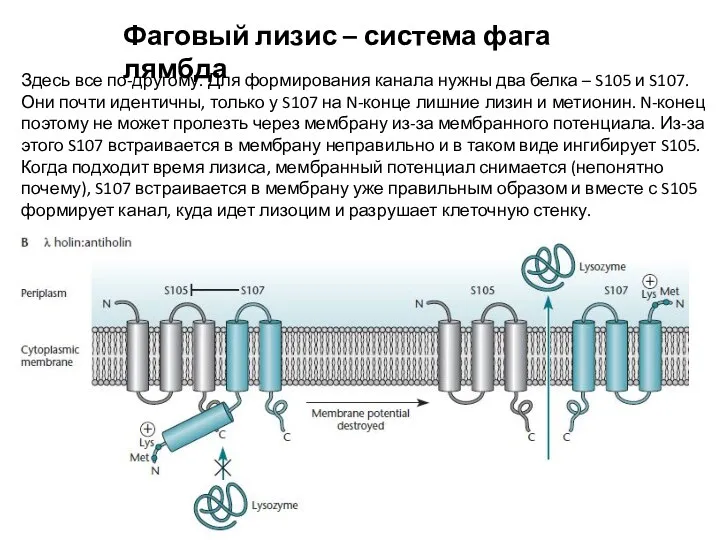
- 16. Фаговый лизис – система фага лямбда Здесь все по-другому. Для формирования канала нужны два белка –
- 17. Бактериальные системы борьбы с фагами Если бы бактерии не умели сопротивляться фагам – и тех и
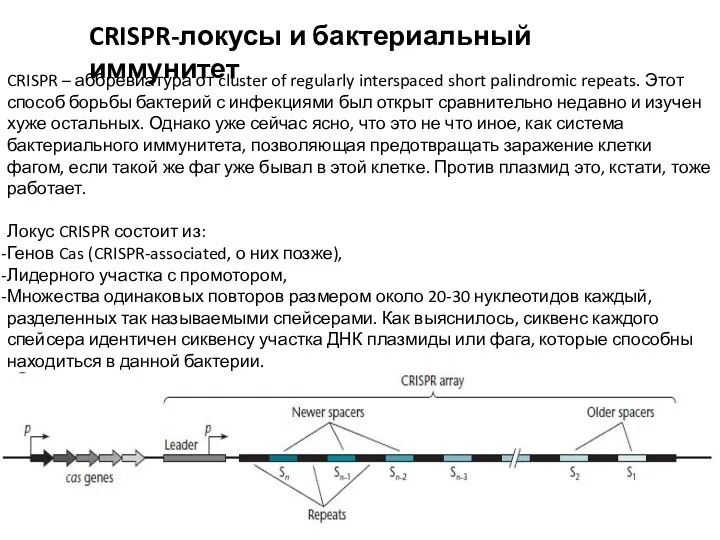
- 18. CRISPR-локусы и бактериальный иммунитет CRISPR – аббревиатура от cluster of regularly interspaced short palindromic repeats. Этот
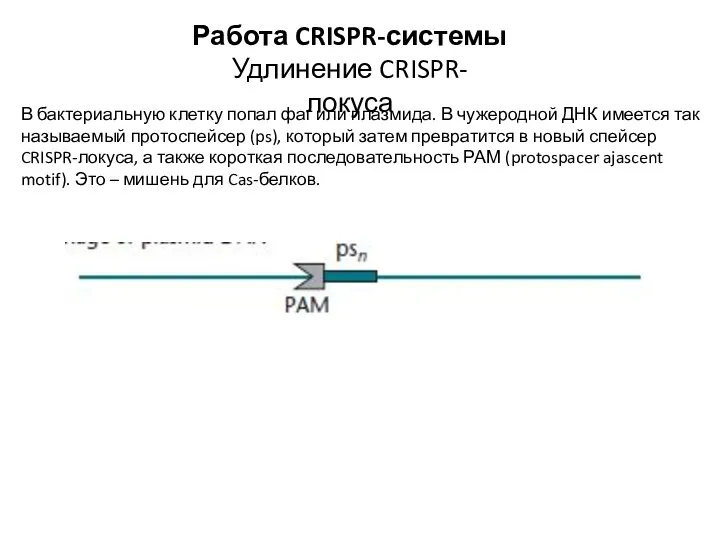
- 19. Работа CRISPR-системы Удлинение CRISPR-локуса В бактериальную клетку попал фаг или плазмида. В чужеродной ДНК имеется так
- 20. Работа CRISPR-системы Удлинение CRISPR-локуса (адаптивная фаза) Cas-белки узнают РАМ в ДНК-пришельце и вырезают из него протоспейсер,
- 22. Скачать презентацию
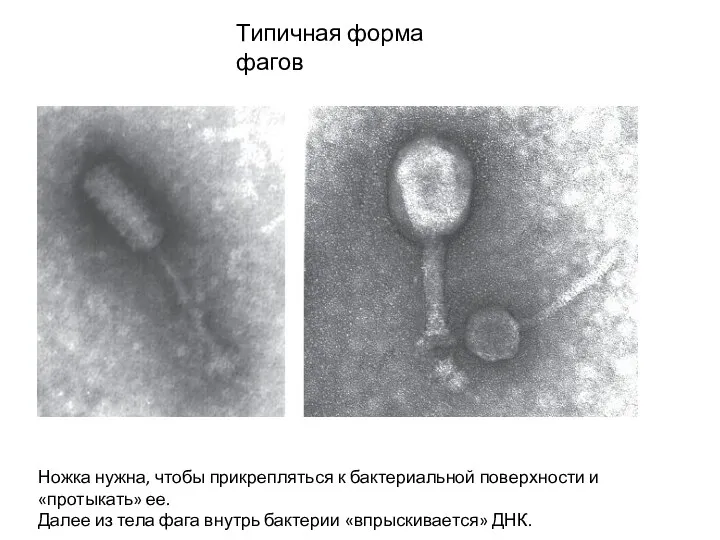
Типичная форма фагов
Ножка нужна, чтобы прикрепляться к бактериальной поверхности и «протыкать»
Типичная форма фагов
Ножка нужна, чтобы прикрепляться к бактериальной поверхности и «протыкать»
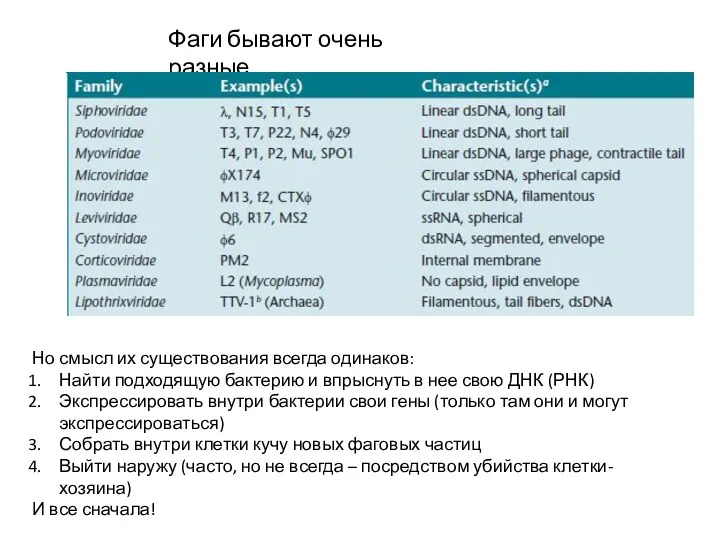
Фаги бывают очень разные…
Но смысл их существования всегда одинаков:
Найти подходящую бактерию
Фаги бывают очень разные…
Но смысл их существования всегда одинаков:
Найти подходящую бактерию
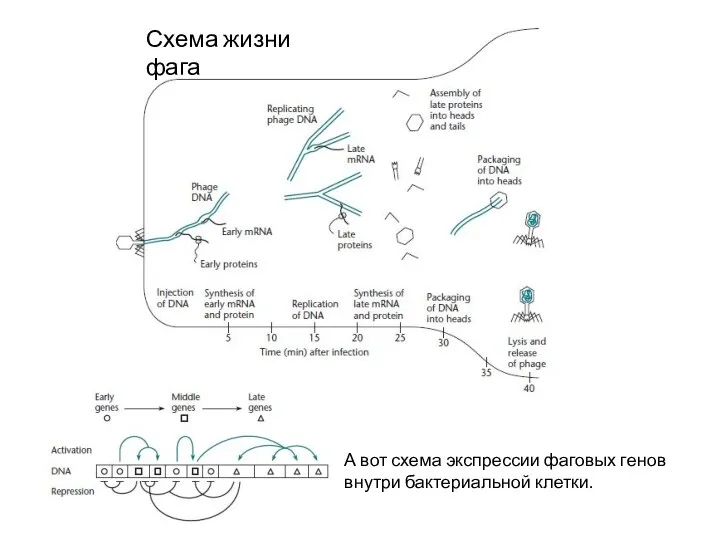
Схема жизни фага
А вот схема экспрессии фаговых генов внутри бактериальной клетки.
Схема жизни фага
А вот схема экспрессии фаговых генов внутри бактериальной клетки.
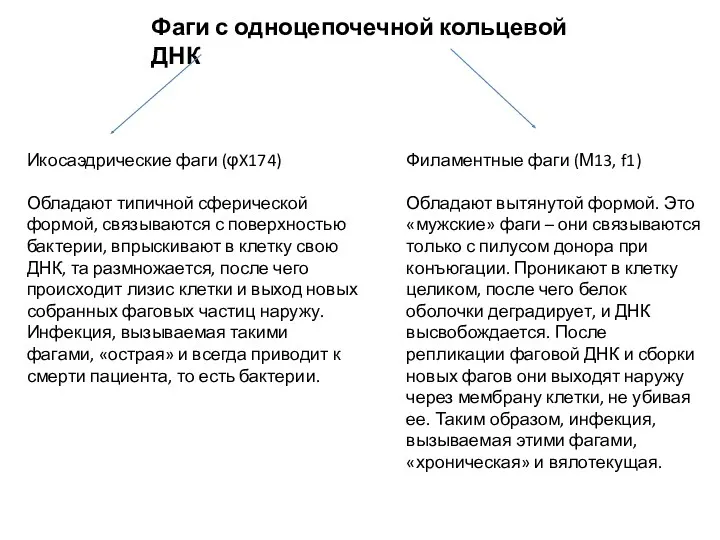
Фаги с одноцепочечной кольцевой ДНК
Икосаэдрические фаги (φX174)
Обладают типичной сферической формой, связываются
Фаги с одноцепочечной кольцевой ДНК
Икосаэдрические фаги (φX174)
Обладают типичной сферической формой, связываются
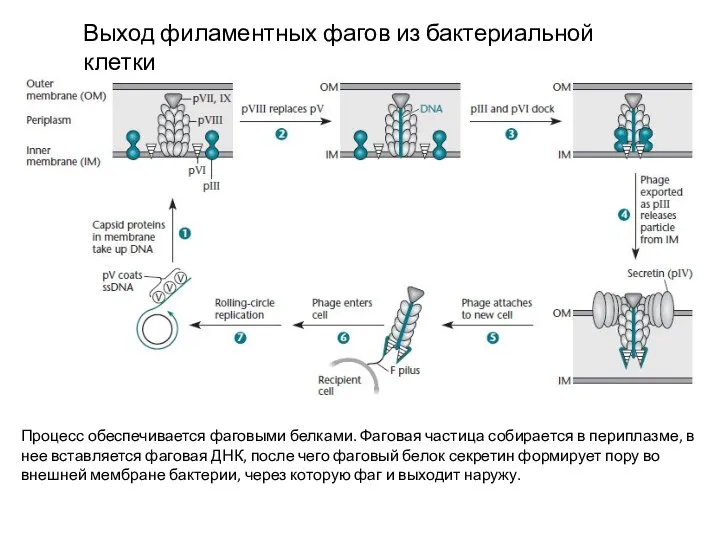
Выход филаментных фагов из бактериальной клетки
Процесс обеспечивается фаговыми белками. Фаговая частица
Выход филаментных фагов из бактериальной клетки
Процесс обеспечивается фаговыми белками. Фаговая частица
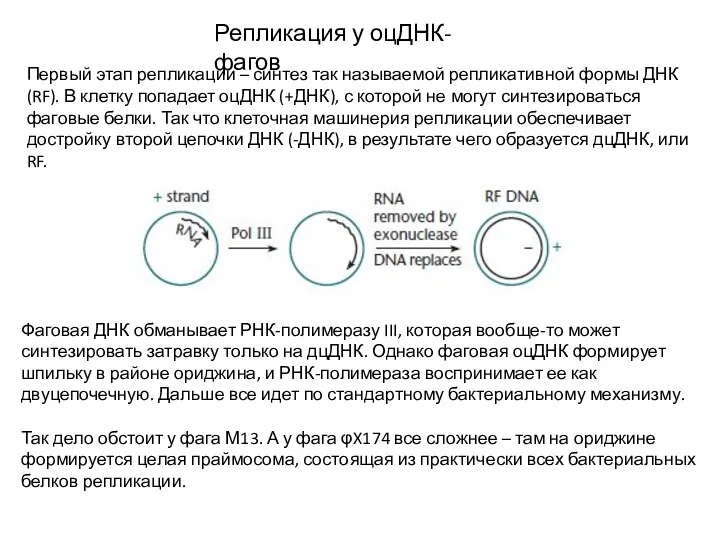
Репликация у оцДНК-фагов
Первый этап репликации – синтез так называемой репликативной формы
Репликация у оцДНК-фагов
Первый этап репликации – синтез так называемой репликативной формы
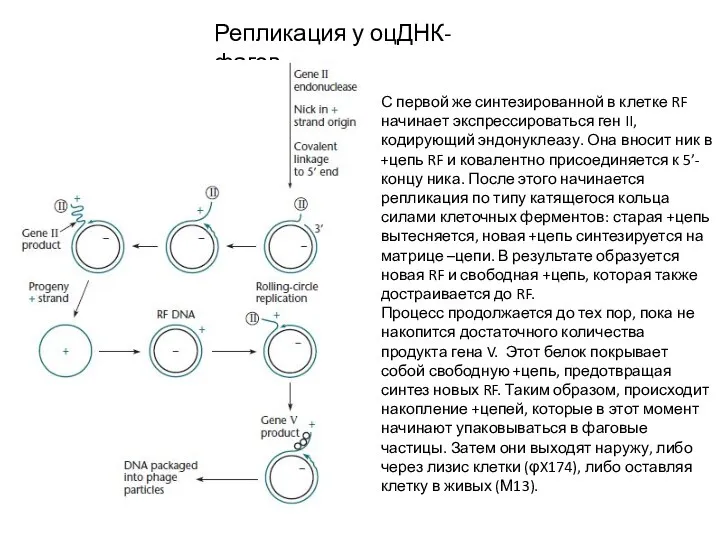
Репликация у оцДНК-фагов
С первой же синтезированной в клетке RF начинает экспрессироваться
Репликация у оцДНК-фагов
С первой же синтезированной в клетке RF начинает экспрессироваться
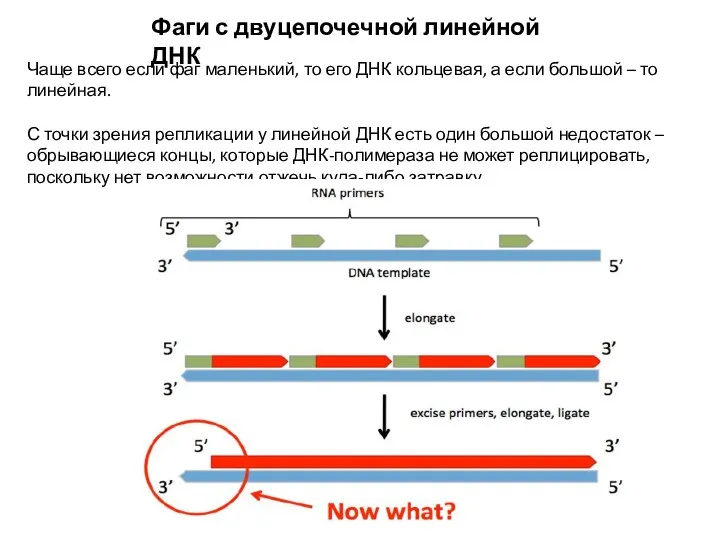
Фаги с двуцепочечной линейной ДНК
Чаще всего если фаг маленький, то его
Фаги с двуцепочечной линейной ДНК
Чаще всего если фаг маленький, то его
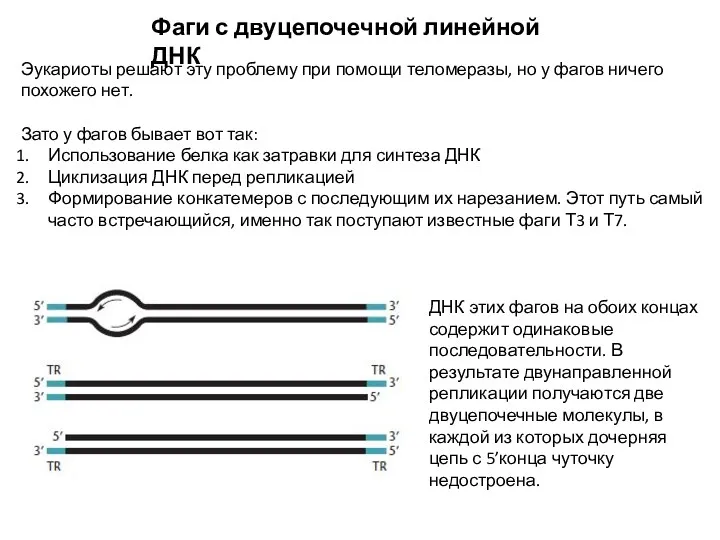
Фаги с двуцепочечной линейной ДНК
Эукариоты решают эту проблему при помощи теломеразы,
Фаги с двуцепочечной линейной ДНК
Эукариоты решают эту проблему при помощи теломеразы,
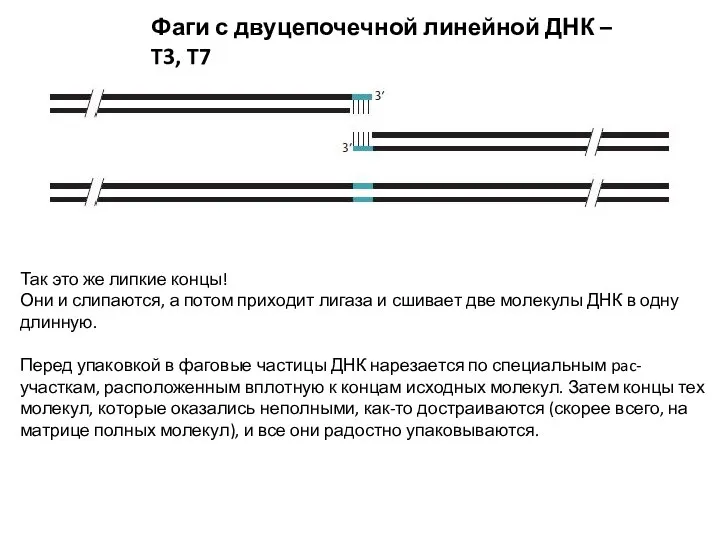
Фаги с двуцепочечной линейной ДНК – T3, T7
Так это же липкие
Фаги с двуцепочечной линейной ДНК – T3, T7
Так это же липкие
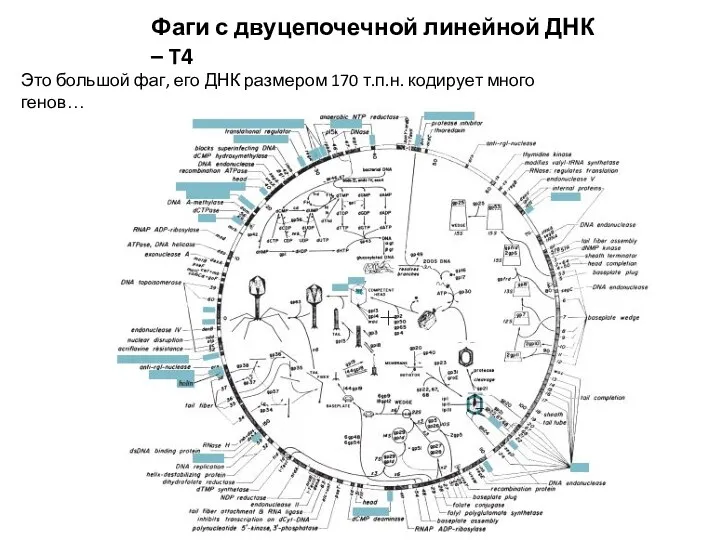
Фаги с двуцепочечной линейной ДНК – T4
Это большой фаг, его ДНК
Фаги с двуцепочечной линейной ДНК – T4
Это большой фаг, его ДНК
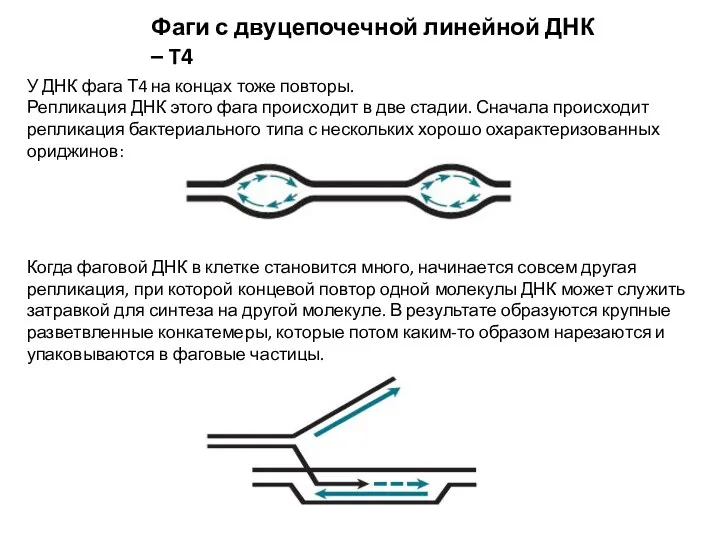
Фаги с двуцепочечной линейной ДНК – T4
У ДНК фага Т4 на
Фаги с двуцепочечной линейной ДНК – T4
У ДНК фага Т4 на

Фаговый лизис
Как мы уже знаем, некоторые фаги не убивают клетку, которую
Фаговый лизис
Как мы уже знаем, некоторые фаги не убивают клетку, которую
Фаговый лизис – система фага Т4
Антихолин связан с холином в периплазме
Фаговый лизис – система фага Т4
Антихолин связан с холином в периплазме
Фаговый лизис – система фага лямбда
Здесь все по-другому. Для формирования канала
Фаговый лизис – система фага лямбда
Здесь все по-другому. Для формирования канала

Бактериальные системы борьбы с фагами
Если бы бактерии не умели сопротивляться фагам
Бактериальные системы борьбы с фагами
Если бы бактерии не умели сопротивляться фагам
CRISPR-локусы и бактериальный иммунитет
CRISPR – аббревиатура от cluster of regularly interspaced
CRISPR-локусы и бактериальный иммунитет
CRISPR – аббревиатура от cluster of regularly interspaced
Работа CRISPR-системы
Удлинение CRISPR-локуса
В бактериальную клетку попал фаг или плазмида. В чужеродной
Работа CRISPR-системы
Удлинение CRISPR-локуса
В бактериальную клетку попал фаг или плазмида. В чужеродной
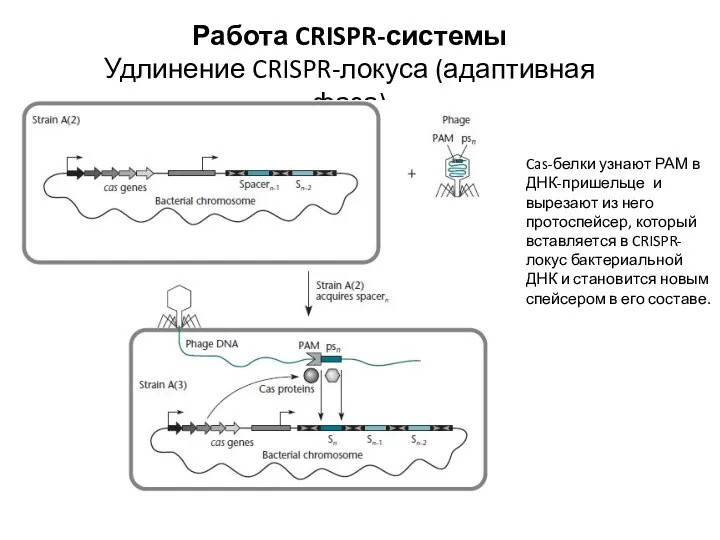
Работа CRISPR-системы
Удлинение CRISPR-локуса (адаптивная фаза)
Cas-белки узнают РАМ в ДНК-пришельце и вырезают
Работа CRISPR-системы
Удлинение CRISPR-локуса (адаптивная фаза)
Cas-белки узнают РАМ в ДНК-пришельце и вырезают
 Декоративные растения. Основы ландшафтной композиции
Декоративные растения. Основы ландшафтной композиции Кісткові риби
Кісткові риби Общие закономерности адаптации организма к различным условиям
Общие закономерности адаптации организма к различным условиям Модификационная изменчивость
Модификационная изменчивость Геоботаник из села Усть-Река. Тихомиров Борис Анатольевич
Геоботаник из села Усть-Река. Тихомиров Борис Анатольевич Гетерозис ғылымының теориясы мен болжамы
Гетерозис ғылымының теориясы мен болжамы Нейроглия. Гематоэнцефалический барьер
Нейроглия. Гематоэнцефалический барьер Бактериофаг құрылысы және маңызы
Бактериофаг құрылысы және маңызы Чудесные Цветники осенью
Чудесные Цветники осенью урок биологии в 8 классе Живая крепость
урок биологии в 8 классе Живая крепость Домашние мелкие грызуны и необходимые им минералы
Домашние мелкие грызуны и необходимые им минералы Митоз
Митоз В гости к осени. 2 класс
В гости к осени. 2 класс Мастер-класс. Структурно-логические технологиии(применение на уроках биологии схем,таблиц, разноуровневых заданий).
Мастер-класс. Структурно-логические технологиии(применение на уроках биологии схем,таблиц, разноуровневых заданий). Экологические сообщества. Экосистема, биогеоценоз
Экологические сообщества. Экосистема, биогеоценоз Urinary system
Urinary system Різноманітність ракоподібних
Різноманітність ракоподібних Физиологические механизмы слуховой и вестибулярной сенсорных систем
Физиологические механизмы слуховой и вестибулярной сенсорных систем Мәдени өсімдіктер мен үй жануарларының шығу орталықтары
Мәдени өсімдіктер мен үй жануарларының шығу орталықтары Шишковидное тело (corpus pineale)
Шишковидное тело (corpus pineale) Современные представления о физиологии синапсов
Современные представления о физиологии синапсов Внутренняя среда организма. Кровь, тканевая жидкость и лимфа
Внутренняя среда организма. Кровь, тканевая жидкость и лимфа Семейство Соколиные. Совы
Семейство Соколиные. Совы Ткани внутренней среды организма. Костная ткань
Ткани внутренней среды организма. Костная ткань Будова і функції скелета
Будова і функції скелета Плоды. Строение плода. Классификация плодов
Плоды. Строение плода. Классификация плодов Влияние сроков посева на рост и развитие сеянцев сосны и ели с закрытой корневой системой
Влияние сроков посева на рост и развитие сеянцев сосны и ели с закрытой корневой системой Общая характеристика грибов
Общая характеристика грибов