- Биофизика гормональных рецепторов

Содержание
- 2. 4 ОСНОВНЫЕ СИСТЕМЫ РЕГУЛЯЦИИ МЕТАБОЛИЗМА: Центральная нервная система за счет передачи сигналов посредством нервных импульсов и
- 3. ЭНДОКРИННАЯ СИСТЕМА РЕГУЛИРУЕТ ОБМЕН ВЕЩЕСТВ ПОСРЕДСТВОМ ГОРМОНОВ ГОРМОНЫ - БИОЛОГИЧЕСКИ АКТИВНЫЕ ОРГАНИЧЕСКИЕ СОЕДИНЕНИЯ, КОТОРЫЕ ВЫРАБАТЫВАЮТСЯ В
- 4. КЛАССИЧЕСКИМ ГОРМОНАМ ПРИСУЩ РЯД ПРИЗНАКОВ: Дистантность действия – синтез в железах внутренней секреции, а регуляция отдаленных
- 5. ОРГАНИЗАЦИЯ НЕРВНО-ГОРМОНАЛЬНОЙ РЕГУЛЯЦИИ Существует строгая иерархия или соподчиненность гормонов. Поддержание уровня гормонов в организме в большинстве
- 7. РЕГУЛЯЦИЯ УРОВНЯ ГОРМОНОВ В ОРГАНИЗМЕ Изменение концентрации метаболитов в клетках-мишенях по механизму отрицательной обратной связи подавляет
- 8. КЛАССИФИКАЦИЯ ГОРМОНОВ по биологическим функциям; по механизму действия; по химическому строению; различают 4 группы: 1. Белково-пептидные
- 9. КЛАССИФИКАЦИЯ ГОРМОНОВ ПО БИОЛОГИЧЕСКИМ ФУНКЦИЯМ ЭТА КЛАССИФИКАЦИЯ УСЛОВНА, Т.К. МОГУ ВЫПОЛНЯТЬ РАЗНЫЕ ФУНКЦИИ
- 10. КЛАССИФИКАЦИЯ ГОРМОНОВ ПО ХИМИЧЕСКОМУ СТРОЕНИЮ
- 11. 1. БЕЛКОВО - ПЕПТИДНЫЕ ГОРМОНЫ Гормоны гипоталамуса; гормоны гипофиза; гормоны поджелудочной железы - инсулин, глюкагон; гормоны
- 12. НЕКОТОРЫЕ ПРЕДСТАВИТЕЛИ БЕЛКОВО - ПЕПТИДНЫХ ГОРМОНОВ: ТИРЕОЛИБЕРИНА, ИНСУЛИНА И СОМАТОСТАТИНА
- 13. 2. ГОРМОНЫ - ПРОИЗВОДНЫЕ АМИНОКИСЛОТ Являются производными аминокислоты - тирозина. К ним относятся гормоны щитовидной железы
- 14. ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
- 15. СХЕМА СИНТЕЗА ТРИЙОДТИРОНИНОВ
- 16. 3. ГОРМОНЫ СТЕРОИДНОЙ ПРИРОДЫ Синтезируются из холестерина (на рис.) Гормоны коркового вещества надпочечников – кортикостероиды (кортизол,
- 17. СИНТЕЗ ОСНОВНЫХ КОРТИКОСТЕРОИДОВ
- 18. 4. ЭЙКОЗАНОИДЫ Предшественником всех эйкозаноидов является арахидоновая кислота. Они делятся на 3 группы – простагландины, лейкотриены,
- 19. ХАРАКТЕРИСТИКА РАЗНЫХ ГРУПП ЭЙКАЗОНОИДОВ Простагландины (Pg) — синтезируются практически во всех клетках, кроме эритроцитов и лимфоцитов.
- 20. ВЗАИМОДЕЙСТВИЕ ГОРМОНОВ С РЕЦЕПТОРАМИ КЛЕТОК-МИШЕНЕЙ Для проявления биологической активности связывание гормонов с рецепторами должно приводить к
- 21. ОБЩАЯ ХАРАКТЕРИСТИКА РЕЦЕПТОРОВ Рецепторы могут находится на поверхности клеточной мембраны, внутри клетки – в цитозоле или
- 22. ГОРМОНАЛЬНЫЙ СИГНАЛ МЕНЯЕТ СКОРОСТЬ МЕТАБОЛИЧЕСКИХ ПРОЦЕССОВ ОТВЕТ ПУТЕМ: - ИЗМЕНЕНИЯ АКТИВНОСТИ ФЕРМЕНТОВ - ИЗМЕНЕНИЕМ КОЛИЧЕСТВА ФЕРМЕНТОВ.
- 23. ПО МЕХАНИЗМУ ДЕЙСТВИЯ РАЗЛИЧАЮТ ГОРМОНЫ: - ВЗАИМОДЕЙСТВУЮЩИЕ С МЕМБРАННЫМИ РЕЦЕПТОРАМИ (ПЕПТИДНЫЕ ГОРМОНЫ, АДРЕНАЛИН, ЭЙКАЗОНОИДЫ) И -
- 24. ОСНОВНЫЕ ЭТАПЫ ПЕРЕДАЧИ ГОРМОНАЛЬНОГО СИГНАЛА: ЧЕРЕЗ МЕМБРАННЫЕ И ВНУТРИКЛЕТОЧНЫЕ РЕЦЕПТОРЫ
- 25. МЕХАНИЗМЫ ПЕРЕДАЧИ ГОРМОНАЛЬНОГО СИГНАЛА: БЫСТРЫЙ (МЕМБРАННЫЙ РЕЦЕПТОР) И МЕДЛЕННЫЙ (ВНУТРИКЛЕТОЧНЫЙ РЕЦЕПТОР) ТИПЫ
- 26. ПЕРЕДАЧА ГОРМОНАЛЬНОГО СИГНАЛА ЧЕРЕЗ АДЕНИЛАТЦИКЛАЗУ Комплекс гормон-рецептор связан с G – белком, который имеет 3 субъединицы
- 27. БИОЛОГИЧЕСКОЕ ДЕЙСТВИЕ ГОРМОНА РОСТА (ИФР – ИНСУЛИНОПОДОБНЫЙ ФАКТОР РОСТА)
- 30. ПУТИ ИСПОЛЬЗОВАНИЯ ЭНЕРГОНОСИТЕЛЕЙ И ВЛИЯНИЕ ГОРМОНОВ НА МЕТАБОЛИЗМ
- 32. Скачать презентацию

4 ОСНОВНЫЕ СИСТЕМЫ РЕГУЛЯЦИИ МЕТАБОЛИЗМА:
Центральная нервная система за счет передачи сигналов
4 ОСНОВНЫЕ СИСТЕМЫ РЕГУЛЯЦИИ МЕТАБОЛИЗМА:
Центральная нервная система за счет передачи сигналов

ЭНДОКРИННАЯ СИСТЕМА РЕГУЛИРУЕТ ОБМЕН ВЕЩЕСТВ ПОСРЕДСТВОМ ГОРМОНОВ
ГОРМОНЫ - БИОЛОГИЧЕСКИ АКТИВНЫЕ ОРГАНИЧЕСКИЕ
ЭНДОКРИННАЯ СИСТЕМА РЕГУЛИРУЕТ ОБМЕН ВЕЩЕСТВ ПОСРЕДСТВОМ ГОРМОНОВ ГОРМОНЫ - БИОЛОГИЧЕСКИ АКТИВНЫЕ ОРГАНИЧЕСКИЕ

КЛАССИЧЕСКИМ ГОРМОНАМ ПРИСУЩ РЯД ПРИЗНАКОВ:
Дистантность действия – синтез в железах внутренней
КЛАССИЧЕСКИМ ГОРМОНАМ ПРИСУЩ РЯД ПРИЗНАКОВ:
Дистантность действия – синтез в железах внутренней

ОРГАНИЗАЦИЯ НЕРВНО-ГОРМОНАЛЬНОЙ РЕГУЛЯЦИИ
Существует строгая иерархия или соподчиненность гормонов.
Поддержание уровня гормонов в
ОРГАНИЗАЦИЯ НЕРВНО-ГОРМОНАЛЬНОЙ РЕГУЛЯЦИИ
Существует строгая иерархия или соподчиненность гормонов.
Поддержание уровня гормонов в
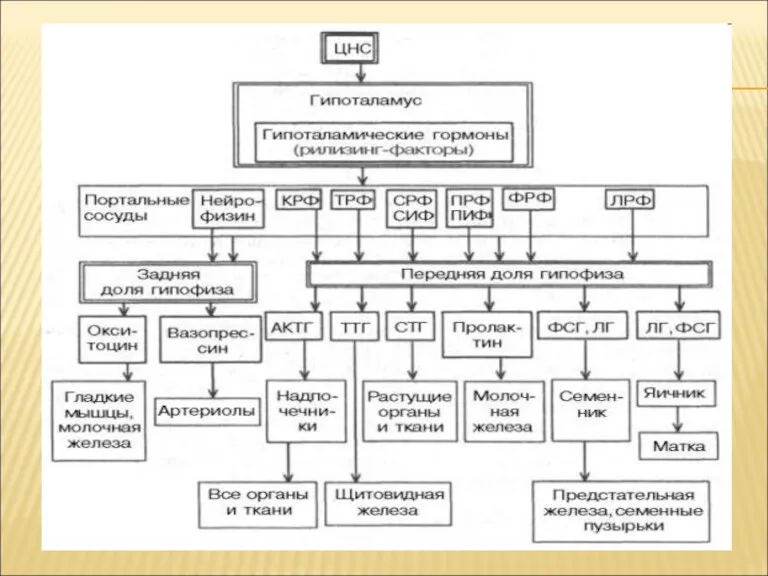

РЕГУЛЯЦИЯ УРОВНЯ ГОРМОНОВ В ОРГАНИЗМЕ
Изменение концентрации метаболитов в клетках-мишенях по механизму
РЕГУЛЯЦИЯ УРОВНЯ ГОРМОНОВ В ОРГАНИЗМЕ
Изменение концентрации метаболитов в клетках-мишенях по механизму

КЛАССИФИКАЦИЯ ГОРМОНОВ
по биологическим функциям;
по механизму действия;
по химическому строению;
различают 4 группы:
КЛАССИФИКАЦИЯ ГОРМОНОВ
по биологическим функциям;
по механизму действия;
по химическому строению;
различают 4 группы:

КЛАССИФИКАЦИЯ ГОРМОНОВ ПО БИОЛОГИЧЕСКИМ ФУНКЦИЯМ
ЭТА КЛАССИФИКАЦИЯ УСЛОВНА, Т.К. МОГУ ВЫПОЛНЯТЬ
КЛАССИФИКАЦИЯ ГОРМОНОВ ПО БИОЛОГИЧЕСКИМ ФУНКЦИЯМ ЭТА КЛАССИФИКАЦИЯ УСЛОВНА, Т.К. МОГУ ВЫПОЛНЯТЬ

КЛАССИФИКАЦИЯ ГОРМОНОВ ПО ХИМИЧЕСКОМУ СТРОЕНИЮ
КЛАССИФИКАЦИЯ ГОРМОНОВ ПО ХИМИЧЕСКОМУ СТРОЕНИЮ

1. БЕЛКОВО - ПЕПТИДНЫЕ
ГОРМОНЫ
Гормоны гипоталамуса; гормоны гипофиза; гормоны поджелудочной железы -
1. БЕЛКОВО - ПЕПТИДНЫЕ
ГОРМОНЫ
Гормоны гипоталамуса; гормоны гипофиза; гормоны поджелудочной железы -
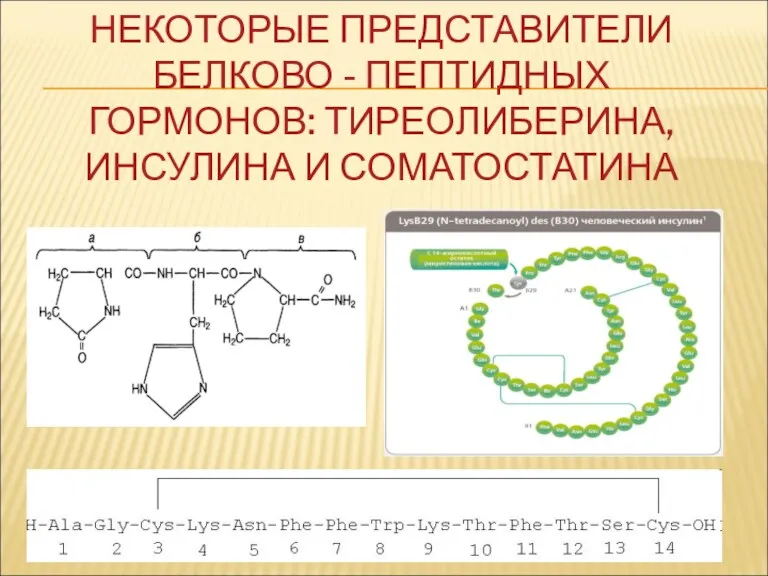
НЕКОТОРЫЕ ПРЕДСТАВИТЕЛИ БЕЛКОВО - ПЕПТИДНЫХ
ГОРМОНОВ: ТИРЕОЛИБЕРИНА, ИНСУЛИНА И СОМАТОСТАТИНА
НЕКОТОРЫЕ ПРЕДСТАВИТЕЛИ БЕЛКОВО - ПЕПТИДНЫХ
ГОРМОНОВ: ТИРЕОЛИБЕРИНА, ИНСУЛИНА И СОМАТОСТАТИНА
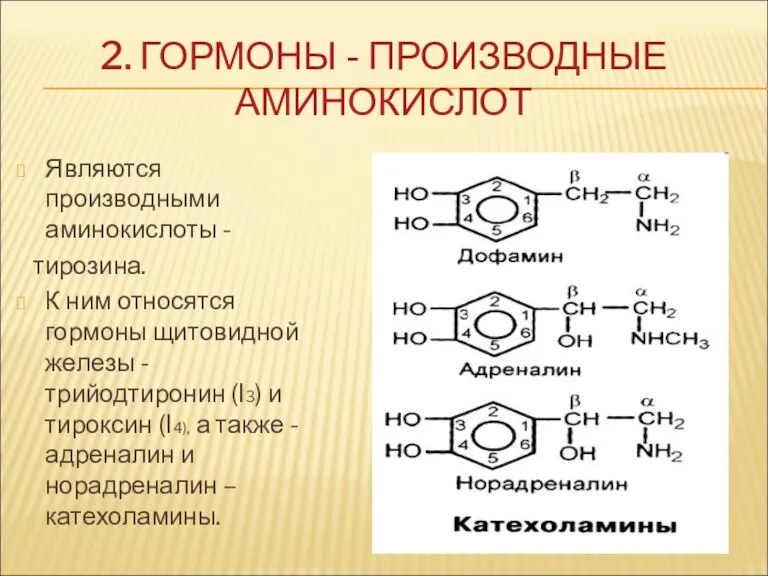
2. ГОРМОНЫ - ПРОИЗВОДНЫЕ АМИНОКИСЛОТ
Являются производными аминокислоты -
тирозина.
К ним
2. ГОРМОНЫ - ПРОИЗВОДНЫЕ АМИНОКИСЛОТ
Являются производными аминокислоты -
тирозина.
К ним
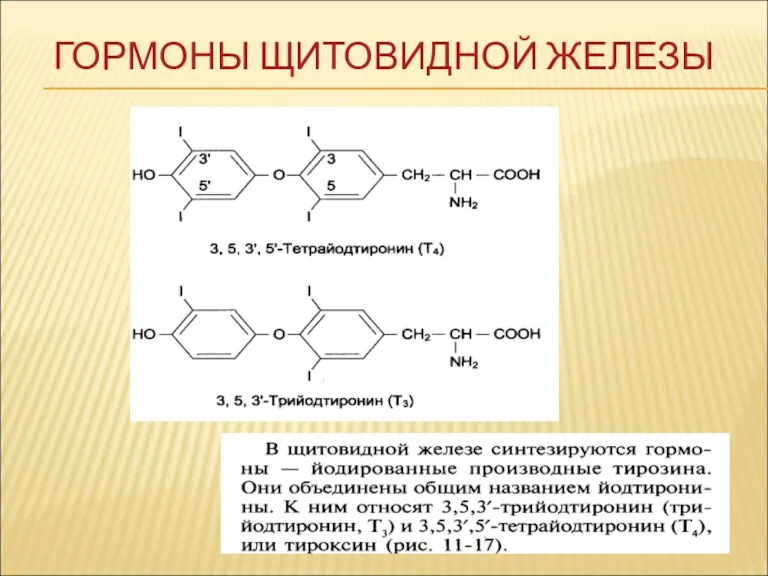
ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ

СХЕМА СИНТЕЗА ТРИЙОДТИРОНИНОВ
СХЕМА СИНТЕЗА ТРИЙОДТИРОНИНОВ
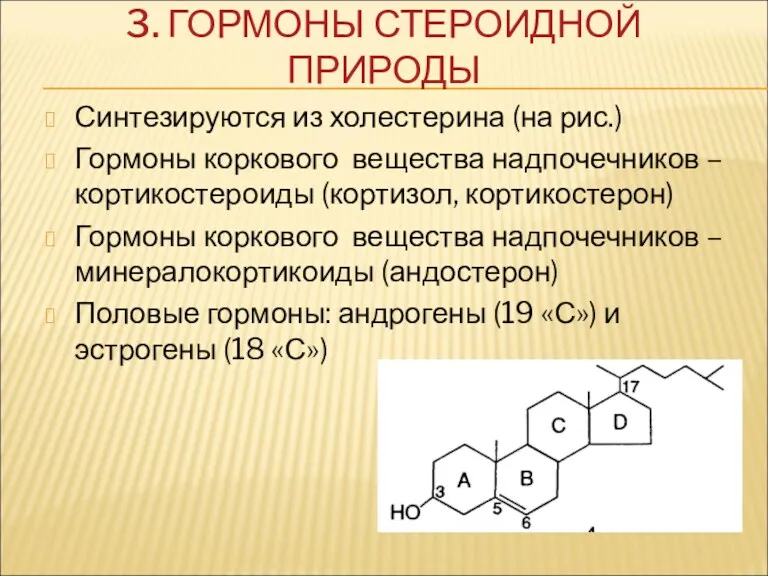
3. ГОРМОНЫ СТЕРОИДНОЙ ПРИРОДЫ
Синтезируются из холестерина (на рис.)
Гормоны коркового вещества надпочечников
3. ГОРМОНЫ СТЕРОИДНОЙ ПРИРОДЫ
Синтезируются из холестерина (на рис.)
Гормоны коркового вещества надпочечников
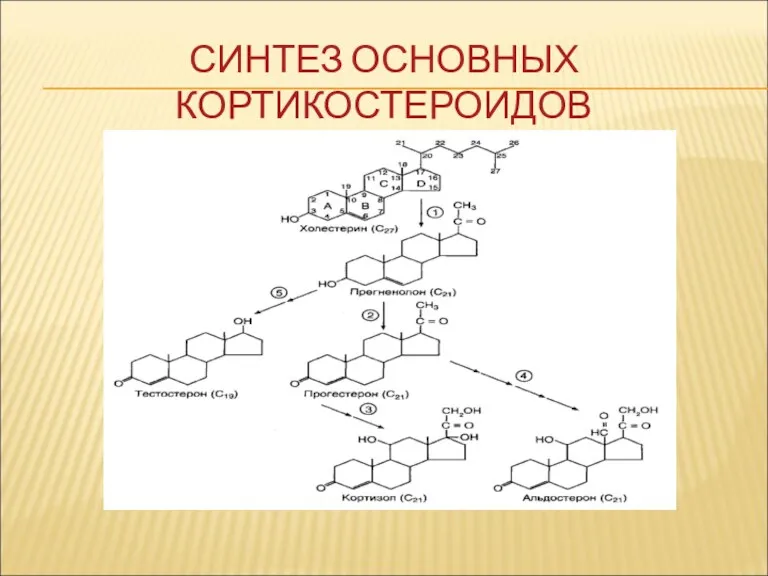
СИНТЕЗ ОСНОВНЫХ КОРТИКОСТЕРОИДОВ
СИНТЕЗ ОСНОВНЫХ КОРТИКОСТЕРОИДОВ
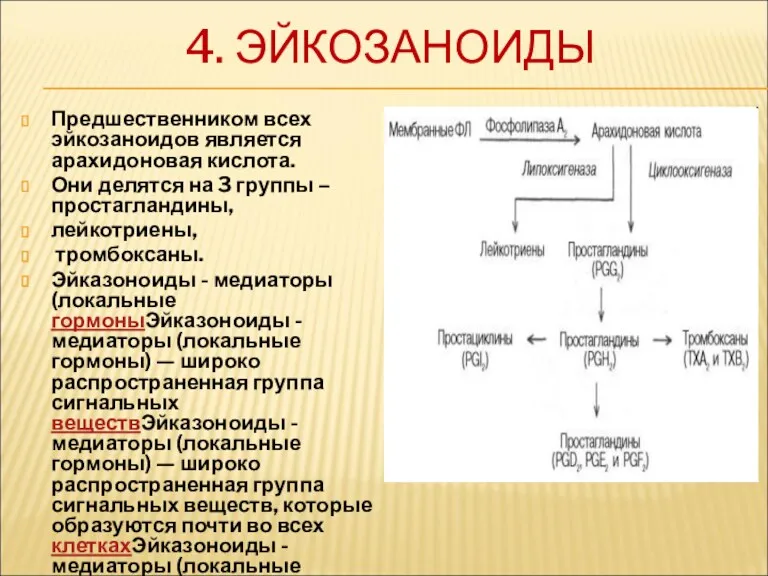
4. ЭЙКОЗАНОИДЫ
Предшественником всех эйкозаноидов является арахидоновая кислота.
Они делятся на 3
4. ЭЙКОЗАНОИДЫ
Предшественником всех эйкозаноидов является арахидоновая кислота.
Они делятся на 3

ХАРАКТЕРИСТИКА РАЗНЫХ ГРУПП ЭЙКАЗОНОИДОВ
Простагландины (Pg) — синтезируются практически во всех клетках, кроме
ХАРАКТЕРИСТИКА РАЗНЫХ ГРУПП ЭЙКАЗОНОИДОВ
Простагландины (Pg) — синтезируются практически во всех клетках, кроме

ВЗАИМОДЕЙСТВИЕ ГОРМОНОВ С РЕЦЕПТОРАМИ КЛЕТОК-МИШЕНЕЙ
Для проявления биологической активности связывание гормонов с
ВЗАИМОДЕЙСТВИЕ ГОРМОНОВ С РЕЦЕПТОРАМИ КЛЕТОК-МИШЕНЕЙ
Для проявления биологической активности связывание гормонов с

ОБЩАЯ ХАРАКТЕРИСТИКА РЕЦЕПТОРОВ
Рецепторы могут находится на поверхности клеточной мембраны, внутри клетки
ОБЩАЯ ХАРАКТЕРИСТИКА РЕЦЕПТОРОВ
Рецепторы могут находится на поверхности клеточной мембраны, внутри клетки

ГОРМОНАЛЬНЫЙ СИГНАЛ МЕНЯЕТ СКОРОСТЬ МЕТАБОЛИЧЕСКИХ ПРОЦЕССОВ ОТВЕТ ПУТЕМ:
- ИЗМЕНЕНИЯ АКТИВНОСТИ
ГОРМОНАЛЬНЫЙ СИГНАЛ МЕНЯЕТ СКОРОСТЬ МЕТАБОЛИЧЕСКИХ ПРОЦЕССОВ ОТВЕТ ПУТЕМ: - ИЗМЕНЕНИЯ АКТИВНОСТИ

ПО МЕХАНИЗМУ ДЕЙСТВИЯ РАЗЛИЧАЮТ ГОРМОНЫ:
- ВЗАИМОДЕЙСТВУЮЩИЕ С МЕМБРАННЫМИ РЕЦЕПТОРАМИ (ПЕПТИДНЫЕ
ПО МЕХАНИЗМУ ДЕЙСТВИЯ РАЗЛИЧАЮТ ГОРМОНЫ: - ВЗАИМОДЕЙСТВУЮЩИЕ С МЕМБРАННЫМИ РЕЦЕПТОРАМИ (ПЕПТИДНЫЕ

ОСНОВНЫЕ ЭТАПЫ ПЕРЕДАЧИ ГОРМОНАЛЬНОГО СИГНАЛА:
ЧЕРЕЗ МЕМБРАННЫЕ И ВНУТРИКЛЕТОЧНЫЕ РЕЦЕПТОРЫ
ОСНОВНЫЕ ЭТАПЫ ПЕРЕДАЧИ ГОРМОНАЛЬНОГО СИГНАЛА:
ЧЕРЕЗ МЕМБРАННЫЕ И ВНУТРИКЛЕТОЧНЫЕ РЕЦЕПТОРЫ
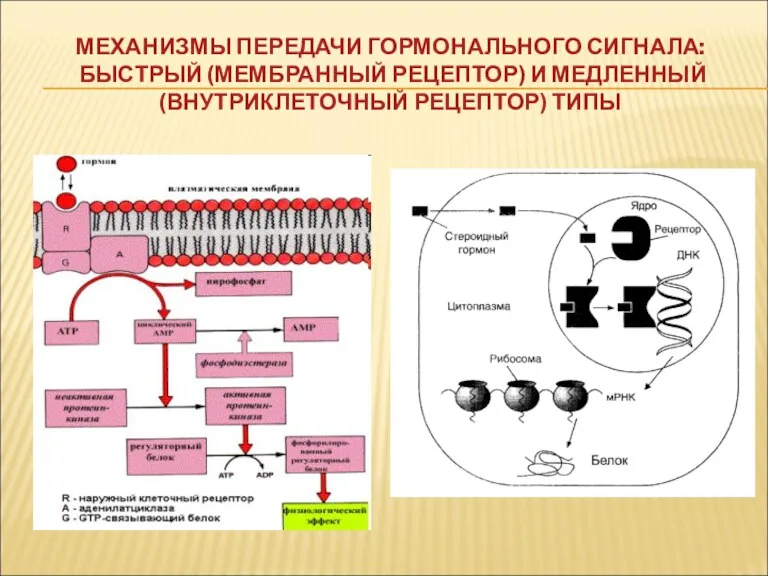
МЕХАНИЗМЫ ПЕРЕДАЧИ ГОРМОНАЛЬНОГО СИГНАЛА:
БЫСТРЫЙ (МЕМБРАННЫЙ РЕЦЕПТОР) И МЕДЛЕННЫЙ (ВНУТРИКЛЕТОЧНЫЙ РЕЦЕПТОР)
МЕХАНИЗМЫ ПЕРЕДАЧИ ГОРМОНАЛЬНОГО СИГНАЛА: БЫСТРЫЙ (МЕМБРАННЫЙ РЕЦЕПТОР) И МЕДЛЕННЫЙ (ВНУТРИКЛЕТОЧНЫЙ РЕЦЕПТОР)
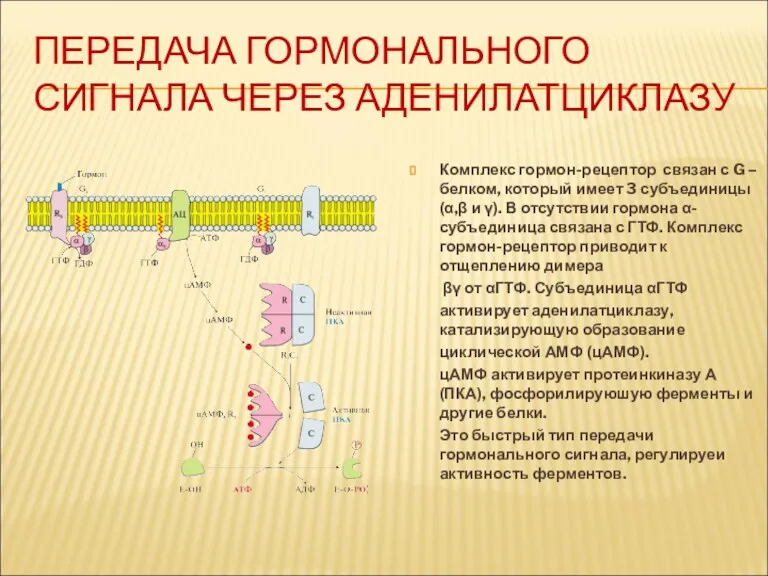
ПЕРЕДАЧА ГОРМОНАЛЬНОГО СИГНАЛА ЧЕРЕЗ АДЕНИЛАТЦИКЛАЗУ
Комплекс гормон-рецептор связан с G – белком,
ПЕРЕДАЧА ГОРМОНАЛЬНОГО СИГНАЛА ЧЕРЕЗ АДЕНИЛАТЦИКЛАЗУ
Комплекс гормон-рецептор связан с G – белком,
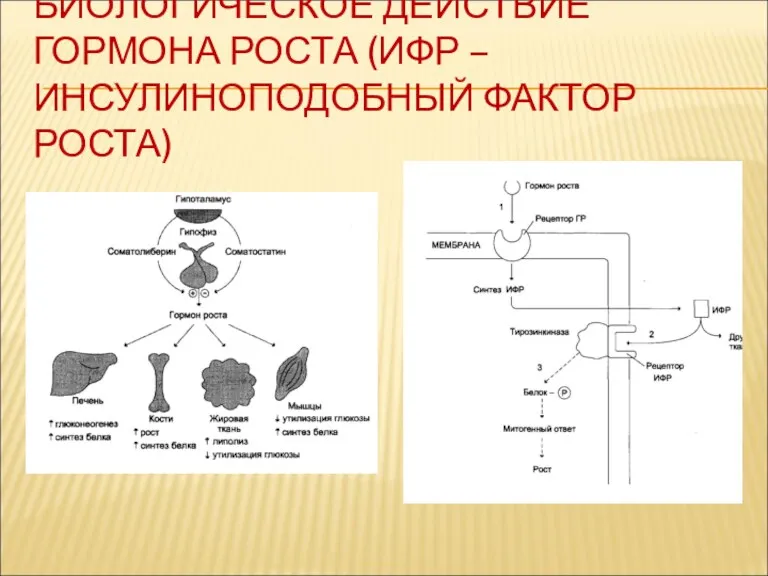
БИОЛОГИЧЕСКОЕ ДЕЙСТВИЕ ГОРМОНА РОСТА (ИФР – ИНСУЛИНОПОДОБНЫЙ ФАКТОР РОСТА)
БИОЛОГИЧЕСКОЕ ДЕЙСТВИЕ ГОРМОНА РОСТА (ИФР – ИНСУЛИНОПОДОБНЫЙ ФАКТОР РОСТА)

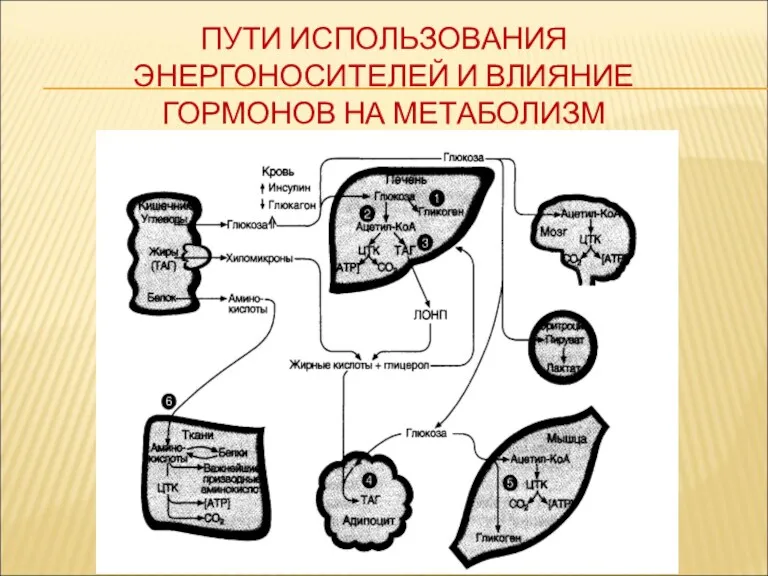
ПУТИ ИСПОЛЬЗОВАНИЯ ЭНЕРГОНОСИТЕЛЕЙ И ВЛИЯНИЕ ГОРМОНОВ НА МЕТАБОЛИЗМ
ПУТИ ИСПОЛЬЗОВАНИЯ ЭНЕРГОНОСИТЕЛЕЙ И ВЛИЯНИЕ ГОРМОНОВ НА МЕТАБОЛИЗМ
 Поле и его обитатели
Поле и его обитатели Биотические факторы среды
Биотические факторы среды Лесные дары. Ягоды
Лесные дары. Ягоды Опорно-двигательная система человека
Опорно-двигательная система человека Костная и мышечная системы. Скелет головы
Костная и мышечная системы. Скелет головы Антропогенез
Антропогенез Ботаника
Ботаника Кальцій у організмі людини
Кальцій у організмі людини Энергетический обмен. Метаболизм (обмен веществ)
Энергетический обмен. Метаболизм (обмен веществ) Фиксация молекулярного азота (азотфиксация, диазотрофия) микроорганизмами
Фиксация молекулярного азота (азотфиксация, диазотрофия) микроорганизмами Семенные растения
Семенные растения Структурные компоненты клетки и их физиологические функции
Структурные компоненты клетки и их физиологические функции Технология выращивания комнатных растений
Технология выращивания комнатных растений Орган зрения
Орган зрения Размножение и развитие животных. 3 класс
Размножение и развитие животных. 3 класс Биологические полимеры - нуклеиновые кислоты
Биологические полимеры - нуклеиновые кислоты Экскурсия по Лапландскому заповеднику
Экскурсия по Лапландскому заповеднику Презентация к внеклассному мероприятию Муравьи- общественные насекомые
Презентация к внеклассному мероприятию Муравьи- общественные насекомые презентация к уроку по теме: Цветок. Строение и значение цветка
презентация к уроку по теме: Цветок. Строение и значение цветка Задачи по генетике
Задачи по генетике Биотические факторы
Биотические факторы Слуховая сенсорная система
Слуховая сенсорная система Приспособленность организмов к определенной среде обитания
Приспособленность организмов к определенной среде обитания Генетична рівновага в популяціях і її порушення
Генетична рівновага в популяціях і її порушення Государственная итоговая аттестация обучающихся за основное общее образование по биологии в 2023 году
Государственная итоговая аттестация обучающихся за основное общее образование по биологии в 2023 году Лекарственные и съедобные растения, произрастающие в Якутии
Лекарственные и съедобные растения, произрастающие в Якутии Питание и пищеварение
Питание и пищеварение Красная книга растений
Красная книга растений