- Биофизика рецепции. Проблема клеточного узнавания

Содержание
- 2. Проблема клеточного узнавания. I. При формировании ткани и в ходе её функционирования важную роль играют процессы
- 3. Рецептор(-ы) – высокоспециализированное образование, способное воспринять, трансформировать и передать энергию внешнего стимула в нервную систему. Рецептором
- 4. Механорецепторы приспособлены к восприятию механической энергии раздражающего стимула. Восприятие механического раздражителя необходимо как самым низшим организмам,
- 5. 3. Хеморецепторы чувствительны к действию химических агентов. У наземных животных они образуют периферические отделы обонятельной и
- 6. Светочувствительные клетки – палочки и колбочки – снабжены пигментом родопсином, поглощающим свет. Палочки ответственны за сумеречное,
- 7. В основе процесса фоторецепции лежит взаимодействие квантов электромагнитного излучения с молекулами хромолипопротеидов, служащих зрительными пигментами. Следующая
- 8. Механорецепторы – многочисленная группа рецепторов, в которых процессы восприятия и трансформации внешнего воздействия происходят при условии
- 9. Органом равновесия у млекопитающих является вестибулярный аппарат, расположенный во внутреннем ухе. Его рецепторные клетки снабжены волосками.
- 10. Тельце Мейснера (осязательное тельце) –инкапсулированное нервное окончание. Присутствует в дерме кожи, особенно часто в кончиках пальцев,
- 11. Пахучие вещества, находящиеся в воздухе, проникают через слизь и стимулируют обонятельные клетки. Возможно, существует несколько основных
- 12. Терморецептор – группа чувствительных элементов, реагирующих на изменение температуры окружающей среды. Различают тканевые и центральные терморецепторы
- 13. Рецепторы преобразуют энергию раздражителя в электрический сигнал, который возбуждает нейроны. Механизм возбуждения рецепторов связан с изменением
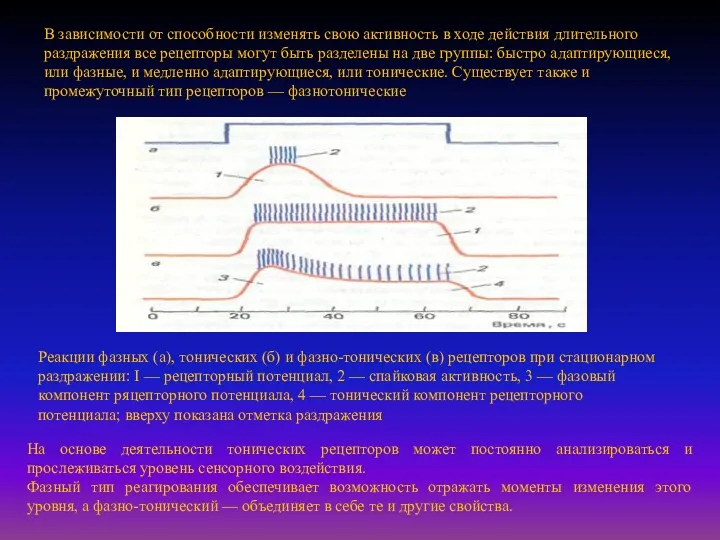
- 14. В зависимости от способности изменять свою активность в ходе действия длительного раздражения все рецепторы могут быть
- 15. Фазные рецепторы возбуждаются в начальный и конечный периоды деформации их мембран при низких (тактильные рецепторы) и
- 16. Адаптационные процессы в рецепторах могут определяться внешними и внутренними факторами. В качестве внешнего фактора в механизме
- 17. Проблема сопряжения между первичным взаимодействием внешнего стимула с рецепторным субстратом и генерацией рецепторного (генераторного) потенциала. Рецепторный
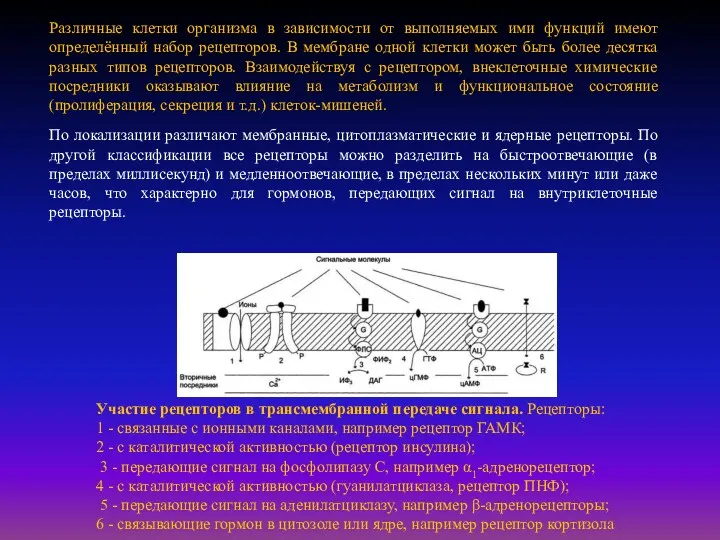
- 18. Роль структуры плазматической мембраны в процессе передачи сигнала. "Узнавание" сигнальных молекул осуществляется с помощью белков-рецепторов, встроенных
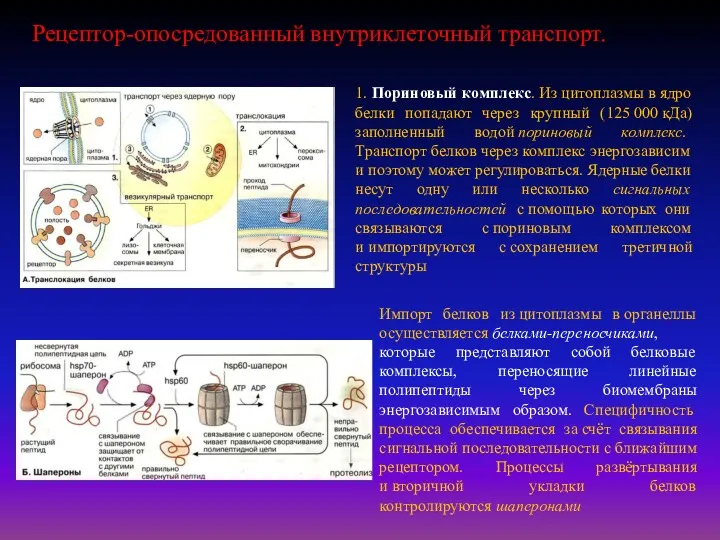
- 19. 1. Пориновый комплекс. Из цитоплазмы в ядро белки попадают через крупный (125 000 кДа) заполненный водой
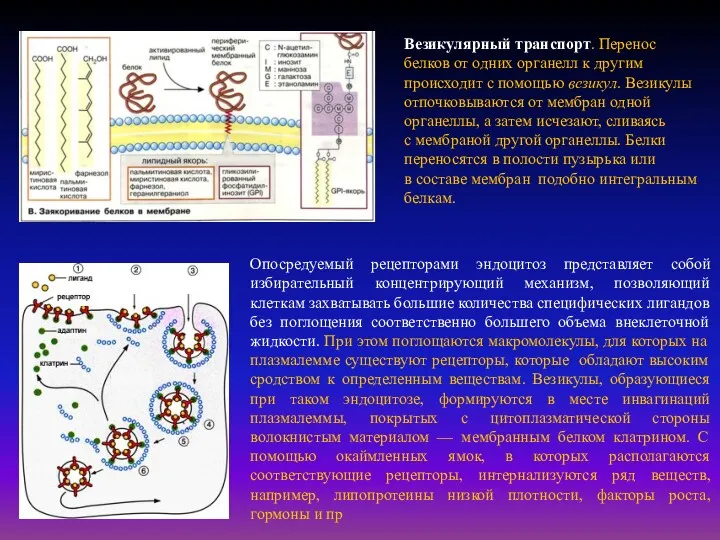
- 20. Опосредуемый рецепторами эндоцитоз представляет собой избирательный концентрирующий механизм, позволяющий клеткам захватывать большие количества специфических лигандов без
- 21. Участие рецепторов в трансмембранной передаче сигнала. Рецепторы: 1 - связанные с ионными каналами, например рецептор ГАМК;
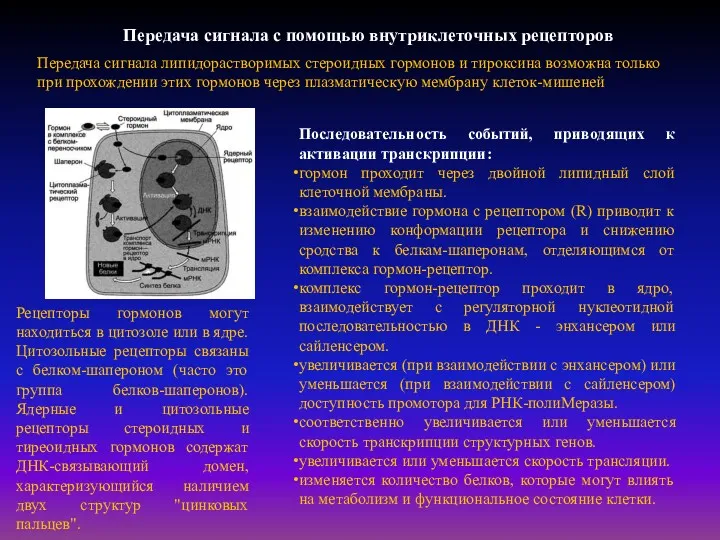
- 22. Передача сигнала с помощью внутриклеточных рецепторов Передача сигнала липидорастворимых стероидных гормонов и тироксина возможна только при
- 23. Рецепторы первого типа - интегральные олигомерные белки, содержащие субъединицу, имеющую центр для связывания сигнальной молекулы и
- 24. Индуктор, соединяясь с рецептором, вызывает конформационный переход, приводящий к открытию канала. После открытия канала сродство рецептора
- 25. Рецепторы второго типа, локализованные в мембранах и не связанные с каналами, подразделяют на 2 большие группы:
- 26. Рецептор инсулина - тирозиновая протеинкиназа, фосфорилирующая белки по ОН-группам тирозина. Рецептор состоит из двух α- и
- 27. Гуанилатциклаза катализирует образование цГМФ из ГТФ, одного из важных посредников внутриклеточной передачи сигнала Регуляция активности мембранной
- 28. G-белки (ГТФ-связывающие белки) - универсальные посредники при передаче сигналов от рецепторов к ферментам клеточной мембраны, катализирующим
- 29. Фермент аденилатциклаза, катализирующий превращение АТФ в цАМФ, - ключевой фермент аденилатциклазной системы передачи сигнала. При участии
- 30. Последовательность событий, приводящих к активации аденилатциклазы: связывание активатора аденилатциклазной системы, например гормона (Г) с рецептором (Rs),
- 31. Каскадный механизм усиления и подавления сигнала. Передача сигнала от мембранного рецептора через G-белок на фермент аденилатциклазу
- 32. Функционирование инозитолфосфатной системы трансмембранной передачи сигнала обеспечивают: R (рецептор), фосфолипаза С, Gplc - белок, активирующий фосфолипазу
- 34. Скачать презентацию

Проблема клеточного узнавания.
I. При формировании ткани и в ходе её
Проблема клеточного узнавания.
I. При формировании ткани и в ходе её

Рецептор(-ы) – высокоспециализированное образование, способное воспринять, трансформировать и передать энергию внешнего стимула
Рецептор(-ы) – высокоспециализированное образование, способное воспринять, трансформировать и передать энергию внешнего стимула

Механорецепторы приспособлены к восприятию механической энергии раздражающего стимула. Восприятие механического раздражителя
Механорецепторы приспособлены к восприятию механической энергии раздражающего стимула. Восприятие механического раздражителя

3. Хеморецепторы чувствительны к действию химических агентов. У наземных животных они
3. Хеморецепторы чувствительны к действию химических агентов. У наземных животных они

Светочувствительные клетки – палочки и колбочки – снабжены пигментом родопсином, поглощающим
Светочувствительные клетки – палочки и колбочки – снабжены пигментом родопсином, поглощающим
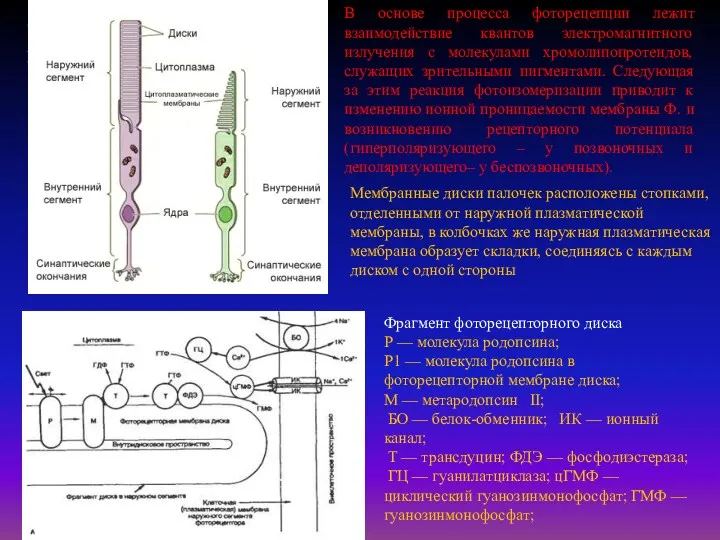
В основе процесса фоторецепции лежит взаимодействие квантов электромагнитного излучения с молекулами
В основе процесса фоторецепции лежит взаимодействие квантов электромагнитного излучения с молекулами
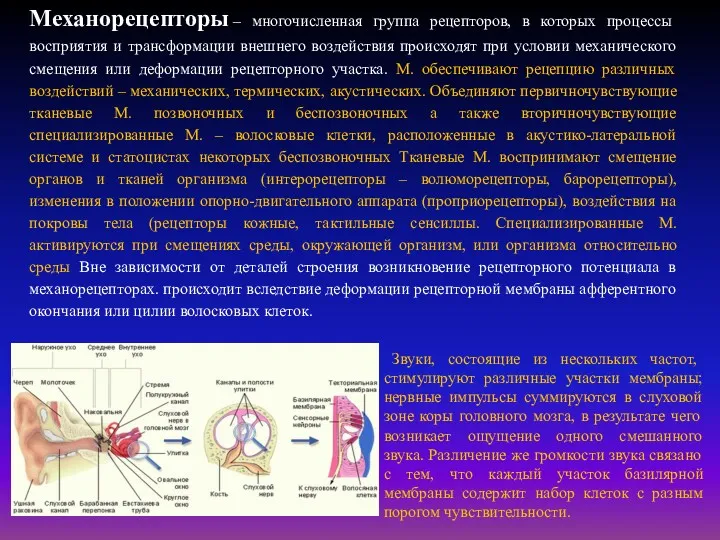
Механорецепторы – многочисленная группа рецепторов, в которых процессы восприятия и трансформации внешнего
Механорецепторы – многочисленная группа рецепторов, в которых процессы восприятия и трансформации внешнего
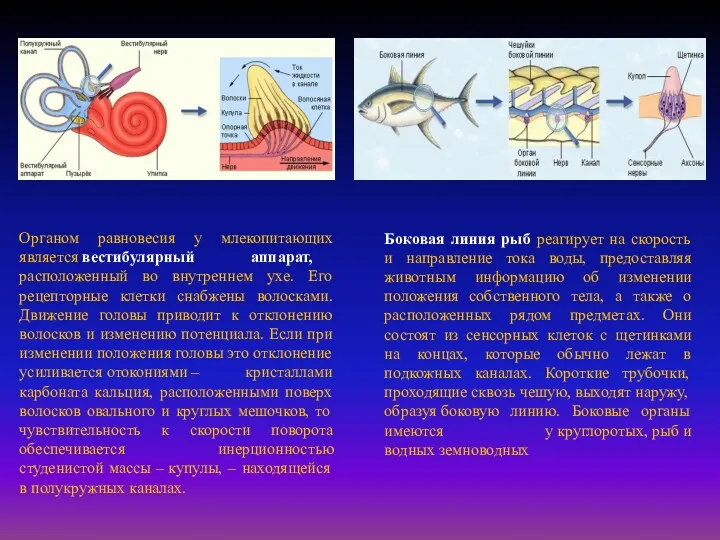
Органом равновесия у млекопитающих является вестибулярный аппарат, расположенный во внутреннем ухе. Его
Органом равновесия у млекопитающих является вестибулярный аппарат, расположенный во внутреннем ухе. Его

Тельце Мейснера (осязательное тельце) –инкапсулированное нервное окончание. Присутствует в дерме кожи, особенно часто в
Тельце Мейснера (осязательное тельце) –инкапсулированное нервное окончание. Присутствует в дерме кожи, особенно часто в
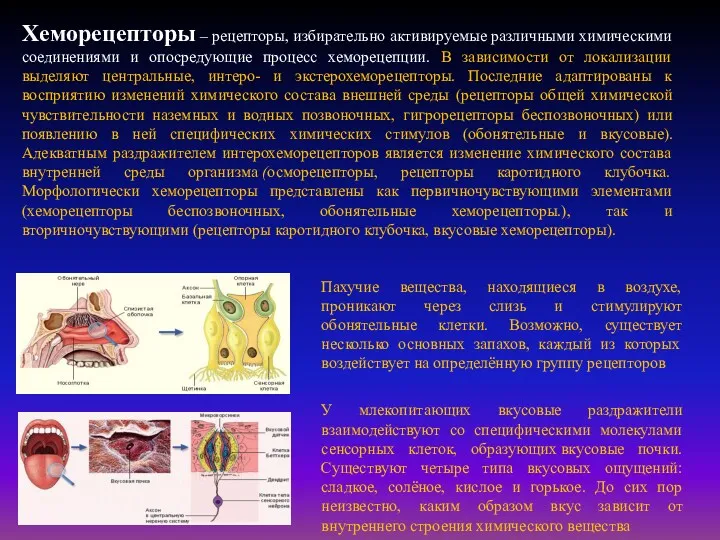
Пахучие вещества, находящиеся в воздухе, проникают через слизь и стимулируют обонятельные
Пахучие вещества, находящиеся в воздухе, проникают через слизь и стимулируют обонятельные

Терморецептор – группа чувствительных элементов, реагирующих на изменение температуры окружающей среды. Различают
Терморецептор – группа чувствительных элементов, реагирующих на изменение температуры окружающей среды. Различают

Рецепторы преобразуют энергию раздражителя в электрический сигнал, который возбуждает нейроны. Механизм
Рецепторы преобразуют энергию раздражителя в электрический сигнал, который возбуждает нейроны. Механизм
В зависимости от способности изменять свою активность в ходе действия длительного
В зависимости от способности изменять свою активность в ходе действия длительного

Фазные рецепторы возбуждаются в начальный и конечный периоды деформации их мембран
Фазные рецепторы возбуждаются в начальный и конечный периоды деформации их мембран

Адаптационные процессы в рецепторах могут определяться внешними и внутренними факторами. В
Адаптационные процессы в рецепторах могут определяться внешними и внутренними факторами. В

Проблема сопряжения между первичным взаимодействием внешнего стимула с рецепторным субстратом и
Проблема сопряжения между первичным взаимодействием внешнего стимула с рецепторным субстратом и

Роль структуры плазматической мембраны
в процессе передачи сигнала.
"Узнавание" сигнальных молекул осуществляется
Роль структуры плазматической мембраны
в процессе передачи сигнала.
"Узнавание" сигнальных молекул осуществляется
1. Пориновый комплекс. Из цитоплазмы в ядро белки попадают через крупный (125 000 кДа) заполненный
1. Пориновый комплекс. Из цитоплазмы в ядро белки попадают через крупный (125 000 кДа) заполненный
Опосредуемый рецепторами эндоцитоз представляет собой избирательный концентрирующий механизм, позволяющий клеткам захватывать
Опосредуемый рецепторами эндоцитоз представляет собой избирательный концентрирующий механизм, позволяющий клеткам захватывать
Участие рецепторов в трансмембранной передаче сигнала. Рецепторы:
1 - связанные с ионными
Участие рецепторов в трансмембранной передаче сигнала. Рецепторы:
1 - связанные с ионными
Передача сигнала с помощью внутриклеточных рецепторов
Передача сигнала липидорастворимых стероидных гормонов и
Передача сигнала с помощью внутриклеточных рецепторов
Передача сигнала липидорастворимых стероидных гормонов и

Рецепторы первого типа - интегральные олигомерные белки, содержащие субъединицу, имеющую центр
Рецепторы первого типа - интегральные олигомерные белки, содержащие субъединицу, имеющую центр
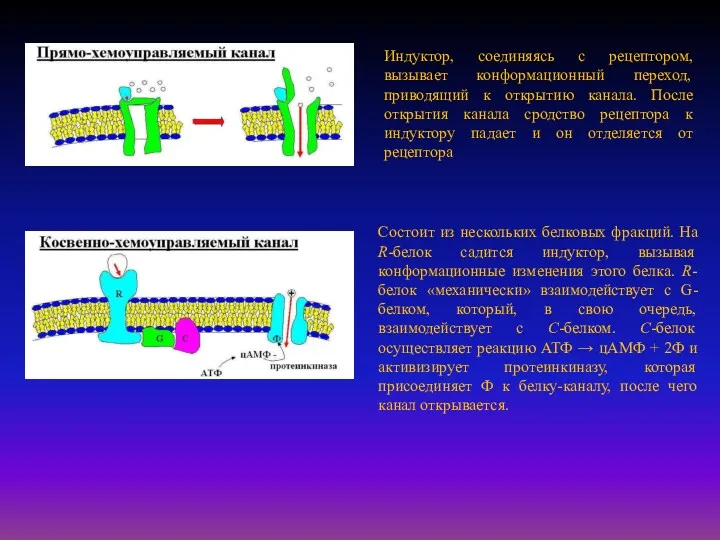
Индуктор, соединяясь с рецептором, вызывает конформационный переход, приводящий к открытию канала.
Индуктор, соединяясь с рецептором, вызывает конформационный переход, приводящий к открытию канала.

Рецепторы второго типа, локализованные в мембранах и не связанные с каналами,
Рецепторы второго типа, локализованные в мембранах и не связанные с каналами,
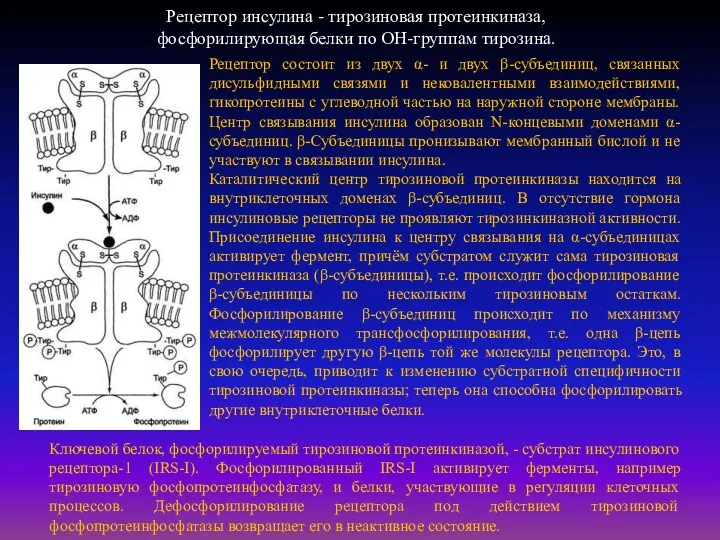
Рецептор инсулина - тирозиновая протеинкиназа,
фосфорилирующая белки по ОН-группам тирозина.
Рецептор состоит
Рецептор инсулина - тирозиновая протеинкиназа,
фосфорилирующая белки по ОН-группам тирозина.
Рецептор состоит
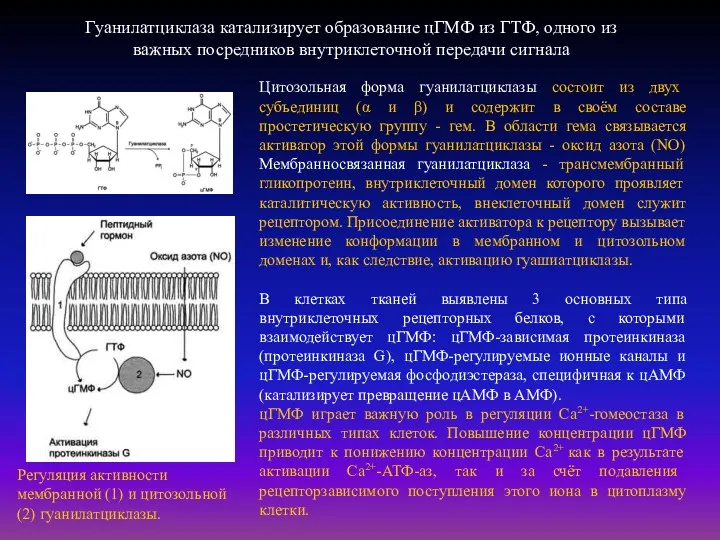
Гуанилатциклаза катализирует образование цГМФ из ГТФ, одного из важных посредников внутриклеточной
Гуанилатциклаза катализирует образование цГМФ из ГТФ, одного из важных посредников внутриклеточной

G-белки (ГТФ-связывающие белки) - универсальные посредники при передаче сигналов от рецепторов
G-белки (ГТФ-связывающие белки) - универсальные посредники при передаче сигналов от рецепторов
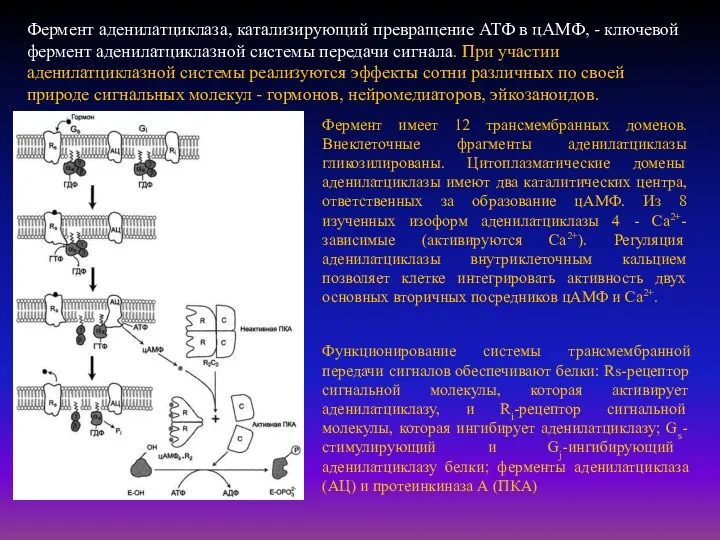
Фермент аденилатциклаза, катализирующий превращение АТФ в цАМФ, - ключевой фермент аденилатциклазной
Фермент аденилатциклаза, катализирующий превращение АТФ в цАМФ, - ключевой фермент аденилатциклазной

Последовательность событий, приводящих к активации аденилатциклазы:
связывание активатора аденилатциклазной системы, например гормона
Последовательность событий, приводящих к активации аденилатциклазы:
связывание активатора аденилатциклазной системы, например гормона

Каскадный механизм усиления и подавления сигнала.
Передача сигнала от мембранного рецептора через
Каскадный механизм усиления и подавления сигнала.
Передача сигнала от мембранного рецептора через
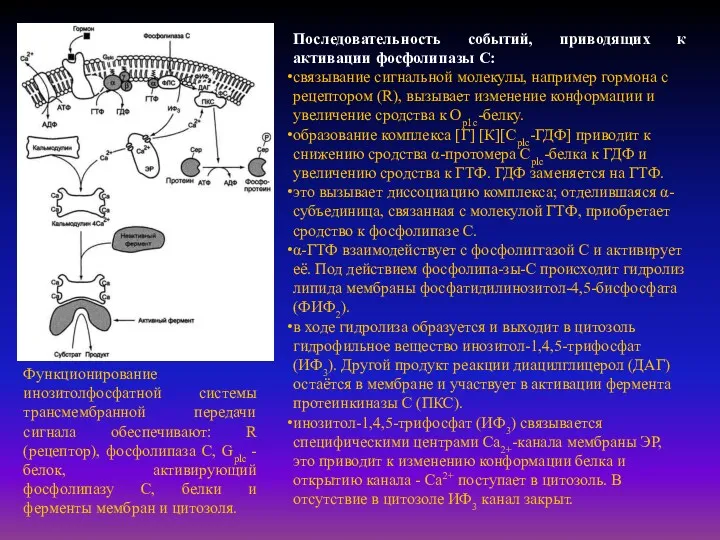
Функционирование инозитолфосфатной системы трансмембранной передачи сигнала обеспечивают: R (рецептор), фосфолипаза С,
Функционирование инозитолфосфатной системы трансмембранной передачи сигнала обеспечивают: R (рецептор), фосфолипаза С,
 презентация по биологии Гидра пресноводнаядля 7 класса
презентация по биологии Гидра пресноводнаядля 7 класса Голосеменные растения
Голосеменные растения 20231130_9_klass._stroenie_lyogkih._gazoobmen
20231130_9_klass._stroenie_lyogkih._gazoobmen Жизнь и творчество И.И.Левитана
Жизнь и творчество И.И.Левитана Генетика 2
Генетика 2 Моногибридное скрещивание
Моногибридное скрещивание Растения Донского края
Растения Донского края Удивительные факты из жизни рыб
Удивительные факты из жизни рыб Тема Тип Членистоногие
Тема Тип Членистоногие Влияние социальных факторов на развитие и становление вида Homo Sapiens
Влияние социальных факторов на развитие и становление вида Homo Sapiens Сравнительная характеристика классов типа Моллюски
Сравнительная характеристика классов типа Моллюски Рукотворная жизнь. Окружающий мир. 4 класс
Рукотворная жизнь. Окружающий мир. 4 класс Animal’s world
Animal’s world Спинной мозг (medulla spinalis). Лекция №3
Спинной мозг (medulla spinalis). Лекция №3 Красота мира растений
Красота мира растений Пищеварение в ротовой полости
Пищеварение в ротовой полости Ядовитые растения, произрастающие в Ленинградской области
Ядовитые растения, произрастающие в Ленинградской области Fruits and their classification. Spreading of fruits and seeds
Fruits and their classification. Spreading of fruits and seeds Пищеварение в желудке и 12-п. кишке. Лекция 26
Пищеварение в желудке и 12-п. кишке. Лекция 26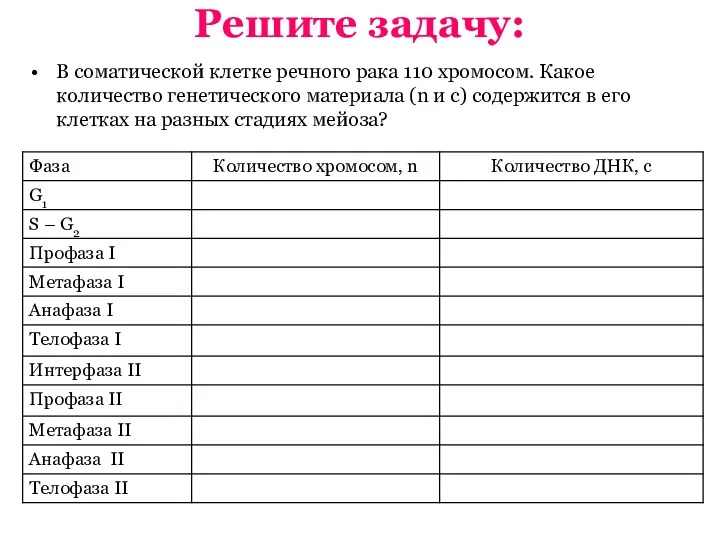 Формы размножения живых организмов
Формы размножения живых организмов Грибы. Окружающий мир 3 класс (по учебнику Н.Ф.Виноградовой, Г.С. Калиновой)
Грибы. Окружающий мир 3 класс (по учебнику Н.Ф.Виноградовой, Г.С. Калиновой) Царства живой природы
Царства живой природы Паразитизм в типе. Плоские черви
Паразитизм в типе. Плоские черви Мейоз. Первое и второе мейотическое деление. Биологическое значение кроссинговера
Мейоз. Первое и второе мейотическое деление. Биологическое значение кроссинговера Человек родился. Биологическая наследственность
Человек родился. Биологическая наследственность Электрофорез белков в геле
Электрофорез белков в геле Тип Черви
Тип Черви Вступ до курсу медичної біології. Структурно-функціональна організація клітини
Вступ до курсу медичної біології. Структурно-функціональна організація клітини