- Биоэнергетика. Тканевое дыхание и окислительное фосфорилирование

Содержание
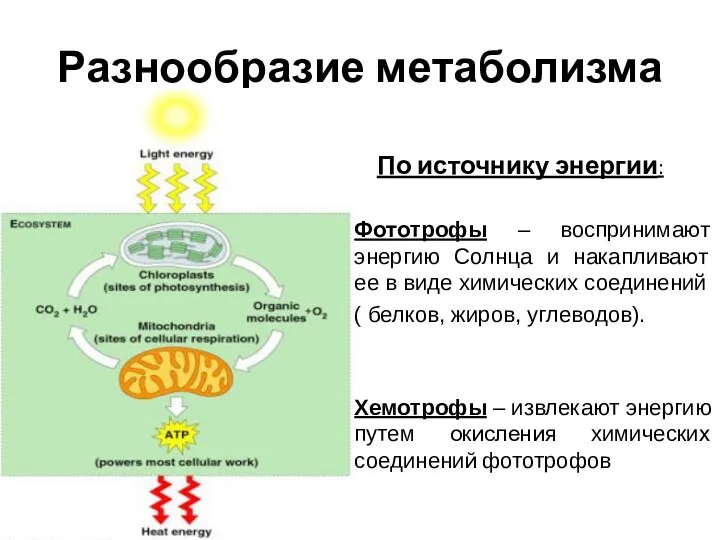
- 2. Разнообразие метаболизма По источнику энергии: Фототрофы – воспринимают энергию Солнца и накапливают ее в виде химических
- 3. 2-ой закон Термодинамики при необратимых процессах энтропия системы и окружающей среды увеличивается системы стремятся к неупорядоченности
- 4. Энтропия Энтропия растет! мера беспорядка (случайности) в системе (S)
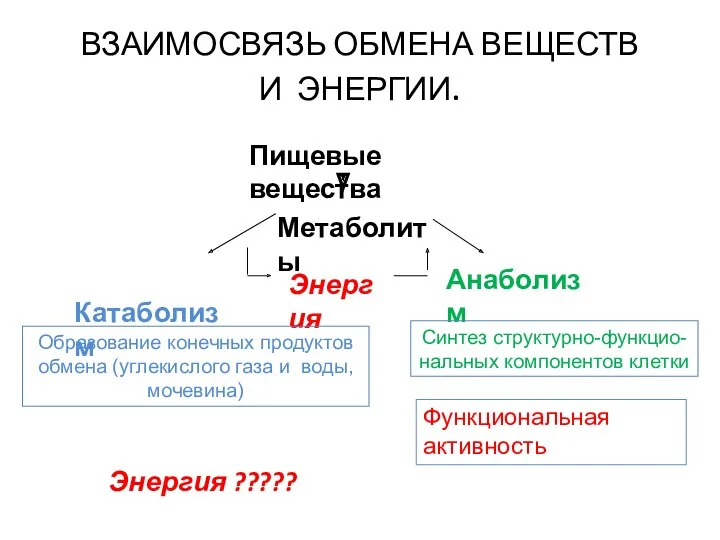
- 5. ВЗАИМОСВЯЗЬ ОБМЕНА ВЕЩЕСТВ И ЭНЕРГИИ. Пищевые вещества Метаболиты Катаболизм Анаболизм Энергия Образование конечных продуктов обмена (углекислого
- 6. АТФ Fritz Albert Lipmann (1899-1986) “ATP is energy-transfering molecule in the cell” (1941) Karl Lohmann (1898-1978)
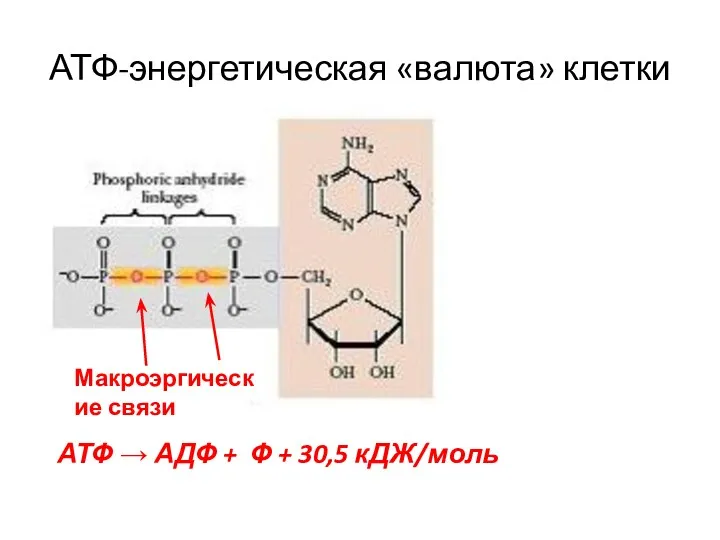
- 7. АТФ-энергетическая «валюта» клетки Макроэргические связи АТФ → АДФ + Ф + 30,5 кДЖ/моль
- 8. Взрослый человек потребляет примерно 60 кг АТФ в день. Во взрослом организме содержится около 50 г
- 9. АТФ образуется в клетке двумя путями: - окислительное фосфорилирование: синтез АТФ с использованием энергии выделяемой при
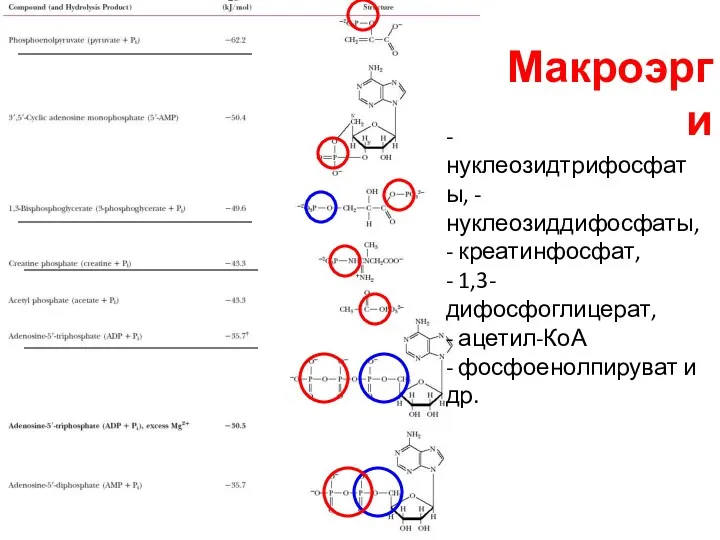
- 10. Макроэрги - нуклеозидтрифосфаты, - нуклеозиддифосфаты, - креатинфосфат, - 1,3-дифосфоглицерат, - ацетил-КоА - фосфоенолпируват и др.
- 11. Синтез АТФ в организме сопряжен с реакцией образования воды Этот процесс происходит многостадийно с помощью многих
- 12. Общий путь катаболизма
- 13. Дыхательная цепь (цепь переноса электронов, ЦПЭ) – последовательность оксидоредуктаз,транспортирующих Н+ и е- от S на О2.
- 14. Ферменты ДЦ локализованы на внутренней мембране митохондриях
- 15. Компоненты ДЦ Пиридиновые ДГ
- 16. Флавиновые ДГ
- 17. Цитохромы b, c1 , c , a, a3 Цит. (Fe3+ ) + е → Цит. (
- 18. Выделение энергии происходит постепенно, порциями. Дыхательная цепь Цепь переноса электронов (ЦПЭ)
- 19. ДЦ состоит из 5 ферментных комплесов: 1) НАДН - КоQ-редуктаза ( НАДН-дегилрогеназа); 2) сукцинат – КоQ-редуктаза
- 20. Принцип работы дыхательной цепи: разделение потоков протонов и электронов, поступающих из матрикса. Электроны передаются на конечный
- 21. Дыхательные ферменты расположены в порядке ↑ величины редокс - потенциала Место того или иного переносчика в
- 22. Редокс-потенциал (Ео′) численно равен электрондвижущей силе в Вольтах, возникающей между растворами окислителя и восстановителя (концентрации 1М,
- 23. Хемиоосмотическая теория окислительного фосфорилирования (Нобелевская премия 1978 г.).
- 24. Основные постулаты хемиоосмотической теории: внутренняя митохондриальная мембрана (ВММ) непроницаема для ионов за счет энергии транспорта электронов
- 25. Связь между транспортом электронов в ДЦ и синтезом АТФ: протонная АТФ -синтаза Дальнейшие исследования (Дж. Уокер,
- 26. Энергетический эффект биологического окисления Пункты сопряжения тканевого дыхания и окислительного фосфорилирования. Синтез 1 моль АТФ ≈
- 27. Дыхательный контроль Регулируется скорость работы дыхательной цепи энергетическим зарядом клетки, т. е. соотношением АТФ/АДФ. Эффективность окислительного
- 28. Ингибиторы ДЦ
- 29. 2,4- ДНФ
- 30. Субстраты ДЦ
- 32. Скачать презентацию
Разнообразие метаболизма
По источнику энергии:
Фототрофы – воспринимают энергию Солнца и накапливают ее в виде
Разнообразие метаболизма
По источнику энергии:
Фототрофы – воспринимают энергию Солнца и накапливают ее в виде
2-ой закон Термодинамики
при необратимых процессах энтропия системы и окружающей среды увеличивается
системы стремятся к
2-ой закон Термодинамики
при необратимых процессах энтропия системы и окружающей среды увеличивается
системы стремятся к

Энтропия
Энтропия
растет!
мера беспорядка (случайности) в системе (S)
Энтропия
Энтропия
растет!
мера беспорядка (случайности) в системе (S)

ВЗАИМОСВЯЗЬ ОБМЕНА ВЕЩЕСТВ
И ЭНЕРГИИ.
Пищевые вещества
Метаболиты
Катаболизм
Анаболизм
Энергия
Образование конечных продуктов обмена
ВЗАИМОСВЯЗЬ ОБМЕНА ВЕЩЕСТВ
И ЭНЕРГИИ.
Пищевые вещества
Метаболиты
Катаболизм
Анаболизм
Энергия
Образование конечных продуктов обмена
АТФ
Fritz Albert Lipmann (1899-1986)
“ATP is energy-transfering molecule in the cell” (1941)
Karl Lohmann
АТФ
Fritz Albert Lipmann (1899-1986)
“ATP is energy-transfering molecule in the cell” (1941)
Karl Lohmann

АТФ-энергетическая «валюта» клетки
Макроэргические связи
АТФ → АДФ + Ф + 30,5 кДЖ/моль
АТФ-энергетическая «валюта» клетки
Макроэргические связи
АТФ → АДФ + Ф + 30,5 кДЖ/моль
Взрослый человек потребляет примерно 60 кг АТФ в день.
Во взрослом организме содержится
Взрослый человек потребляет примерно 60 кг АТФ в день.
Во взрослом организме содержится

АТФ образуется в клетке двумя путями: - окислительное фосфорилирование: синтез АТФ с использованием
АТФ образуется в клетке двумя путями: - окислительное фосфорилирование: синтез АТФ с использованием

Макроэрги
- нуклеозидтрифосфаты, - нуклеозиддифосфаты, - креатинфосфат,
- 1,3-дифосфоглицерат,
- ацетил-КоА
- фосфоенолпируват и
Макроэрги
- нуклеозидтрифосфаты, - нуклеозиддифосфаты, - креатинфосфат,
- 1,3-дифосфоглицерат,
- ацетил-КоА
- фосфоенолпируват и
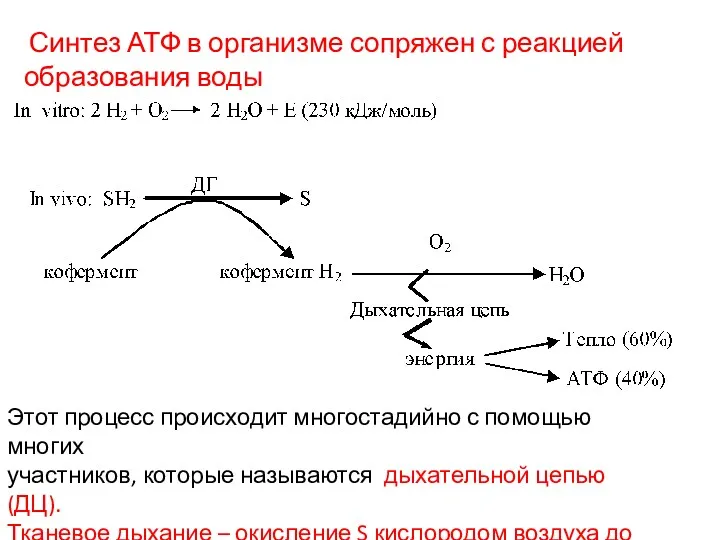
Синтез АТФ в организме сопряжен с реакцией
образования воды
Этот процесс происходит многостадийно
Синтез АТФ в организме сопряжен с реакцией
образования воды
Этот процесс происходит многостадийно
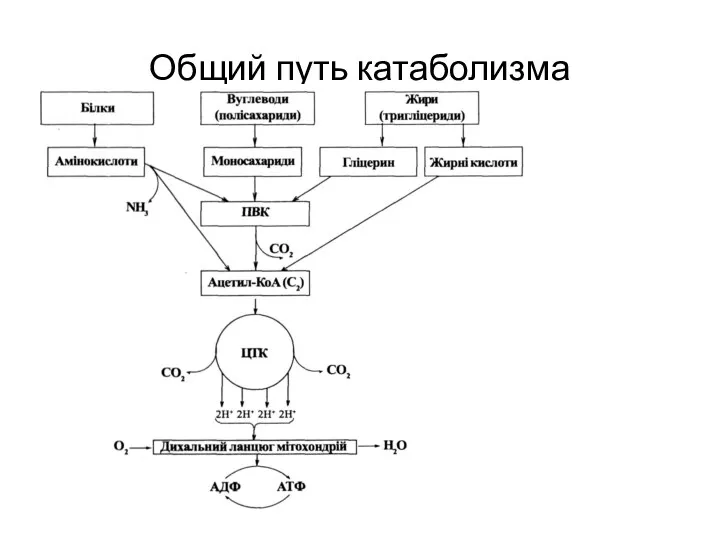
Общий путь катаболизма
Общий путь катаболизма
Дыхательная цепь (цепь переноса электронов, ЦПЭ) – последовательность оксидоредуктаз,транспортирующих Н+ и е- от
Дыхательная цепь (цепь переноса электронов, ЦПЭ) – последовательность оксидоредуктаз,транспортирующих Н+ и е- от

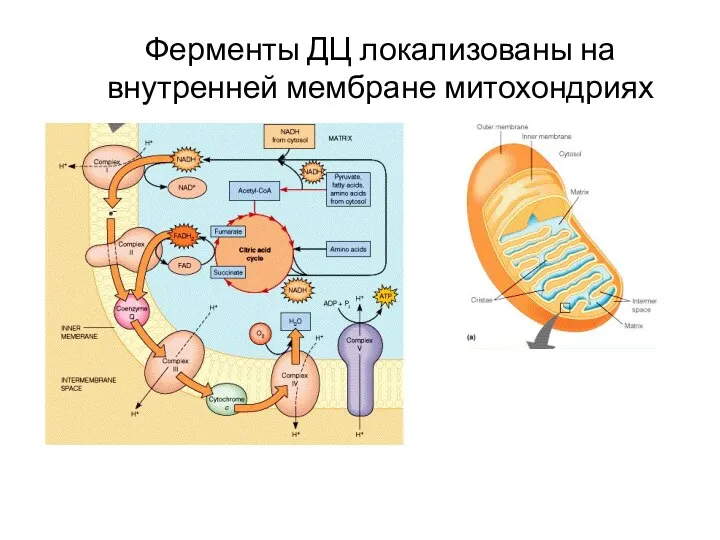
Ферменты ДЦ локализованы на внутренней мембране митохондриях
Ферменты ДЦ локализованы на внутренней мембране митохондриях
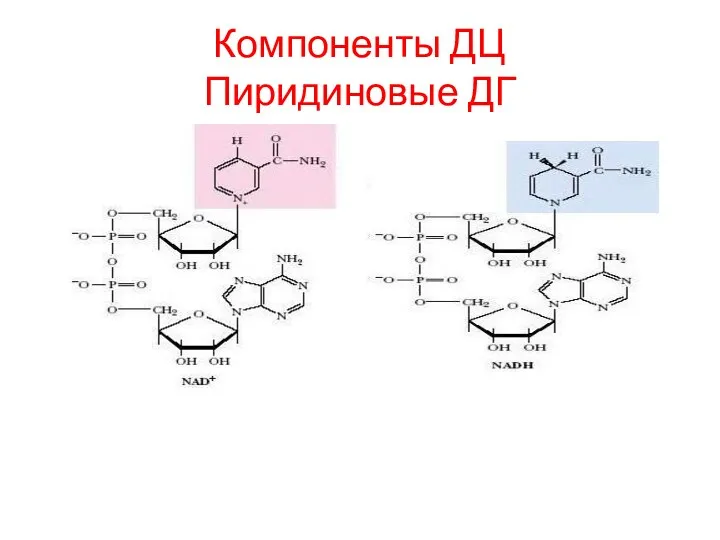
Компоненты ДЦ
Пиридиновые ДГ
Компоненты ДЦ
Пиридиновые ДГ
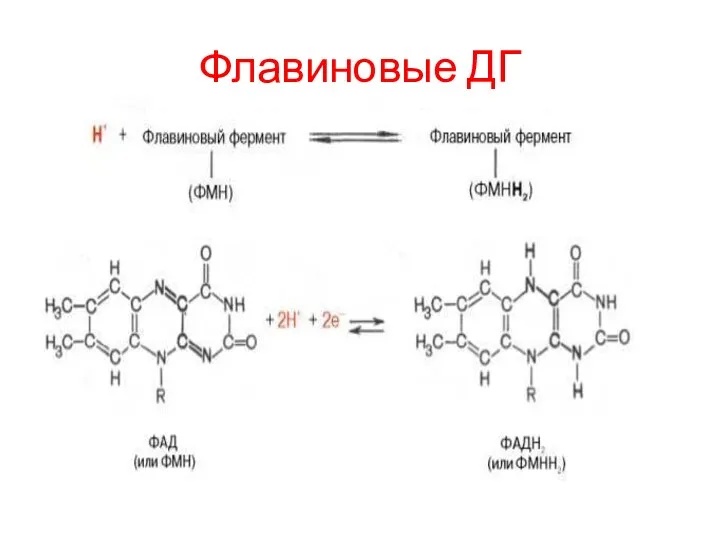
Флавиновые ДГ
Флавиновые ДГ
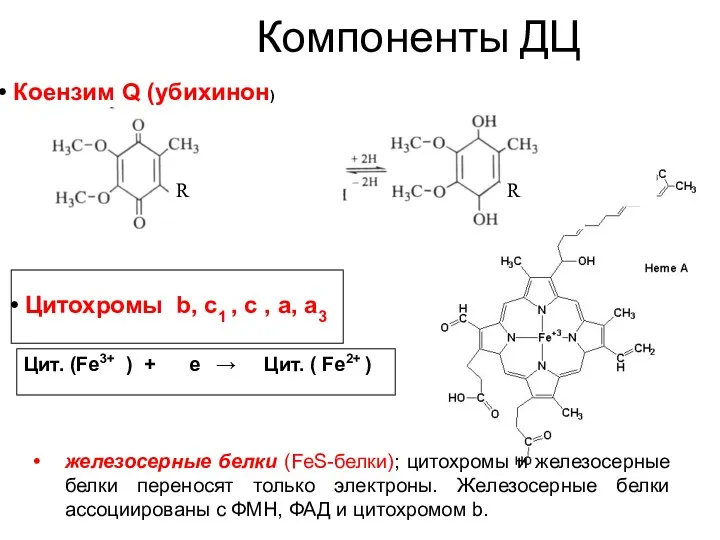
Цитохромы b, c1 , c , a, a3
Цит. (Fe3+ ) + е →
Цит. (Fe3+ ) + е →
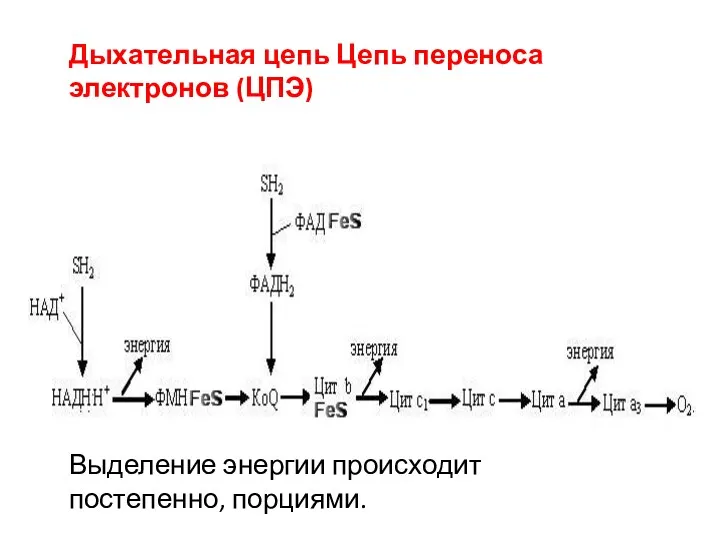
Выделение энергии происходит постепенно, порциями.
Дыхательная цепь Цепь переноса электронов (ЦПЭ)
Выделение энергии происходит постепенно, порциями.
Дыхательная цепь Цепь переноса электронов (ЦПЭ)
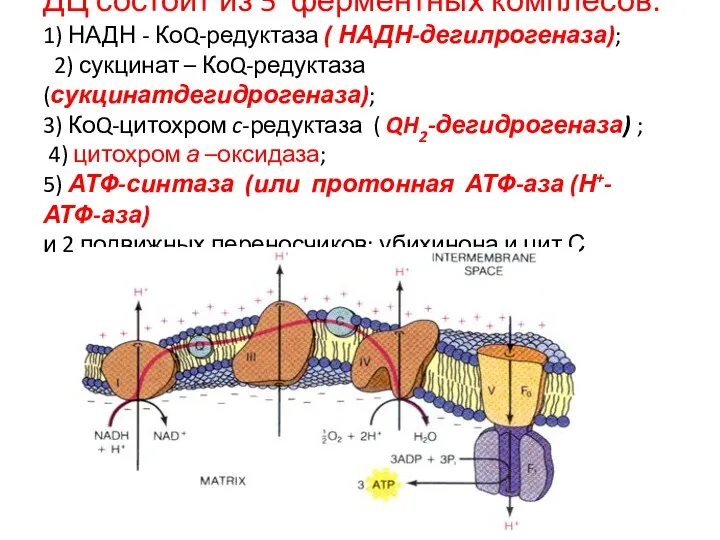
ДЦ состоит из 5 ферментных комплесов:
1) НАДН - КоQ-редуктаза ( НАДН-дегилрогеназа); 2)
ДЦ состоит из 5 ферментных комплесов: 1) НАДН - КоQ-редуктаза ( НАДН-дегилрогеназа); 2)
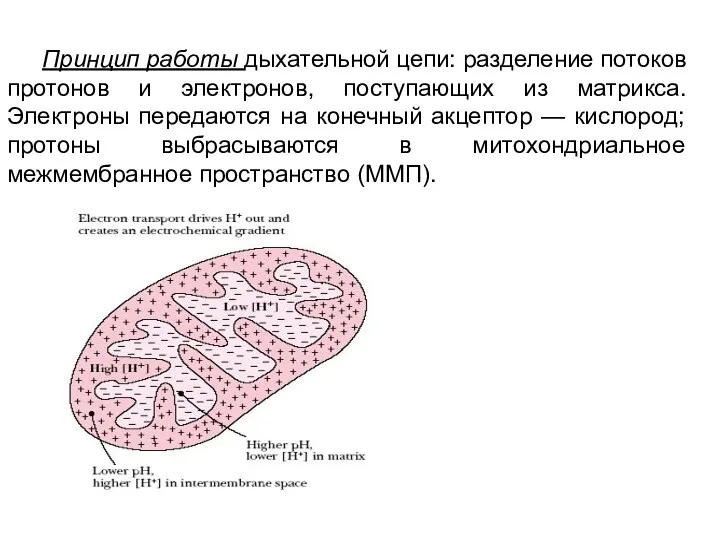
Принцип работы дыхательной цепи: разделение потоков протонов и электронов, поступающих из матрикса. Электроны
Принцип работы дыхательной цепи: разделение потоков протонов и электронов, поступающих из матрикса. Электроны
Дыхательные ферменты расположены в порядке
↑ величины редокс - потенциала
Место того или иного
Дыхательные ферменты расположены в порядке
↑ величины редокс - потенциала
Место того или иного

Редокс-потенциал (Ео′)
численно равен электрондвижущей силе в Вольтах, возникающей между растворами окислителя и
Редокс-потенциал (Ео′)
численно равен электрондвижущей силе в Вольтах, возникающей между растворами окислителя и

Хемиоосмотическая теория окислительного фосфорилирования (Нобелевская премия 1978 г.).
Хемиоосмотическая теория окислительного фосфорилирования (Нобелевская премия 1978 г.).

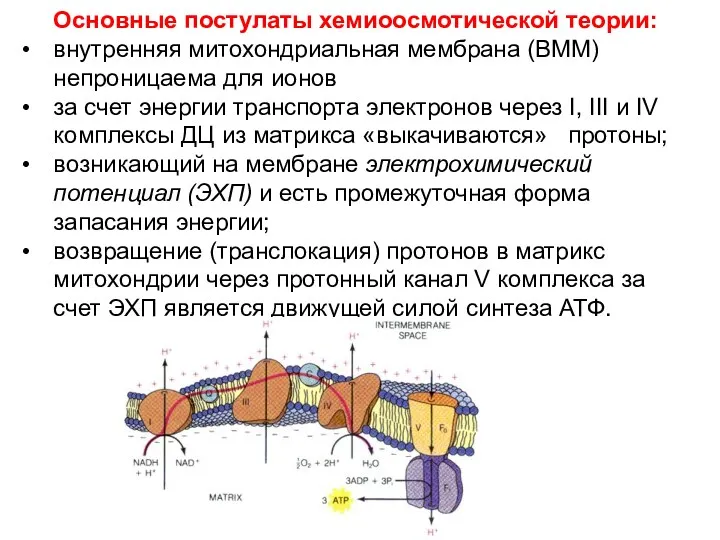
Основные постулаты хемиоосмотической теории:
внутренняя митохондриальная мембрана (ВММ) непроницаема для ионов
за счет энергии транспорта
Основные постулаты хемиоосмотической теории:
внутренняя митохондриальная мембрана (ВММ) непроницаема для ионов
за счет энергии транспорта
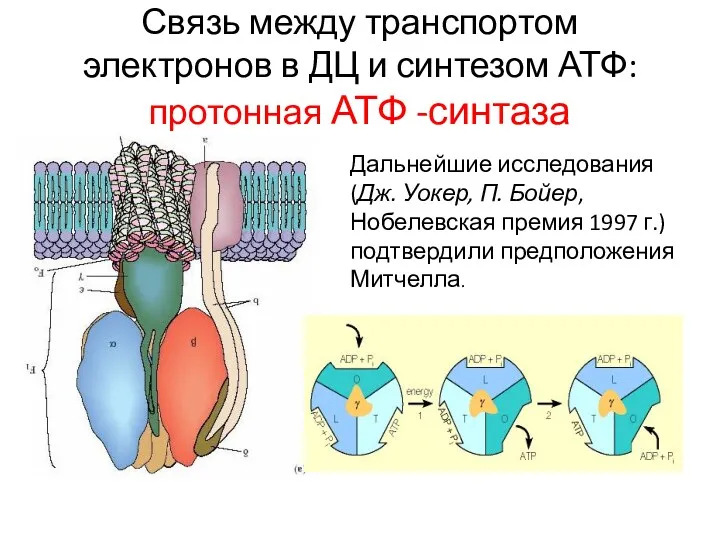
Связь между транспортом электронов в ДЦ и синтезом АТФ: протонная АТФ -синтаза
Дальнейшие исследования
Связь между транспортом электронов в ДЦ и синтезом АТФ: протонная АТФ -синтаза
Дальнейшие исследования
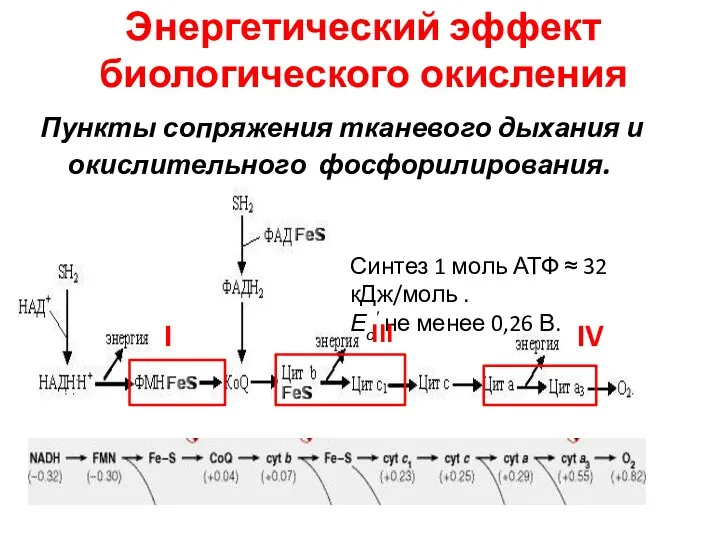
Энергетический эффект биологического окисления
Пункты сопряжения тканевого дыхания и окислительного фосфорилирования.
Синтез 1 моль АТФ
Энергетический эффект биологического окисления
Пункты сопряжения тканевого дыхания и окислительного фосфорилирования.
Синтез 1 моль АТФ
Дыхательный контроль
Регулируется скорость работы дыхательной цепи энергетическим зарядом клетки, т. е. соотношением АТФ/АДФ.
Дыхательный контроль Регулируется скорость работы дыхательной цепи энергетическим зарядом клетки, т. е. соотношением АТФ/АДФ.

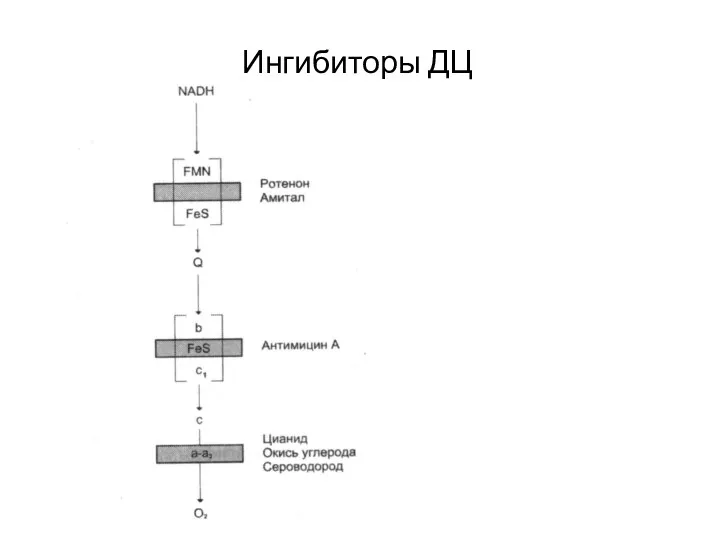
Ингибиторы ДЦ
Ингибиторы ДЦ
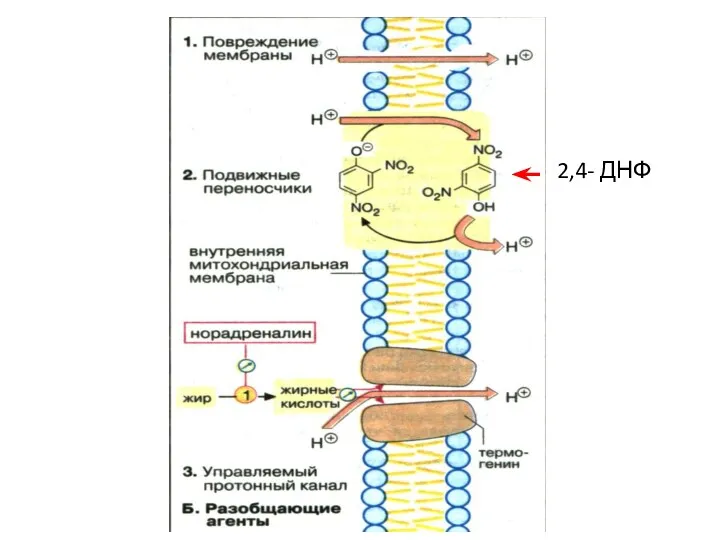
2,4- ДНФ
2,4- ДНФ
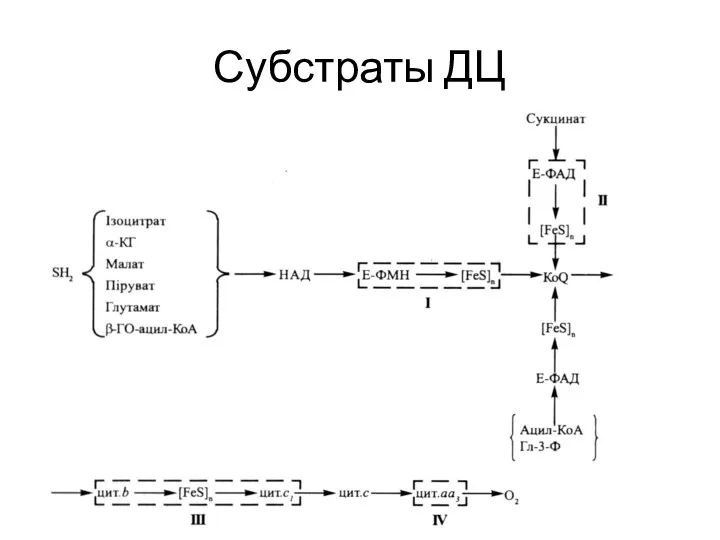
Субстраты ДЦ
Субстраты ДЦ
 Исследовательская работа. Какие удивительные эти пауки
Исследовательская работа. Какие удивительные эти пауки Сообщество, экосистема, биогеоценоз
Сообщество, экосистема, биогеоценоз Плоские черви. (Лекция 4)
Плоские черви. (Лекция 4) Цитрусовые. Виды. Строение
Цитрусовые. Виды. Строение Царство растения. Водоросли
Царство растения. Водоросли Анатомическое строение стебля
Анатомическое строение стебля Synecology or - Community Ecology лекция
Synecology or - Community Ecology лекция Класс земноводные, или амфибии
Класс земноводные, или амфибии Презентация по биологии Борьба за существование
Презентация по биологии Борьба за существование Находки останков мамонтов на территории России
Находки останков мамонтов на территории России Отдел Голосеменные. Гинкго. Гнетум
Отдел Голосеменные. Гинкго. Гнетум Генетика микроорганизмов
Генетика микроорганизмов Продукты, полезные для глаз
Продукты, полезные для глаз Биология как наука, её история. Методы исследования в биологии
Биология как наука, её история. Методы исследования в биологии Влияние фитонцидов различных растений на жизнедеятельность колорадского жука
Влияние фитонцидов различных растений на жизнедеятельность колорадского жука Биогеоценозы. Экосистемы. Строение и свойства
Биогеоценозы. Экосистемы. Строение и свойства Презентация к уроку Моногибридное скрещивание
Презентация к уроку Моногибридное скрещивание Деревья
Деревья Собачий клещ (Ixodes ricinus)
Собачий клещ (Ixodes ricinus) Одноклеточные животные
Одноклеточные животные Органи чуття та їх значення
Органи чуття та їх значення Ткани животных и растений
Ткани животных и растений Индивидуальное развитие (онтогенез) у животных
Индивидуальное развитие (онтогенез) у животных Изучение внешнего строения, скелета и зубной системы млекопитающих. Лабораторная работа. 7 класс
Изучение внешнего строения, скелета и зубной системы млекопитающих. Лабораторная работа. 7 класс Биология. Автор термина биология
Биология. Автор термина биология Линии в кролиководстве
Линии в кролиководстве Генетика пола
Генетика пола Симбиоз и симбиогенез организмов
Симбиоз и симбиогенез организмов