- Биоэнергетика бактериальной клетки

Содержание
- 3. Утилизация углеводов 1. Расщепление экзоферментами до моно и дисахаров (гексоз), которые проникают в клетку активным транспортом
- 4. СН3 – СО-СООН Пировиноградная кислота является исходным соединением в процессах биосинтеза и распада
- 5. Пути расщепления глюкозы 1. ФДФ ( фруктозо 1,6-дифоставтный путь: Эбдена Мейергофа-Парнаса) 2. Пентозный цикл(окислительный ментозофосфатный путь:
- 6. Глюкоза-6-фосфат Глюкоза вначале фосфорилируется в положение 6 Глюкоза-6-фосфат –метаболически активная форма глюкозы-исходное соединение для всех 3
- 7. ФДФ-путь
- 8. ФДФ-путь Все реакции обратимы за исключением ФОСФОФРУКТОКИНАЗЫ, ГЕКСОКИНАЗЫ и ПИРУВАТКИНАЗЫ Баланс: 2 пирувата 2 АТФ 2
- 9. Пентозный цикл
- 10. Пентозный цикл Итог: подготовка исходных веществ для биосинтетических процессов. )пентозофосфаты= предшественники нуклеотидов) 1СО2 2 НАДФ Н
- 11. КДФГ
- 12. КДФГ путь
- 13. КДФГ-путь Принципиальное значение имеет при использовании глюконовой кислоты в качестве источника С и энергии. Баланс: 1
- 14. Получение АТФ АТФ-универсальный переносчик химической энергии между реакциями дающими и использующими энергию Энергией богаты пиросфотные связи
- 15. Получение АТФ Регенерация АТФ осуществляется за счет 3 процессов: 1. Фосфорилирование на уровне субстрата ( при
- 16. Окислительное фосфорилирование При окислительном фосфорилировании происходит одновременный перенос 2 электронов, при этом от субстрата отщепляются 2
- 17. Процесс дегидрирования Н+ переносится дегидрогеназами на коферменты НАД и НАДФ НАД участвует в процессах дыхания и
- 18. Распад гексоз Ключевым веществом распада гексоз является пируват Пируват при участии ко-факторов и дегидрогеназы превращается в
- 20. Распад гексоз (ЦТК) Атомы Н+ при помощи изоцитрат, альфа-кетоклюторат и малат-дегидрогеназ поступают в АТФ-генерирующую систему дыхательной
- 21. ЦТК ЦТК выполняет не только функцию, связанную с получением энергии, но ипоставляет исходные соединения для синтеза
- 22. Анаплерические реакции Восполнение потери промежуточных продуктов происходит при помощи АНАПЛЕРИЧЕСКИХ реакций, одной из функций которых, является
- 23. Анаплерические реакции При осте на среде с глюкозой , она используется для синтеза всех компонентов клетки.
- 24. Цикл Кребса -Корнберга Рост на среде с АЦЕТАТОМ возможен благодаря глиоксилатному циклу или циклу Кребса-Корнберга. ОН
- 25. Цикл Кребса-Корнберга
- 26. Цикл Кребса-Корнберга
- 27. Цикл дикарбоновых кислот Когда рост бактерий протекает на среде с глиоксилатом, его окисление происходит по циклу
- 29. Брожение В анаэробных условиях при сбраживании углеводов используется энергия , полученная в результате превращения триозофосфата в
- 30. ФДФ-путь
- 31. Брожение При брожении отщепившийся от триозофосфата (Н+) переносится на НАД Одновременно происходит фосфорилирование на уровне субстрата
- 32. Реакция Стикленда Сопряженное сбраживание 2 аминокислот-реакция Стикленда Пептолитические клостридии гидролизируют белки с образованием аминокислот, которые сбраживаются
- 33. Реакция Стикленда Аланин сбраживается с глицином Аланин – днор (Н+) Глицин –акцептор (Н+) Донор дезаминируется с
- 34. Реакция Стикленда Аланин + глицин + Н2О 3СР3СООН + 3NН3 + СО2
- 35. СПИРТОВОЕ БРОЖЕНИЕ Спиртовым брожением называется процесс расщепления сахара микроорганизмами с образованием этилового спирта и углекислого газа.
- 36. Молочнокислое брожение Молочнокислое брожение- это анаэробное превращение сахара молочнокислыми бактериями с образованием молочной кислоты. По характеру
- 37. Молочнокислое брожение Процесс превращения глюкозы до пировиноградной кислоты у гомоферментатавных молочнокислых бактерий протекает Г-6-Ф пути. Далее
- 38. Гетероферментативное молочнокислое брожение Отсутствие фермента адьдолазы меняет начальный путь превращения глюкозы. После фосфорилирования гексоза окисляется (отщепляется
- 39. Маслянокислое брожение Маслянокислое брожение - это процесс превращения сахара маслянокислыми бактериями в анаэробных условиях с образованием
- 40. Маслянокислое брожение Пировиноградная кислота декарбоксилируется с образованием углекислого газа и уксусного альдегида. Далее под действием фермента
- 41. Муравьинокислое брожение СН3-СО-СООН НСООН Ас КоА Н2 СО2 Происходит образование кислот и газов, которые выявляются в
- 42. Муравьинокислое брожение По второму варианту происходит конденсация 2 молекул пирувата с последующими двумя декарбоксилированием с образование
- 43. Регуляция углеводного обмена Эффект Пастера Торможение брожения дыханием Связан с конкуренцией между системами дыхания и брожения
- 44. Регуляция углеводного обмена Эффект Пастера Эффект Пастера обеспечивает экономию для клетки углеводов, так как дыхание энергетически
- 45. Катаболическая репрессия Приспособление клетки к использованию в первую очередь наиболее легко усвояемых источников энергии При присутствии
- 46. Строение лактозного оперона
- 47. Функционирование лактозного оперона
- 48. Катаболическая репрессия К.Р. Связана с фосфотрансферной системой ФТС обеспечивает накопление в клетке глюкозы, маннозы, манитола в
- 49. ФТС Богатая энергией фосфатная группа передается от ФЕП на белок-переносчик H – Pr, превращая его в
- 50. ФТС
- 51. Катаболическая репрессия При низкой концентрации глюкозы в среде происходит накопление ФЕП, который способен активировать аденилатциклазу, повышая
- 53. Скачать презентацию
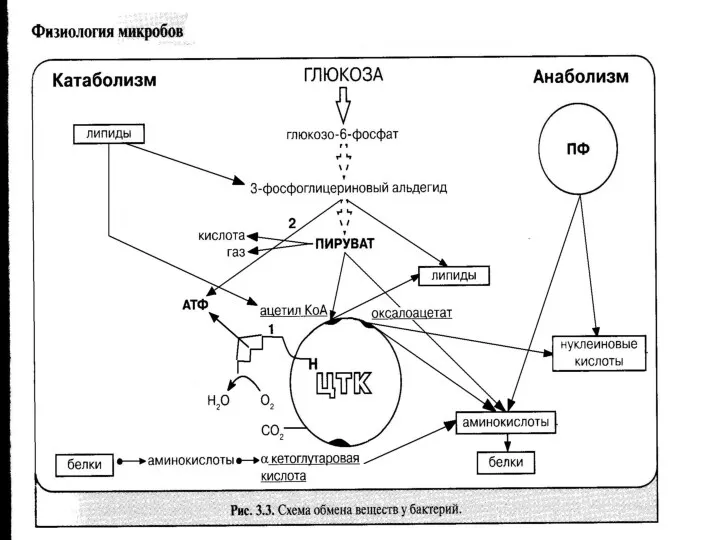

Утилизация углеводов
1. Расщепление экзоферментами до моно и дисахаров (гексоз), которые проникают
Утилизация углеводов
1. Расщепление экзоферментами до моно и дисахаров (гексоз), которые проникают

СН3 – СО-СООН
Пировиноградная кислота является исходным соединением в процессах биосинтеза и
СН3 – СО-СООН
Пировиноградная кислота является исходным соединением в процессах биосинтеза и

Пути расщепления глюкозы
1. ФДФ ( фруктозо 1,6-дифоставтный путь: Эбдена Мейергофа-Парнаса)
2. Пентозный
Пути расщепления глюкозы
1. ФДФ ( фруктозо 1,6-дифоставтный путь: Эбдена Мейергофа-Парнаса)
2. Пентозный

Глюкоза-6-фосфат
Глюкоза вначале фосфорилируется в положение 6
Глюкоза-6-фосфат –метаболически активная форма глюкозы-исходное соединение
Глюкоза-6-фосфат
Глюкоза вначале фосфорилируется в положение 6
Глюкоза-6-фосфат –метаболически активная форма глюкозы-исходное соединение
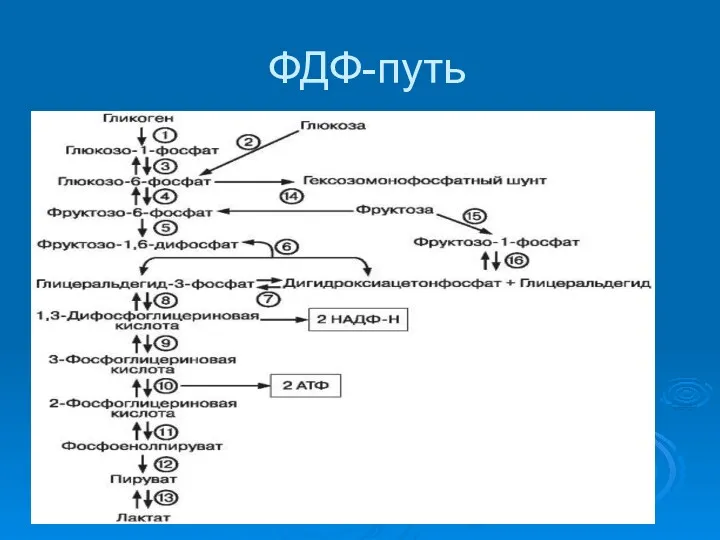
ФДФ-путь
ФДФ-путь

ФДФ-путь
Все реакции обратимы за исключением ФОСФОФРУКТОКИНАЗЫ, ГЕКСОКИНАЗЫ и ПИРУВАТКИНАЗЫ
Баланс: 2 пирувата
ФДФ-путь
Все реакции обратимы за исключением ФОСФОФРУКТОКИНАЗЫ, ГЕКСОКИНАЗЫ и ПИРУВАТКИНАЗЫ
Баланс: 2 пирувата
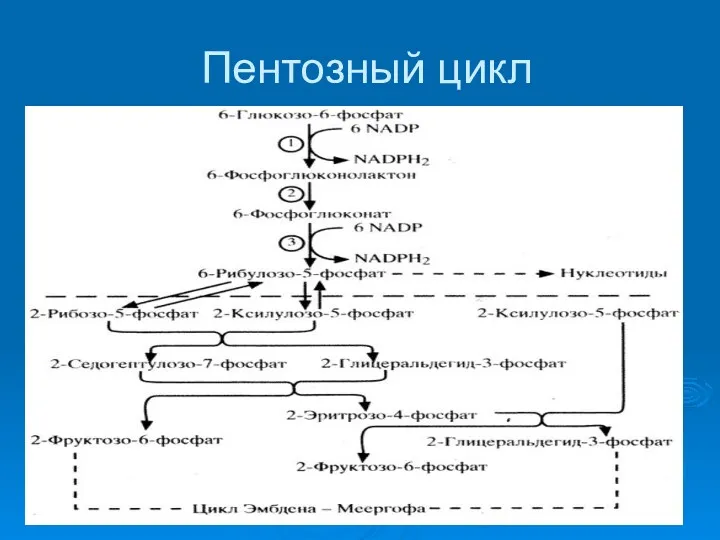
Пентозный цикл
Пентозный цикл

Пентозный цикл
Итог: подготовка исходных веществ для биосинтетических процессов. )пентозофосфаты= предшественники нуклеотидов)
Пентозный цикл
Итог: подготовка исходных веществ для биосинтетических процессов. )пентозофосфаты= предшественники нуклеотидов)
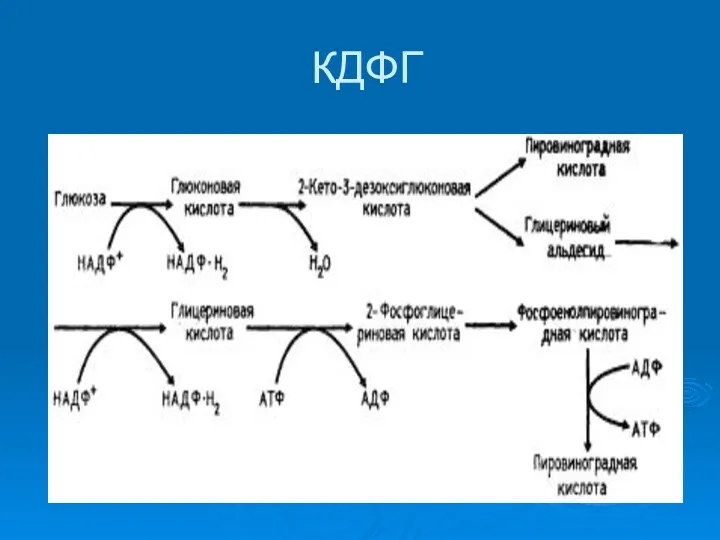
КДФГ
КДФГ

КДФГ путь
КДФГ путь

КДФГ-путь
Принципиальное значение имеет при использовании глюконовой кислоты в качестве источника С
КДФГ-путь
Принципиальное значение имеет при использовании глюконовой кислоты в качестве источника С

Получение АТФ
АТФ-универсальный переносчик химической энергии между реакциями дающими и использующими энергию
Энергией
Получение АТФ
АТФ-универсальный переносчик химической энергии между реакциями дающими и использующими энергию
Энергией

Получение АТФ
Регенерация АТФ осуществляется за счет 3 процессов:
1. Фосфорилирование на уровне
Получение АТФ
Регенерация АТФ осуществляется за счет 3 процессов:
1. Фосфорилирование на уровне

Окислительное фосфорилирование
При окислительном фосфорилировании происходит одновременный перенос 2 электронов, при этом
Окислительное фосфорилирование
При окислительном фосфорилировании происходит одновременный перенос 2 электронов, при этом

Процесс дегидрирования
Н+ переносится дегидрогеназами на коферменты НАД и НАДФ
НАД участвует в
Процесс дегидрирования
Н+ переносится дегидрогеназами на коферменты НАД и НАДФ
НАД участвует в

Распад гексоз
Ключевым веществом распада гексоз является пируват
Пируват при участии ко-факторов и
Распад гексоз
Ключевым веществом распада гексоз является пируват
Пируват при участии ко-факторов и
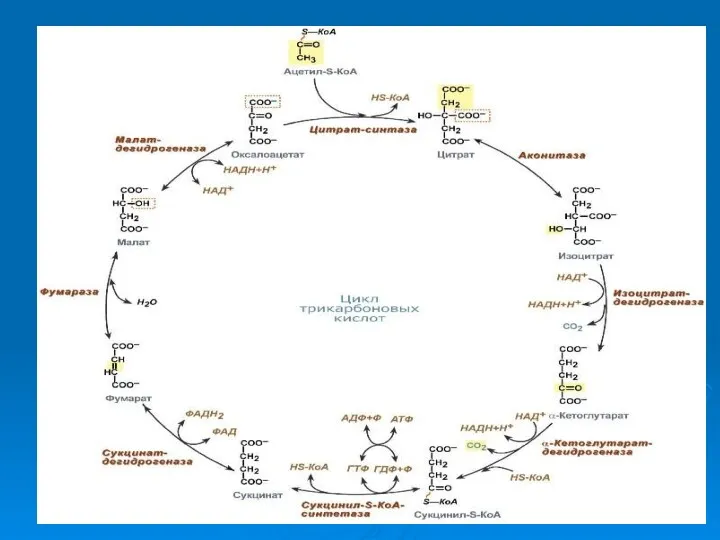

Распад гексоз (ЦТК)
Атомы Н+ при помощи изоцитрат, альфа-кетоклюторат и малат-дегидрогеназ поступают
Распад гексоз (ЦТК)
Атомы Н+ при помощи изоцитрат, альфа-кетоклюторат и малат-дегидрогеназ поступают

ЦТК
ЦТК выполняет не только функцию, связанную с получением энергии, но ипоставляет
ЦТК
ЦТК выполняет не только функцию, связанную с получением энергии, но ипоставляет

Анаплерические реакции
Восполнение потери промежуточных продуктов происходит при помощи АНАПЛЕРИЧЕСКИХ реакций, одной
Анаплерические реакции
Восполнение потери промежуточных продуктов происходит при помощи АНАПЛЕРИЧЕСКИХ реакций, одной

Анаплерические реакции
При осте на среде с глюкозой , она используется для
Анаплерические реакции
При осте на среде с глюкозой , она используется для

Цикл Кребса -Корнберга
Рост на среде с АЦЕТАТОМ возможен благодаря глиоксилатному циклу
Цикл Кребса -Корнберга
Рост на среде с АЦЕТАТОМ возможен благодаря глиоксилатному циклу
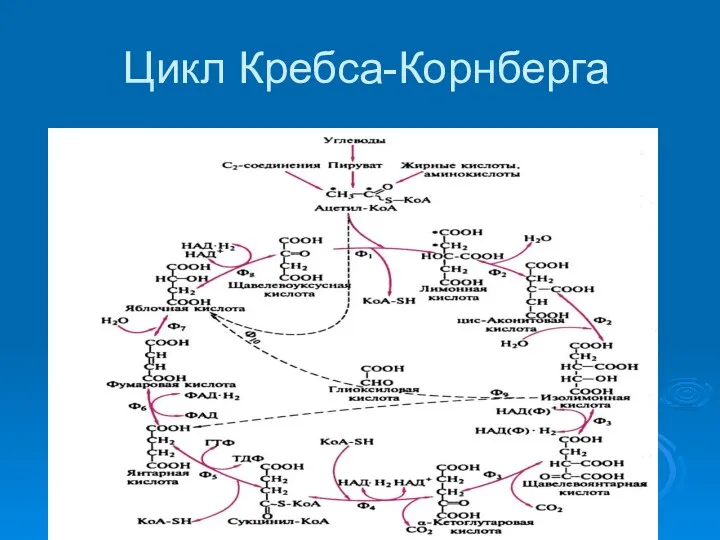
Цикл Кребса-Корнберга
Цикл Кребса-Корнберга
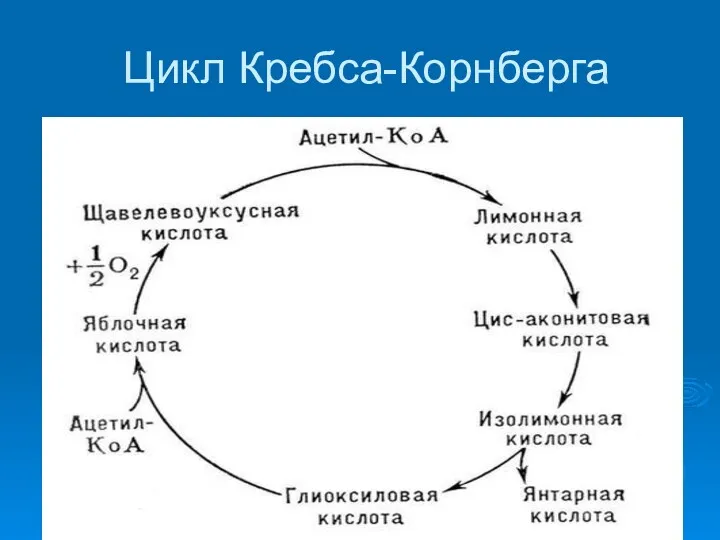
Цикл Кребса-Корнберга
Цикл Кребса-Корнберга

Цикл дикарбоновых кислот
Когда рост бактерий протекает на среде с глиоксилатом, его
Цикл дикарбоновых кислот
Когда рост бактерий протекает на среде с глиоксилатом, его


Брожение
В анаэробных условиях при сбраживании углеводов используется энергия , полученная в
Брожение
В анаэробных условиях при сбраживании углеводов используется энергия , полученная в
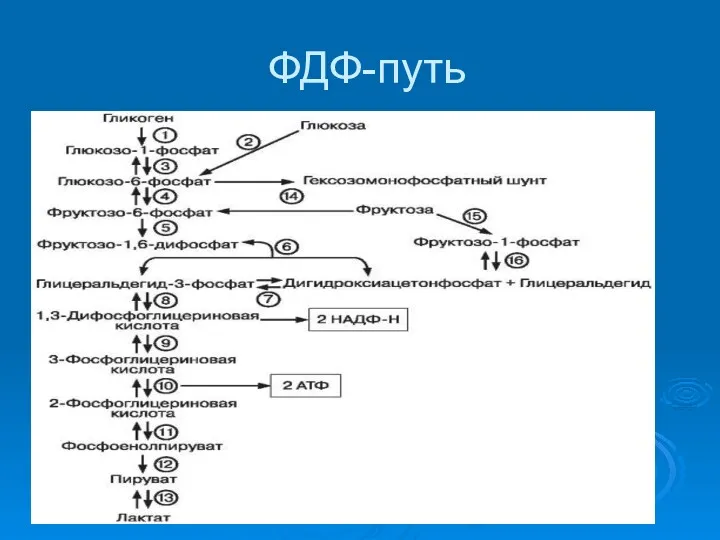
ФДФ-путь
ФДФ-путь

Брожение
При брожении отщепившийся от триозофосфата (Н+) переносится на НАД
Одновременно происходит фосфорилирование
Брожение
При брожении отщепившийся от триозофосфата (Н+) переносится на НАД
Одновременно происходит фосфорилирование

Реакция Стикленда
Сопряженное сбраживание 2 аминокислот-реакция Стикленда
Пептолитические клостридии гидролизируют белки с образованием
Реакция Стикленда
Сопряженное сбраживание 2 аминокислот-реакция Стикленда
Пептолитические клостридии гидролизируют белки с образованием

Реакция Стикленда
Аланин сбраживается с глицином
Аланин – днор (Н+)
Глицин –акцептор (Н+)
Донор дезаминируется
Реакция Стикленда
Аланин сбраживается с глицином
Аланин – днор (Н+)
Глицин –акцептор (Н+)
Донор дезаминируется

Реакция Стикленда
Аланин + глицин + Н2О
3СР3СООН + 3NН3 + СО2
Реакция Стикленда
Аланин + глицин + Н2О
3СР3СООН + 3NН3 + СО2

СПИРТОВОЕ БРОЖЕНИЕ
Спиртовым брожением называется процесс расщепления сахара микроорганизмами с образованием этилового
СПИРТОВОЕ БРОЖЕНИЕ
Спиртовым брожением называется процесс расщепления сахара микроорганизмами с образованием этилового

Молочнокислое брожение
Молочнокислое брожение- это анаэробное превращение сахара молочнокислыми бактериями с образованием
Молочнокислое брожение
Молочнокислое брожение- это анаэробное превращение сахара молочнокислыми бактериями с образованием

Молочнокислое брожение
Процесс превращения глюкозы до пировиноградной кислоты у гомоферментатавных молочнокислых бактерий
Молочнокислое брожение
Процесс превращения глюкозы до пировиноградной кислоты у гомоферментатавных молочнокислых бактерий

Гетероферментативное молочнокислое брожение
Отсутствие фермента адьдолазы меняет начальный путь превращения глюкозы. После
Гетероферментативное молочнокислое брожение
Отсутствие фермента адьдолазы меняет начальный путь превращения глюкозы. После

Маслянокислое брожение
Маслянокислое брожение - это процесс превращения сахара маслянокислыми бактериями в
Маслянокислое брожение
Маслянокислое брожение - это процесс превращения сахара маслянокислыми бактериями в

Маслянокислое брожение
Пировиноградная кислота декарбоксилируется с образованием углекислого газа и уксусного альдегида.
Маслянокислое брожение
Пировиноградная кислота декарбоксилируется с образованием углекислого газа и уксусного альдегида.
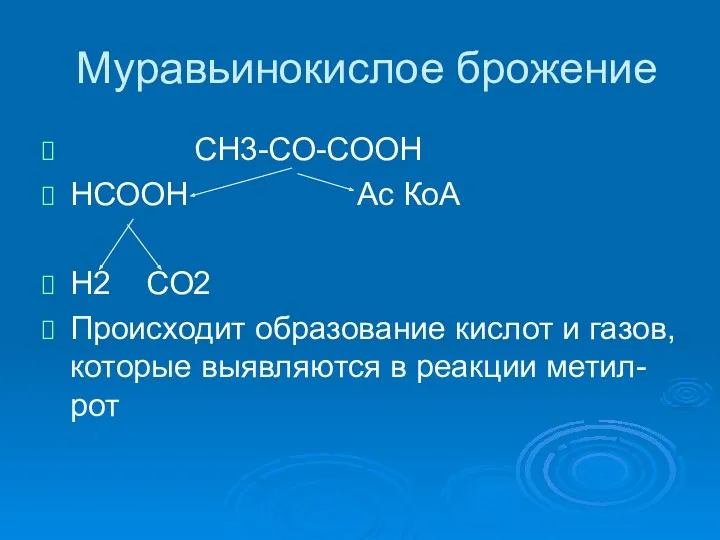
Муравьинокислое брожение
СН3-СО-СООН
НСООН Ас КоА
Н2 СО2
Происходит образование кислот и газов, которые
Муравьинокислое брожение
СН3-СО-СООН
НСООН Ас КоА
Н2 СО2
Происходит образование кислот и газов, которые

Муравьинокислое брожение
По второму варианту происходит конденсация 2 молекул пирувата с последующими
Муравьинокислое брожение
По второму варианту происходит конденсация 2 молекул пирувата с последующими

Регуляция углеводного обмена
Эффект Пастера
Торможение брожения дыханием
Связан с конкуренцией между системами дыхания
Регуляция углеводного обмена
Эффект Пастера
Торможение брожения дыханием
Связан с конкуренцией между системами дыхания

Регуляция углеводного обмена
Эффект Пастера
Эффект Пастера обеспечивает экономию для клетки углеводов, так
Регуляция углеводного обмена
Эффект Пастера
Эффект Пастера обеспечивает экономию для клетки углеводов, так

Катаболическая репрессия
Приспособление клетки к использованию в первую очередь наиболее легко усвояемых
Катаболическая репрессия
Приспособление клетки к использованию в первую очередь наиболее легко усвояемых
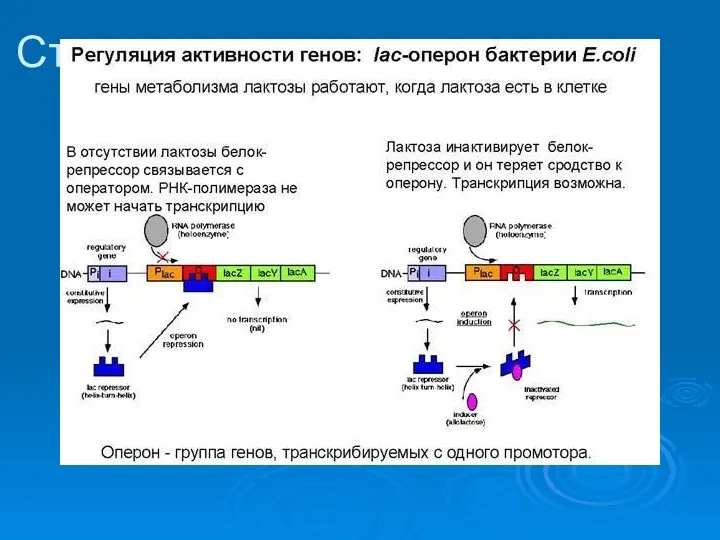
Строение лактозного оперона
Строение лактозного оперона
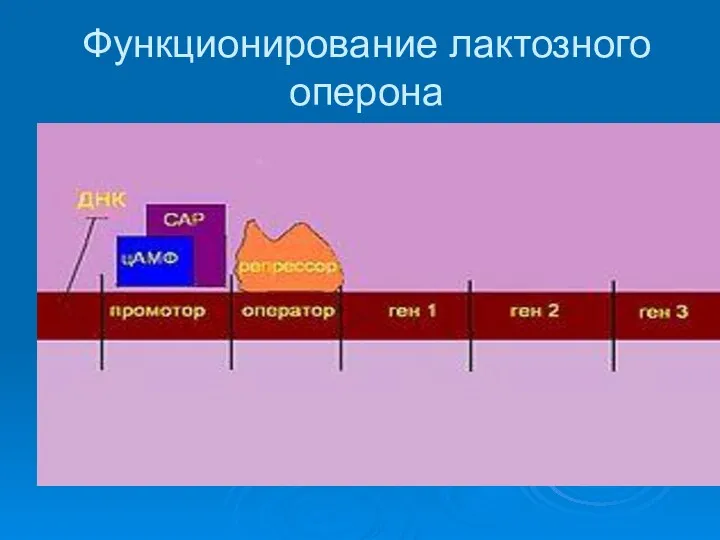
Функционирование лактозного оперона
Функционирование лактозного оперона

Катаболическая репрессия
К.Р. Связана с фосфотрансферной системой
ФТС обеспечивает накопление в клетке глюкозы,
Катаболическая репрессия
К.Р. Связана с фосфотрансферной системой
ФТС обеспечивает накопление в клетке глюкозы,

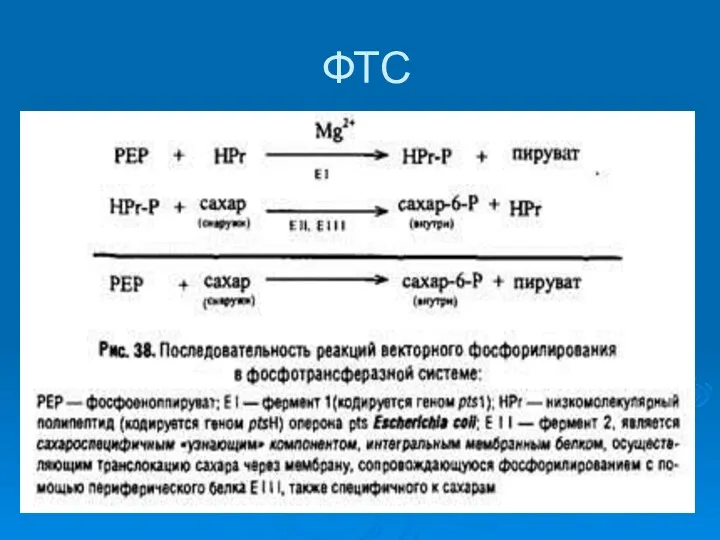
ФТС
Богатая энергией фосфатная группа передается от ФЕП на белок-переносчик H –
ФТС
Богатая энергией фосфатная группа передается от ФЕП на белок-переносчик H –
ФТС
ФТС

Катаболическая репрессия
При низкой концентрации глюкозы в среде происходит накопление ФЕП, который
Катаболическая репрессия
При низкой концентрации глюкозы в среде происходит накопление ФЕП, который
 Елді мекендерді канализациялау жобаларын санитарлық сараптау сатылары. Берілген құжаттардың жеткіліктілігін анықтау
Елді мекендерді канализациялау жобаларын санитарлық сараптау сатылары. Берілген құжаттардың жеткіліктілігін анықтау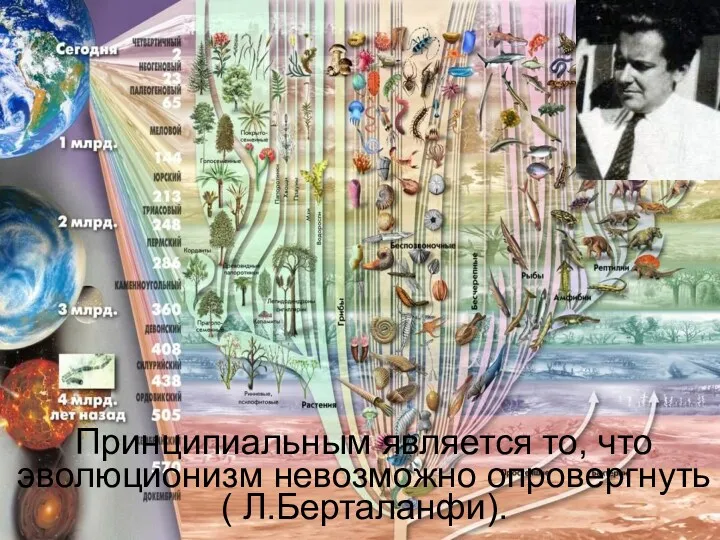 Генетические основы эволюции
Генетические основы эволюции Основы микробиологии и иммунологии
Основы микробиологии и иммунологии Эволюция животного мира
Эволюция животного мира Учение о тканях. Эпителиальные ткани
Учение о тканях. Эпителиальные ткани Западно-алтайский национальный природный заповедник
Западно-алтайский национальный природный заповедник Нарушения кислотно-щелочного равновесия
Нарушения кислотно-щелочного равновесия Твій город на підвіконні. Весняні роботи в місті та в селі (урок № 73 - 74)
Твій город на підвіконні. Весняні роботи в місті та в селі (урок № 73 - 74) Трансгенні організми. (Лекція 7)
Трансгенні організми. (Лекція 7)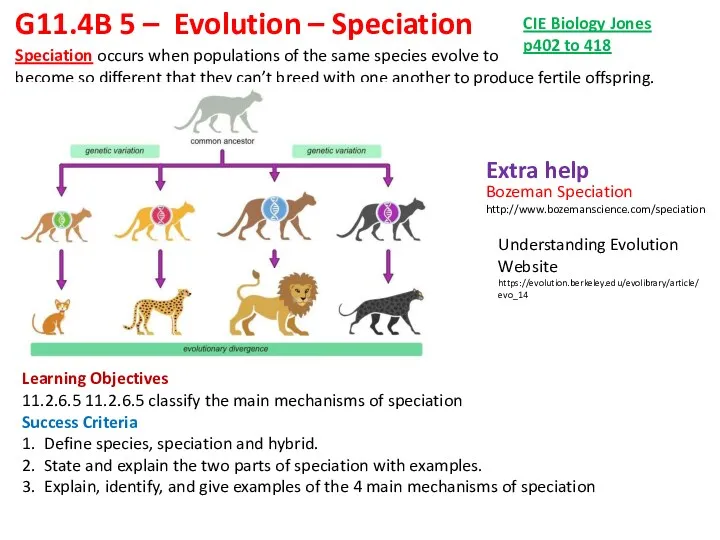 Evolution – Speciation
Evolution – Speciation Морфокинезиологический анализ пояса верхних конечностей
Морфокинезиологический анализ пояса верхних конечностей Роль спинного и продолговатого мозга в регуляции двигательных функций. Спинальный шок
Роль спинного и продолговатого мозга в регуляции двигательных функций. Спинальный шок Основные особенности растительной клетки
Основные особенности растительной клетки Влияние загрязнения окружающей среды на здоровье
Влияние загрязнения окружающей среды на здоровье Нервная система
Нервная система Отряд хищные
Отряд хищные История, легенды и мифы о бабочках
История, легенды и мифы о бабочках Анатомо-физиологические особенности сердечно- сосудистой системы. Анатомия сердца
Анатомо-физиологические особенности сердечно- сосудистой системы. Анатомия сердца Види умовних рефлексів, їхнє гальмування
Види умовних рефлексів, їхнє гальмування Репчатый лук
Репчатый лук Моллюски. Общая характеристика моллюсков
Моллюски. Общая характеристика моллюсков Большой и малый круг кровообращения
Большой и малый круг кровообращения Обмен углеводов
Обмен углеводов Сущность мутационной наследственности. Виды мутаций и факторы, способствующие их возникновению
Сущность мутационной наследственности. Виды мутаций и факторы, способствующие их возникновению Кости пояса верхней конечности
Кости пояса верхней конечности Тренажёр Плоды
Тренажёр Плоды Многообразие земноводных
Многообразие земноводных Изучение влияния табачного дыма на живые организмы
Изучение влияния табачного дыма на живые организмы