- Байгабилов Д., 2 курс

Содержание
- 2. Введение Цитоплазматические органеллы, осуществляющие биоэнергетические процессы в клетках растений и животных, были открыты в 1841 году;
- 3. Митохондрии, присутствуют у большинства животных клеток (исключение составляют лишь эритроциты крови). Митохондрии являются наиболее распространенным видом
- 4. Митохондриальный геном включает 1000-2000 клеточных ядерных генов, кодирующий наиболее важные гены, связанные с биоэнергетикой клеток. Собственный
- 5. Ядерные и митохондриальные гены совместно участвуют в процессах транскрипции, трансляции белков и в дыхании. В мтДНК
- 6. Наличие мутаций в мтДНК было обнаружено в 1988 году. Наследственные или приобретенные мутации в мтДНК (точечные,
- 7. В формировании митохондриального генома будущего ребенка мтДНК отца не участвует. Все мутации и нарушения в митохондриях
- 8. Гетерогенность внутриклеточной популяции митохондрий Митохондриальная популяция единичной клетки состоит из смеси митохондрий разнородной морфологии. Гетерогенность митохондриальной
- 9. Митохондрии растений, в отличие от митохондрий дрожжей и млекопитающих, не формируют непрерывную сеть, являются очень динамичными
- 10. Объектами исследования были митохондрии, изолированные из двух типов растительной тканей: прорастающей (этиолированные 3х-дневные проростки кукурузы Zea
- 11. Данные - Фракция 1 локализовалась на уровне 0,3-0,4 М концентрации сахарозы или на границе между 18%
- 12. 1) C помощью электрофореза в ПАА-геле в денатурирующих условиях были обнаружены незначительные отличия в белковых спектрах
- 13. 3) Анализ активности дыхательных комплексов с помощью метода BN-PAGE показал, что 1-я митохондриальная фракция во всех
- 14. С целью дальнейшей функциональной характеристики различных митохондриальных субпопуляций в клетке было проведено исследование эффективности импорта ДНК
- 15. Следует отметить, что разница между уровнем импорта ДНК в 1-ю и 3-ю фракции митохондрий репы (в
- 16. Феномен различия в активности импорта ДНК в различные митохондриальные субпопуляции может быть обусловлен тем, что митохондрии
- 17. Заключение Митохондрии — органеллы энергообеспечения метаболических процесов в клетке. ДНК митохондрий наследуются почти исключительно по материнской
- 19. Скачать презентацию

Введение
Цитоплазматические органеллы, осуществляющие биоэнергетические процессы в клетках растений и животных,
Введение
Цитоплазматические органеллы, осуществляющие биоэнергетические процессы в клетках растений и животных,
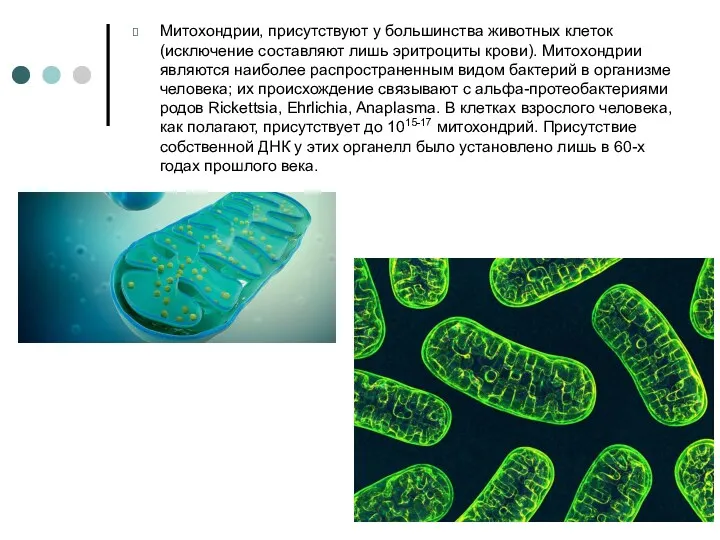
Митохондрии, присутствуют у большинства животных клеток (исключение составляют лишь эритроциты крови).
Митохондрии, присутствуют у большинства животных клеток (исключение составляют лишь эритроциты крови).
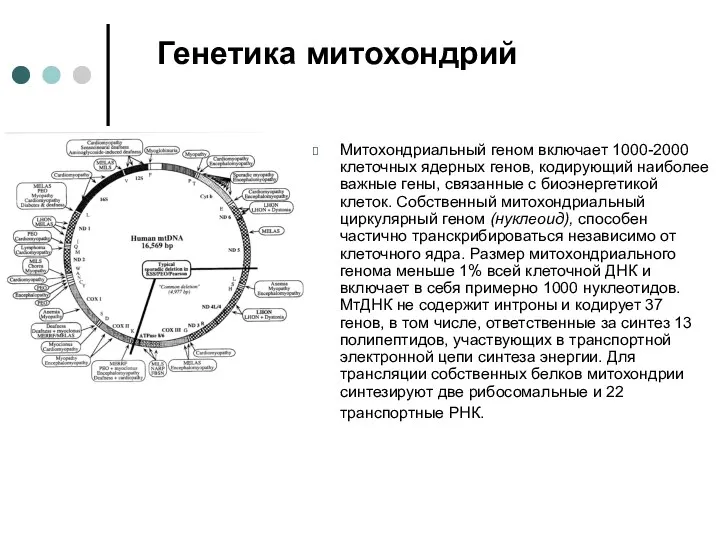
Митохондриальный геном включает 1000-2000 клеточных ядерных генов, кодирующий наиболее важные гены,
Митохондриальный геном включает 1000-2000 клеточных ядерных генов, кодирующий наиболее важные гены,
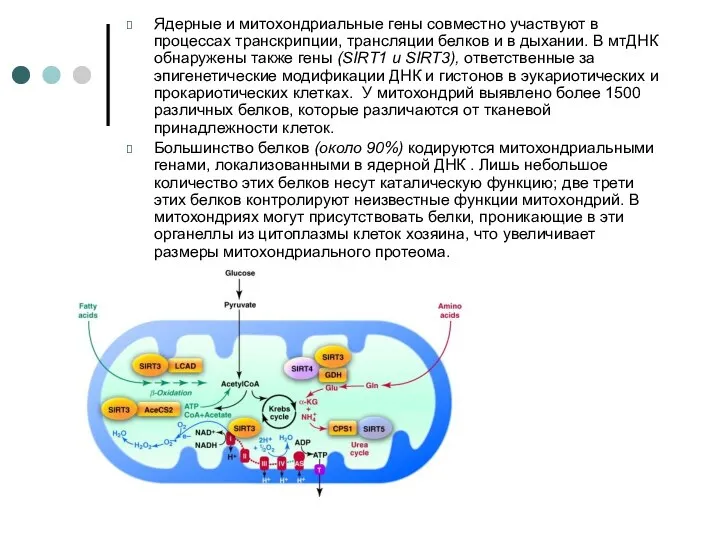
Ядерные и митохондриальные гены совместно участвуют в процессах транскрипции, трансляции белков
Ядерные и митохондриальные гены совместно участвуют в процессах транскрипции, трансляции белков
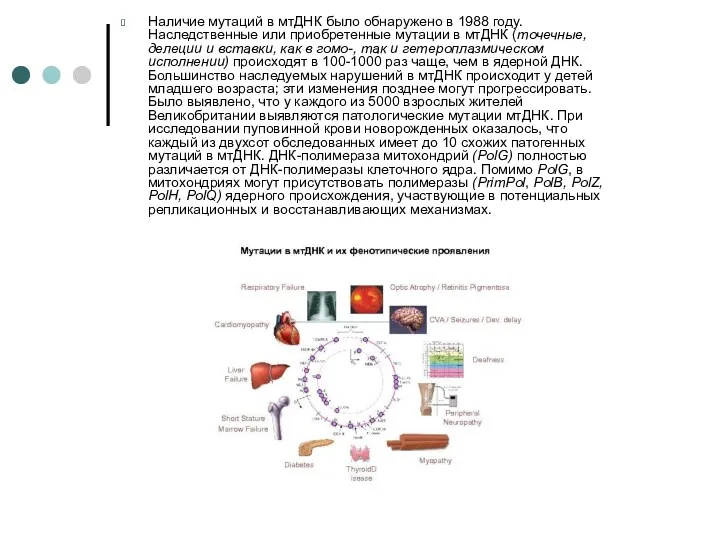
Наличие мутаций в мтДНК было обнаружено в 1988 году. Наследственные или
Наличие мутаций в мтДНК было обнаружено в 1988 году. Наследственные или

В формировании митохондриального генома будущего ребенка мтДНК отца не участвует. Все
В формировании митохондриального генома будущего ребенка мтДНК отца не участвует. Все
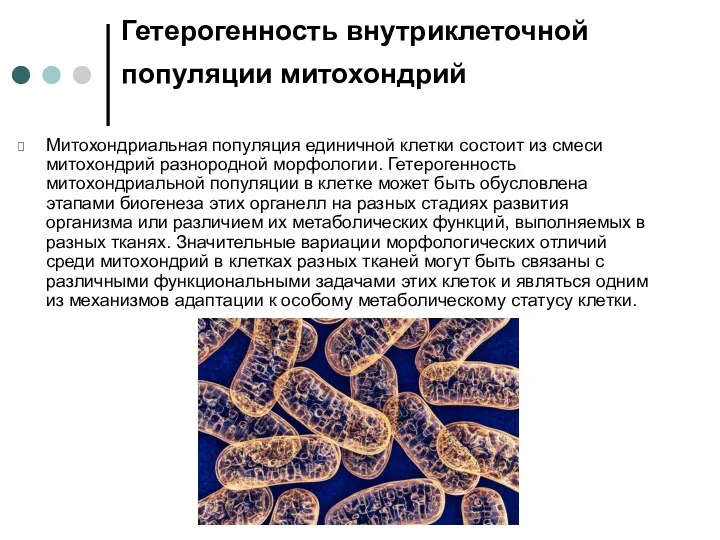
Гетерогенность внутриклеточной популяции митохондрий
Митохондриальная популяция единичной клетки состоит из смеси
Гетерогенность внутриклеточной популяции митохондрий
Митохондриальная популяция единичной клетки состоит из смеси
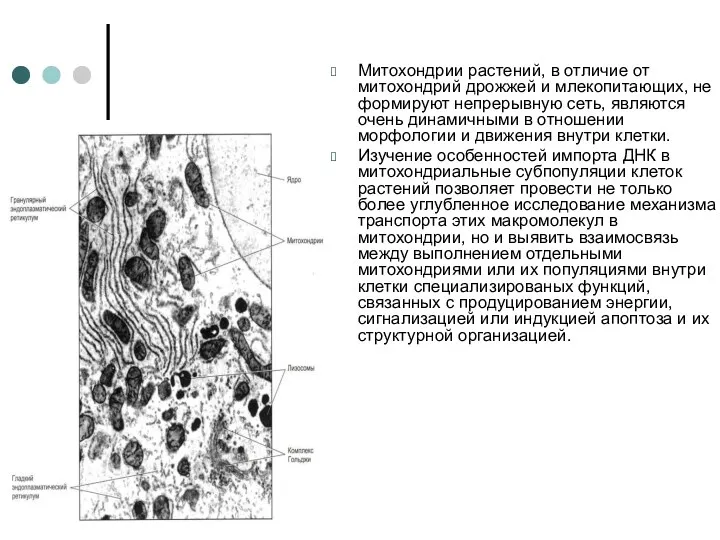
Митохондрии растений, в отличие от митохондрий дрожжей и млекопитающих, не формируют
Митохондрии растений, в отличие от митохондрий дрожжей и млекопитающих, не формируют
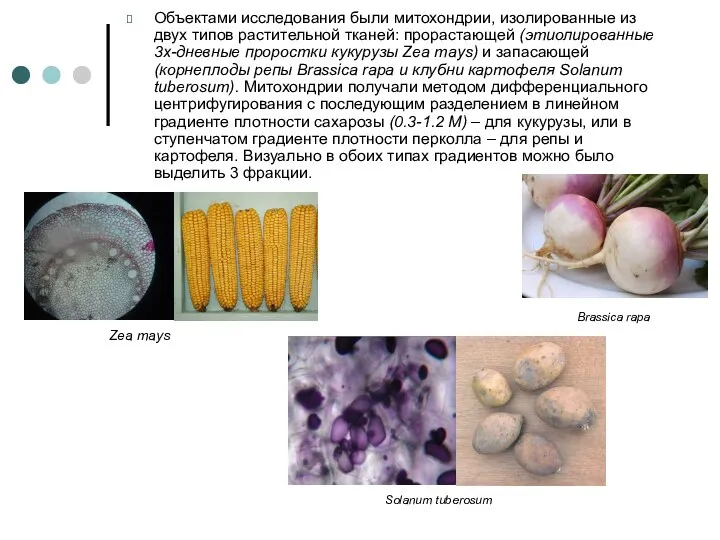
Объектами исследования были митохондрии, изолированные из двух типов растительной тканей: прорастающей
Объектами исследования были митохондрии, изолированные из двух типов растительной тканей: прорастающей

Данные
- Фракция 1 локализовалась на уровне 0,3-0,4 М концентрации сахарозы или
Данные
- Фракция 1 локализовалась на уровне 0,3-0,4 М концентрации сахарозы или

1) C помощью электрофореза в ПАА-геле в денатурирующих условиях были обнаружены
1) C помощью электрофореза в ПАА-геле в денатурирующих условиях были обнаружены

3) Анализ активности дыхательных комплексов с помощью метода BN-PAGE показал, что
3) Анализ активности дыхательных комплексов с помощью метода BN-PAGE показал, что

С целью дальнейшей функциональной характеристики различных митохондриальных субпопуляций в клетке было
С целью дальнейшей функциональной характеристики различных митохондриальных субпопуляций в клетке было

Следует отметить, что разница между уровнем импорта ДНК в 1-ю и
Следует отметить, что разница между уровнем импорта ДНК в 1-ю и

Феномен различия в активности импорта ДНК в различные митохондриальные субпопуляции может
Феномен различия в активности импорта ДНК в различные митохондриальные субпопуляции может

Заключение
Митохондрии — органеллы энергообеспечения метаболических процесов в клетке.
ДНК митохондрий наследуются почти исключительно по
Заключение
Митохондрии — органеллы энергообеспечения метаболических процесов в клетке. ДНК митохондрий наследуются почти исключительно по
 Органы чуств. Регуляция деятельности организма
Органы чуств. Регуляция деятельности организма Обеспечение клеток энергией. Фотосинтез
Обеспечение клеток энергией. Фотосинтез Скелет головы и скелет туловища
Скелет головы и скелет туловища Звук. Звуковая волна. Уши и слух
Звук. Звуковая волна. Уши и слух Высшие растения. Споровые: мхи, папоротники, хвощи и плауны
Высшие растения. Споровые: мхи, папоротники, хвощи и плауны Выделение веществ как важнейший процесс жизнедеятельности организмов. 6 класс
Выделение веществ как важнейший процесс жизнедеятельности организмов. 6 класс Физико-химические свойства крови
Физико-химические свойства крови Органические и неорганические вещества входящие в состав клетки
Органические и неорганические вещества входящие в состав клетки Химический элемент цинк
Химический элемент цинк Витамины - кофакторы ферментативных реакций. Классификация витаминов. Гиповитаминозы. Гипервитаминозы. Антивитамины
Витамины - кофакторы ферментативных реакций. Классификация витаминов. Гиповитаминозы. Гипервитаминозы. Антивитамины Изменчивость. Виды
Изменчивость. Виды Курс зоологии (тип моллюски)
Курс зоологии (тип моллюски) Презентация Классы и семейства цветковых растений
Презентация Классы и семейства цветковых растений Хімічний склад клітини
Хімічний склад клітини Наследственная изменчивость. Мутации
Наследственная изменчивость. Мутации Презентация по биологии для учащихся 7 класса по теме Класс Пресмыкающиеся
Презентация по биологии для учащихся 7 класса по теме Класс Пресмыкающиеся Червона книга України
Червона книга України Тяжёлые металлы. Главный источник поступления тяжелых металлов
Тяжёлые металлы. Главный источник поступления тяжелых металлов Презентация Членистоногие
Презентация Членистоногие V.A. Motsart hayoti va ijodi
V.A. Motsart hayoti va ijodi Патогенные микроорганизмы
Патогенные микроорганизмы Органы чувств. 5 класс
Органы чувств. 5 класс Условия роста и развития растений
Условия роста и развития растений Class muscle
Class muscle Место человека в живой природе
Место человека в живой природе Как появился человек на Земле
Как появился человек на Земле Растения Донского края
Растения Донского края Введение в биохимию. Строение и функции белков
Введение в биохимию. Строение и функции белков